Spread of Oropouche Virus into the Central Nervous System in Mouse


Abstract
:1. Introduction
2. Materials and Methods

{kind=link}
| ANIMAL | 20D/2011 | 20C/2011 | 13E/2011 | 13*/2011 | 007/2011 | 006/2011 | 12A/2011 | 12B/2011 | 12C/2011 | 008/2011 | 21B/2011 | 21A/2011 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3dpi | 3dpi | 3dpi | 3dpi | 3dpi | 3dpi | 4dpi | 4dpi | 4dpi | 4dpi | 6dpi | 6dpi | |||
| Telencephalon: | Neocortex | Auditory | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | na | na | na | na | 0 (0/0) | 0 (0/0) |
| Motor | ++ (0/++) | + (0/++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (+/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | + (0/++) | 0 (0/0) | 0 (0/0) | ||
| Prefrontal | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (+++/++++) | 0 (0/0) | 0 (0/0) | ||
| Retrosplenial | + (0/++) | ++ (+/++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (+/+++++) | 0 (0/0) | 0 (0/0) | ||
| Somatosensory | + (0/+) | + (0/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++ (+/++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (+/++++) | 0 (0/0) | 0 (0/0) | ||
| Visual | na | + (+/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | + (+/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | * (0/*) | 0 (0/0) | 0 (0/0) | ||
| Piriform | * (0/*) | + (0/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (+/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | ||
| Hippoc | CA1 | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | + (+/+) | 0 (0/0) | 0 (0/0) | |
| CA3 | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | ||
| Dentate gyrus | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (+++/+++) | 0 (0/0) | 0 (0/0) | ||
| Subcortical nuclei | Nucleus accumbens | 0 (0/0) | 0 (0/0) | na | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | |
| Amygdaloid complex | * (*/*) | * (*/*) | 0 (0/0) | 0 (0/0) | na | + (+/++) | na | na | na | na | 0 (0/0) | 0 (0/0) | ||
| Anterior olfactory nucleus | na | 0 (0/0) | na | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (*/++++) | + (0/+) | na | ||
| Striatum | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (+++/+++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | ||
| Septal nuclei | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (+++/++++) | na | na | na | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | ||
| Ventral pallidum | na | na | 0 (0/0) | na | na | na | na | na | na | na | na | na | ||
| Diencephalon | Dorsal thalamus | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++ (+/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | |
| Hypothalamus | 0 (0/0) | + (0/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (++/++++) | na | na | na | ++++ (++++/++++) | 0 (0/0) | 0 (0/0) | ||
| Mesencephalon | Inferior colliculus | 0 (0/0) | + (*/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | + (0/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | + (0/+) | 0 (0/0) | 0 (0/0) | |
| Periaqueductal grey | + (0/+) | + (*/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | +++ (+/++++) | ++(++/++) | 0 (0/0) | ||
| Superior colliculus | 0 (0/0) | + (+/+) | 0 (0/0) | 0 (0/0) | na | ++++ (+++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | na | 0 (0/0) | 0 (0/0) | ||
| Other regions | 0 (0/0) | +++(0/+++) | 0 (0/0) | 0 (0/0) | + (0/+++) | ++++ (++/++++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | ++++ (++++/++++) | 0 (0/0) | ++ (0/++) | ||
| Rhombencephalon | Pons | 0 (0/0) | ++ (+/++) | na | 0 (0/0) | 0 (0/0) | ++++ (+++/++++) | na | na | na | ++ (+/+++) | 0 (0/0) | 0 (0/0) | |
| Cerebellum | na | + (0/+) | 0 (0/0) | 0 (0/0) | 0 (0/0) | + (+/++) | 0 (0/0) | 0 (0/0) | 0 (0/0) | 0 (0/*) | 0 (0/0) | 0 (0/0) | ||
| ANIMAL | 20F/2011 | 26F/2011 | 26C/2011 | 18A/2011 | 001/2010 | 002/2010 | p1/2011 | 26B/2011 | 26A/2011 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 7dpi | 6dpi | 6dpi | 6dpi | 6dpi | 6dpi | 6dpi | 5dpi | 4dpi | |||
| Telencephalon: | Neocortex | Auditory | + (0/++) | na | +++ (+/++++) | + (0/+++) | ++ (++/++) | ++ (++/++) | + (+/+) | + (*/+) | + (0/+) |
| Motor | + (*/+) | ++ (+/+++) | +++ (++/+++) | + (0/+) | +++ (+/++++ | + (+/+) | ++++(++++/++++) | ++++(++++/++++) | + (0/++) | ||
| Prefrontal | + (*/+) | + (+/++) | + (+/++) | 0 (0/0) | ++++(++++/++++) | + (+/+) | +++ (++/+++) | na | + (*/++) | ||
| Retrosplenial | + (+/+) | +++ (++/++++) | + (0/++) | + (0/+) | ++ (++/+++) | ++ (++/++) | ++++(++++/++++) | ++ (+/++++) | + (+/++) | ||
| Somatosensory | + (0/+) | + (+/+) | +++ (0/++++) | ++ (0/+++) | ++++ (++/++++) | * (*/*) | ++++(++++/++++) | +++ (+/++++) | + (0/++) | ||
| Visual | + (0/+) | + (+/+) | + (0/++) | ++(+/++) | ++ (++/++) | * (*/*) | na | +++ (+/++++) | +++(+++/+++) | ||
| Piriform | + (+/+) | + (+/+) | + (0/+) | + (+/++) | ++ (+/+++) | + (+/++) | ++ (++/++) | ++ (++/++) | + (+/+) | ||
| Hippoc. | CA1 | ++ (*/++) | + (0/+) | ++ (+/+++) | ++ (0/+++) | +++ (+++/+++) | + (+/+) | ++++(++++/++++) | 0 (0/0) | + (*/+) | |
| CA3 | ++ (+/++) | ++ (+/++) | ++ (+/+++) | + (0/++) | +++ (+++/+++) | + (+/+) | ++ (++/++) | 0 (*/0) | ++ (*/+++) | ||
| Dentate gyrus | + (*/+) | + (+/+) | + (+/+) | + (0/+) | + (+/+) | + (+/+) | + (+/+) | + (0/+) | + (+/+) | ||
| Subcortical nuclei | Nucleus accumbens | + (0/++) | + (+/+) | + (+/+) | * (*/*) | 0 (0/0) | + (*/+) | + (+/+) | + (+/+) | 0 (0/0) | |
| Amygdaloid complex | * (0/*) | na | + (+/+) | na | na | +++(+++/++++) | + (+/+) | 0 (0/0) | 0 (0/0) | ||
| Anterior olfactory nucleus | na | + (+/+) | na | 0 (0/0) | na | na | na | na | + (0/+) | ||
| Striatum | + (*/+) | + (+/+) | + (0/+) | + (*/+) | + (0/++) | + (+/+++) | + (+/++) | + (0/+) | * (*/*) | ||
| Septal nuclei | ++ (+/++) | na | + (+/+) | ++ (++/+++) | +++ (+++/+++) | na | + (+/+) | +++ (++/++++) | 0 (0/0) | ||
| Ventral pallidum | na | na | na | na | na | na | na | na | 0 (0/0) | ||
| Diencephalon | Dorsal thalamus | + (+/+) | na | + (0/+) | + (0/++) | + (+/+) | + (+/+++) | ++ (+/++) | ++ (*/++++) | * (0/*) | |
| Hypothalamus | + (+/+) | + (+/+) | + (0/++) | 0 (0/0) | +++ (+/++++) | ++ (+/+++) | ++ (0/++) | + (0/+) | |||
| Mesencephalon | Inferior colliculus | 0 (0/0) | + (+/+) | + (*/+) | 0 (0/0) | na | na | na | 0 (0/0) | * (*/*) | |
| Periaqueductal grey | ++ (+/+++) | +++ (++/++++) | ++ (+/++) | ++ (++/++) | ++ (+/++0 | + (+/++) | + (0/+) | + (*/++) | ++ (*/+++) | ||
| Superior colliculus | 0 (*/0) | ++++(++++/++++) | 0 (0/0) | * (*/*) | + (+/++) | + (+/*) | 0 (0/0) | + (*/+) | + (+/+) | ||
| Other regions | ++ (+/++) | ++++ (+++/++++) | ++ (++/++++) | +++ (*/+++) | ++ (+/+++) | ++ (+/+++) | * (0/*) | +++ (++/++++) | * (*/*) | ||
| Rhombencephalon | Pons | 0 (*/0) | +++ (+/++++) | + (+/+) | +++ (+++/+++) | na | na | na | ++++(++++/++++) | + (+/+) | |
| Cerebellum | * (*/0) | ++ (+/++) | + (*/+) | 0 (0/0) | + (+/+) | 0 (0/0) | + (+/+) | 0 (0/0) | na | ||
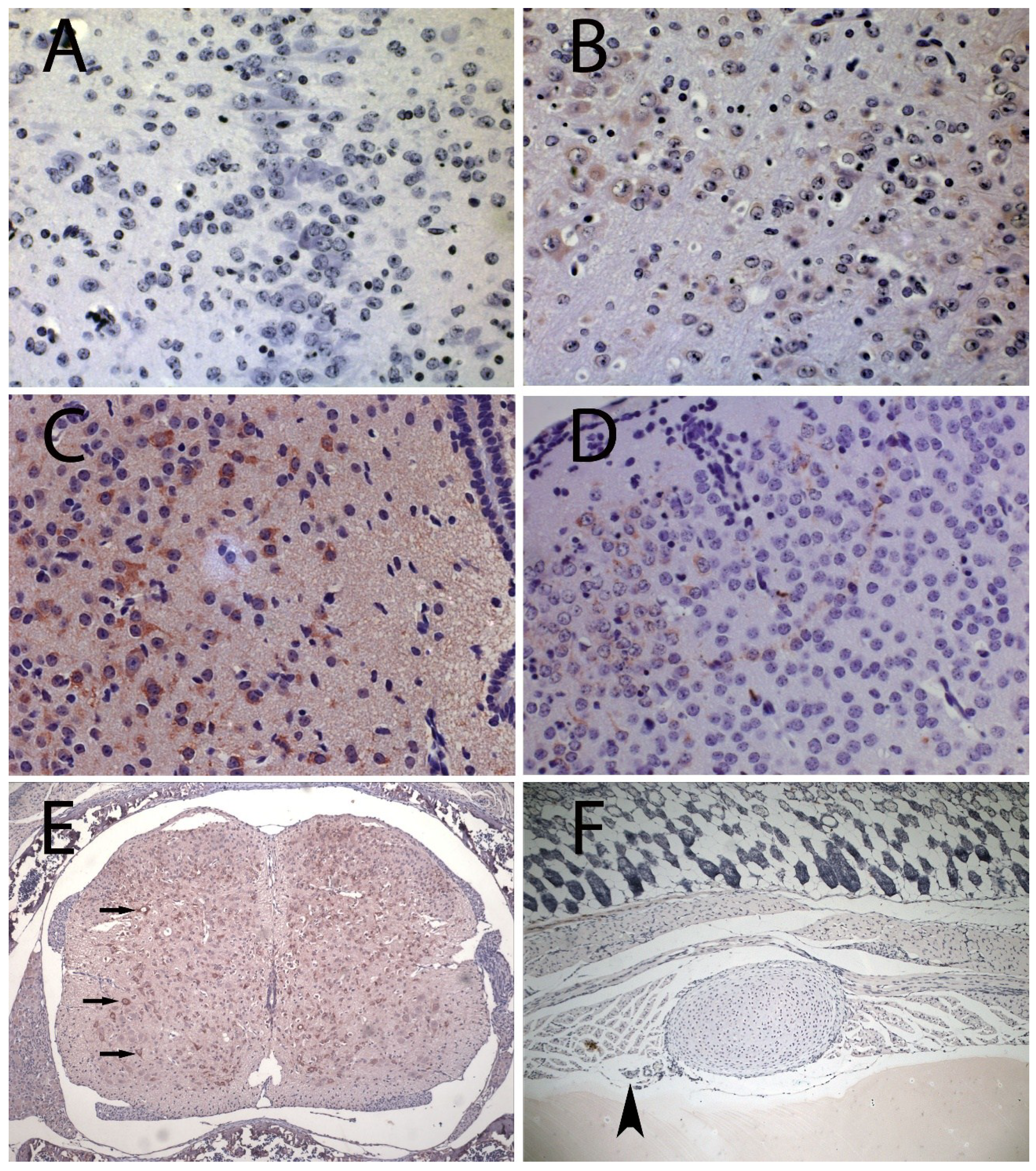
3. Results and Discussion

Supplementary Files
Supplementary File 1Author Contributions
Conflicts of Interest
References and Notes
- Nichol, S. Bunyaviridae. In Fields Virology, 4th ed.; Howley, K., Ed.; Lippincott Willians & Wilkins: Philadelphia, PA, USA, 2001; Volume 2. [Google Scholar]
- Pinheiro, F.P.; Travassos da Rosa, A.P.; Travassos da Rosa, J.F.; Ishak, R.; Freitas, R.B.; Gomes, M.L.; LeDuc, J.W.; Oliva, O.F. Oropouche virus. I. A review of clinical, epidemiological, and ecological findings. Am. J. Trop. Med. Hyg. 1981, 30, 149–160. [Google Scholar]
- Bastos, M.S.; Lessa, N.; Naveca, F.G.; Monte, R.L.; Braga, W.S.; Figueiredo, L.T.; Ramasawmy, R.; Mourao, M.P. Detection of Herpesvirus, Enterovirus, and Arbovirus infection in patients with suspected central nervous system viral infection in the Western Brazilian Amazon. J. Med. Virol. 2014, 86, 1522–1527. [Google Scholar]
- Bastos, M.E.S.; Figueiredo, L.T.; Naveca, F.G.; Monte, R.L.; Lessa, N.; Pinto de Figueiredo, R.M.; Gimaque, J.B.; Pivoto João, G.; Ramasawmy, R.; Mourão, M.P. Identification of Oropouche Orthobunyavirus in the cerebrospinal fluid of three patients in the Amazonas, Brazil. Am. J. Trop. Med. Hyg. 2012, 86, 732–735. [Google Scholar]
- LeDuc, J.W.; Hoch, A.L.; Pinheiro, F.P.; da Rosa, A.P. Epidemic Oropouche virus disease in northern Brazil. Bull. Pan Am. Health Organ. 1981, 15, 97–103. [Google Scholar]
- Pinheiro, F.P.; Rocha, A.G.; Freitas, R.B.; Ohana, B.A.; Travassos da Rosa, A.P.; Rogério, J.S.; Linhares, A.C. Meningitis associated with Oropouche virus infections. Rev. Inst. Med. Trop. Sao Paulo 1982, 24, 246–251. [Google Scholar]
- Watts, D.M.; Lavera, V.; Callahan, J.; Rossi, C.; Oberste, M.S.; Roehrig, J.T.; Cropp, C.B.; Karabatsos, N.; Smith, J.F.; Gubler, D.J.; et al. Venezuelan equine encephalitis and Oropouche virus infections among Peruvian army troops in the Amazon region of Peru. Am. J. Trop. Med. Hyg. 1997, 56, 661–667. [Google Scholar]
- Araujo, R.; Dias, L.B.; Araujo, M.T.; Pinheiro, F.; Oliva, O.F. Ultrastructural changes in the hamster liver after experimental inoculation with Oropouche arbovirus (type BeAn 19991). Rev. Inst. Med. Trop. Sao Paulo 1978, 20, 45–54. [Google Scholar]
- Pinheiro, F.P.; Travassos da Rosa, A.P.; Travassos da Rosa, J.F.; Bensabath, G. An outbreak of Oropouche virus diease in the vicinity of santarem, para, barzil. Tropenmedizin und Parasitologie 1976, 27, 213–223. [Google Scholar]
- Santos, R.I.; Almeida, M.F.; Paula, F.E.; Rodrigues, A.H.; Saranzo, A.M.; Paula, A.E.; Silva, M.L.; Correa, V.M.; Acrani, G.O.; Neder, L.; et al. Experimental infection of suckling mice by subcutaneous inoculation with Oropouche virus. Virus Res. 2012, 170, 25–33. [Google Scholar]
- Rodrigues, A.H.; Santos, R.I.; Arisi, G.M.; Bernardes, E.S.; Silva, M.L.; Rossi, M.A.; Lopes, M.B.; Arruda, E. Oropouche virus experimental infection in the golden hamster (Mesocrisetus auratus). Virus Res. 2011, 155, 35–41. [Google Scholar]
- Acrani, G.O.; Gomes, R.; Proenca-Modena, J.L.; da Silva, A.F.; Carminati, P.O.; Silva, M.L.; Marques Santos, R.I.; Arruda, E. Apoptosis induced by Oropouche virus infection in HeLa cells is dependent on virus protein expression. Virus Res. 2010, 149, 56–63. [Google Scholar]
- Luna, L. Manual of Histologic Staining: Methods of the Armed Forces Institute of Pathology, 3rd ed.; Mcgraw-Hill: New York, NY, USA, 1968. [Google Scholar]
- Allen Institute for Brain Science. Available online: http://mouse.brain-map.org/static/atlas (accessed on 6 August 2014).
- Steiner, I.; Kennedy, P.G.; Pachner, A.R. The neurotropic herpes viruses: Herpes simplex and varicella-zoster. Lancet Neurol. 2007, 6, 1015–1028. [Google Scholar]
- Schnell, M.J.; McGettigan, J.P.; Wirblich, C.; Papaneri, A. The cell biology of rabies virus: Using stealth to reach the brain. Nat. Rev. Microbiol. 2010, 8, 51–61. [Google Scholar]
- Spindler, K.R.; Hsu, T.H. Viral disruption of the blood-brain barrier. Trends Microbiol. 2012, 20, 282–290. [Google Scholar]
- Dodd, K.A.; McElroy, A.K.; Jones, T.L.; Zaki, S.R.; Nichol, S.T.; Spiropoulou, C.F. Rift valley Fever virus encephalitis is associated with an ineffective systemic immune response and activated T cell infiltration into the CNS in an immunocompetent mouse model. PLoS Negl. Trop. Dis. 2014, 8, e2874. [Google Scholar] [CrossRef]
- Talamonti, L.; Padula, P.J.; Canteli, M.S.; Posner, F.; Marczeski, F.P.; Weller, C. Hantavirus pulmonary syndrome: Encephalitis caused by virus Andes. J. Neurovirol. 2011, 17, 189–192. [Google Scholar]
- Huisa, B.N.; Chapin, J.E.; Adair, J.C. Central nervous system complications following Hanta virus cardiopulmonary syndrome. J. Neurovirol. 2009, 15, 202–205. [Google Scholar]
- Bergmann, F.; Krone, B.; Bleich, S.; Prange, H.; Paulus, W. Encephalitis due to a hantavirus infection. J. Infect. 2002, 45, 58–59. [Google Scholar]
- Ruzek, D.; Salat, J.; Singh, S.K.; Kopecky, J. Breakdown of the blood-brain barrier during tick-borne encephalitis in mice is not dependent on CD8+ T-cells. PLoS One 2011, 6, e20472. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, R.I.; Bueno-Júnior, L.S.; Ruggiero, R.N.; Almeida, M.F.; Silva, M.L.; Paula, F.E.; Correa, V.M.A.; Arruda, E. Spread of Oropouche Virus into the Central Nervous System in Mouse. Viruses 2014, 6, 3827-3836. https://doi.org/10.3390/v6103827
Santos RI, Bueno-Júnior LS, Ruggiero RN, Almeida MF, Silva ML, Paula FE, Correa VMA, Arruda E. Spread of Oropouche Virus into the Central Nervous System in Mouse. Viruses. 2014; 6(10):3827-3836. https://doi.org/10.3390/v6103827
Chicago/Turabian StyleSantos, Rodrigo I., Lézio S. Bueno-Júnior, Rafael N. Ruggiero, Mariana F. Almeida, Maria L. Silva, Flávia E. Paula, Vani M. A. Correa, and Eurico Arruda. 2014. "Spread of Oropouche Virus into the Central Nervous System in Mouse" Viruses 6, no. 10: 3827-3836. https://doi.org/10.3390/v6103827
APA StyleSantos, R. I., Bueno-Júnior, L. S., Ruggiero, R. N., Almeida, M. F., Silva, M. L., Paula, F. E., Correa, V. M. A., & Arruda, E. (2014). Spread of Oropouche Virus into the Central Nervous System in Mouse. Viruses, 6(10), 3827-3836. https://doi.org/10.3390/v6103827