First Isolation of a Giant Virus from Wild Hirudo medicinalis Leech: Mimiviridae isolation in Hirudo medicinalis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
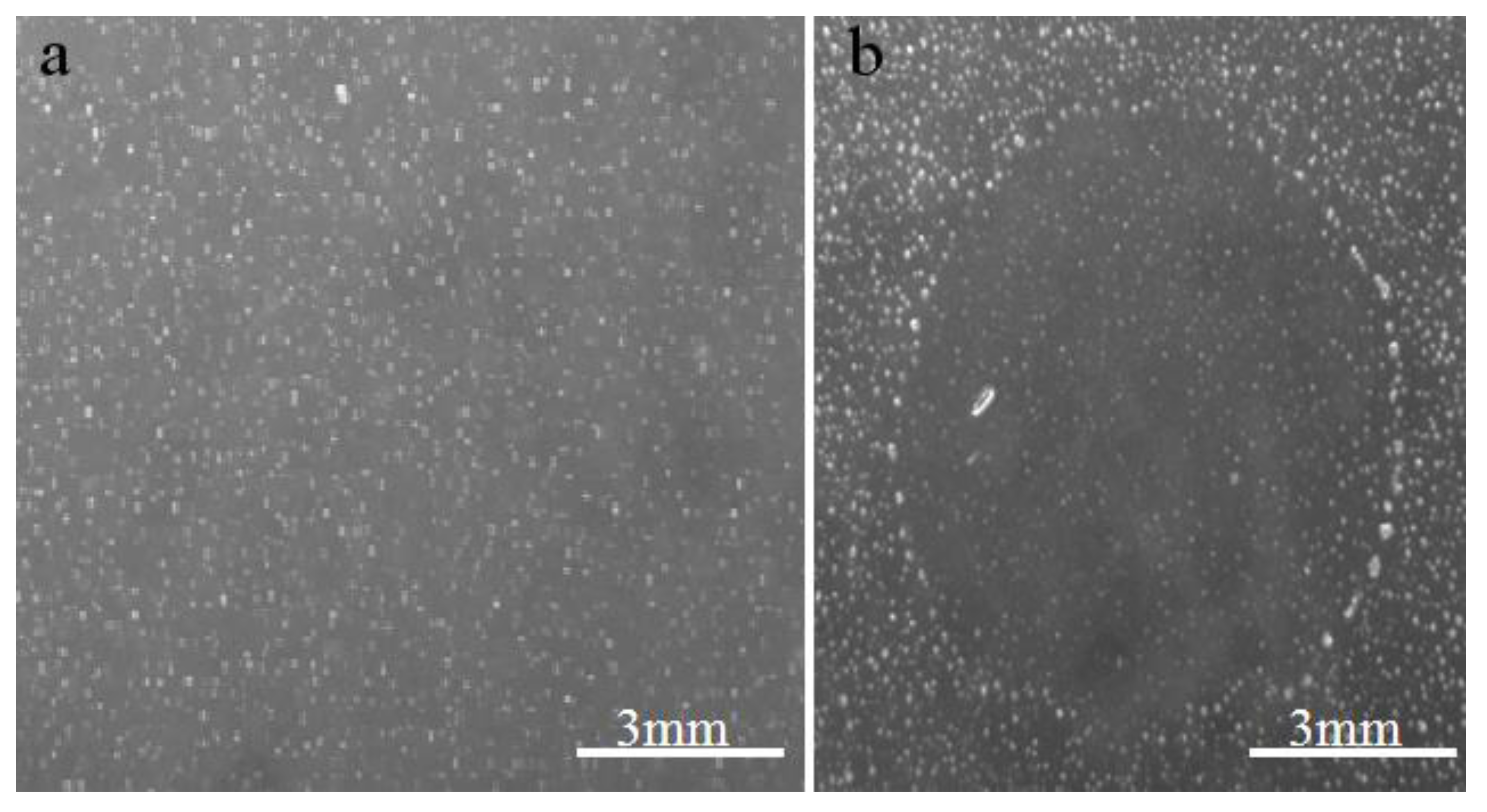
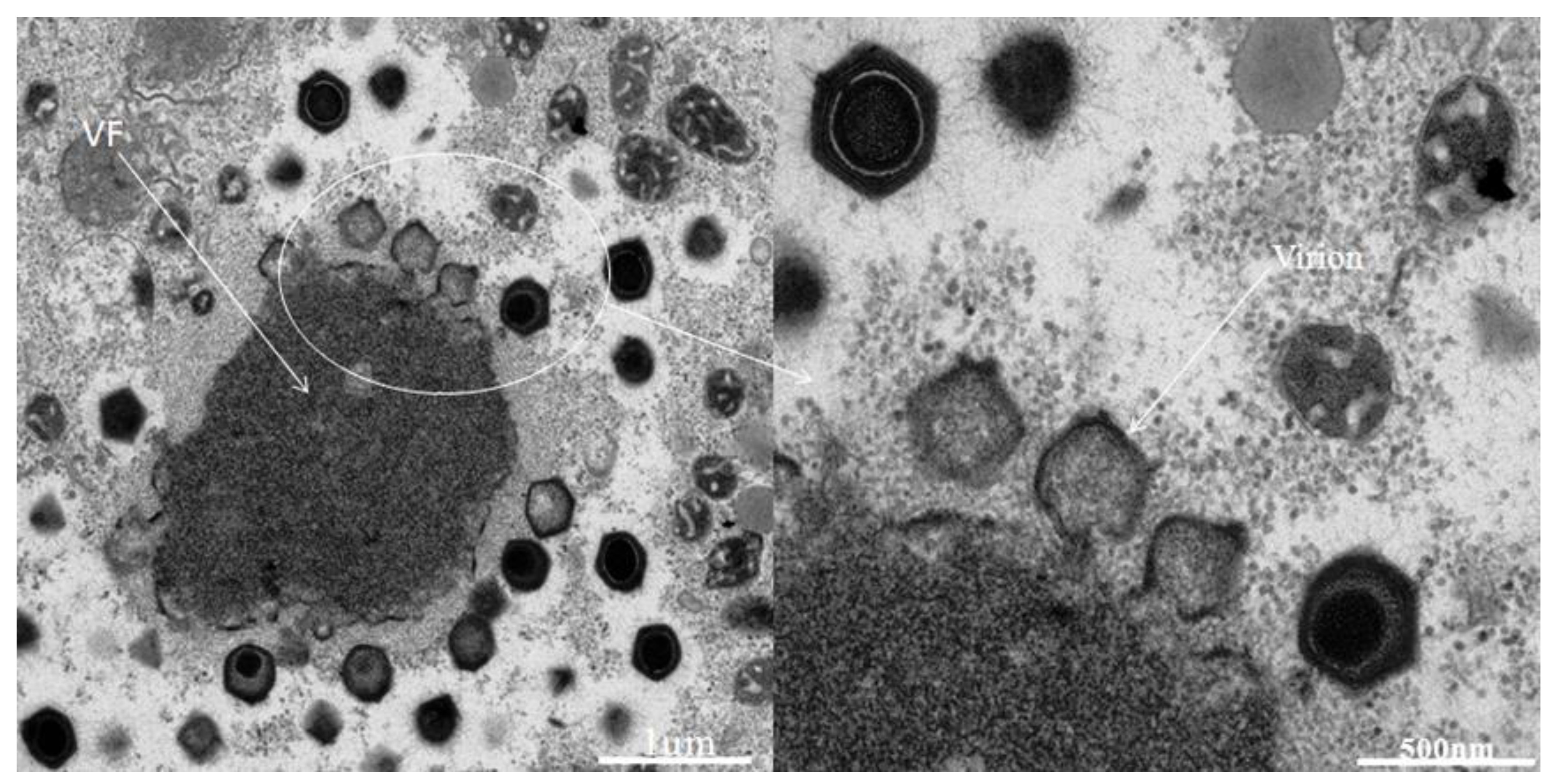
2.1. Virus isolation and preliminary characterization


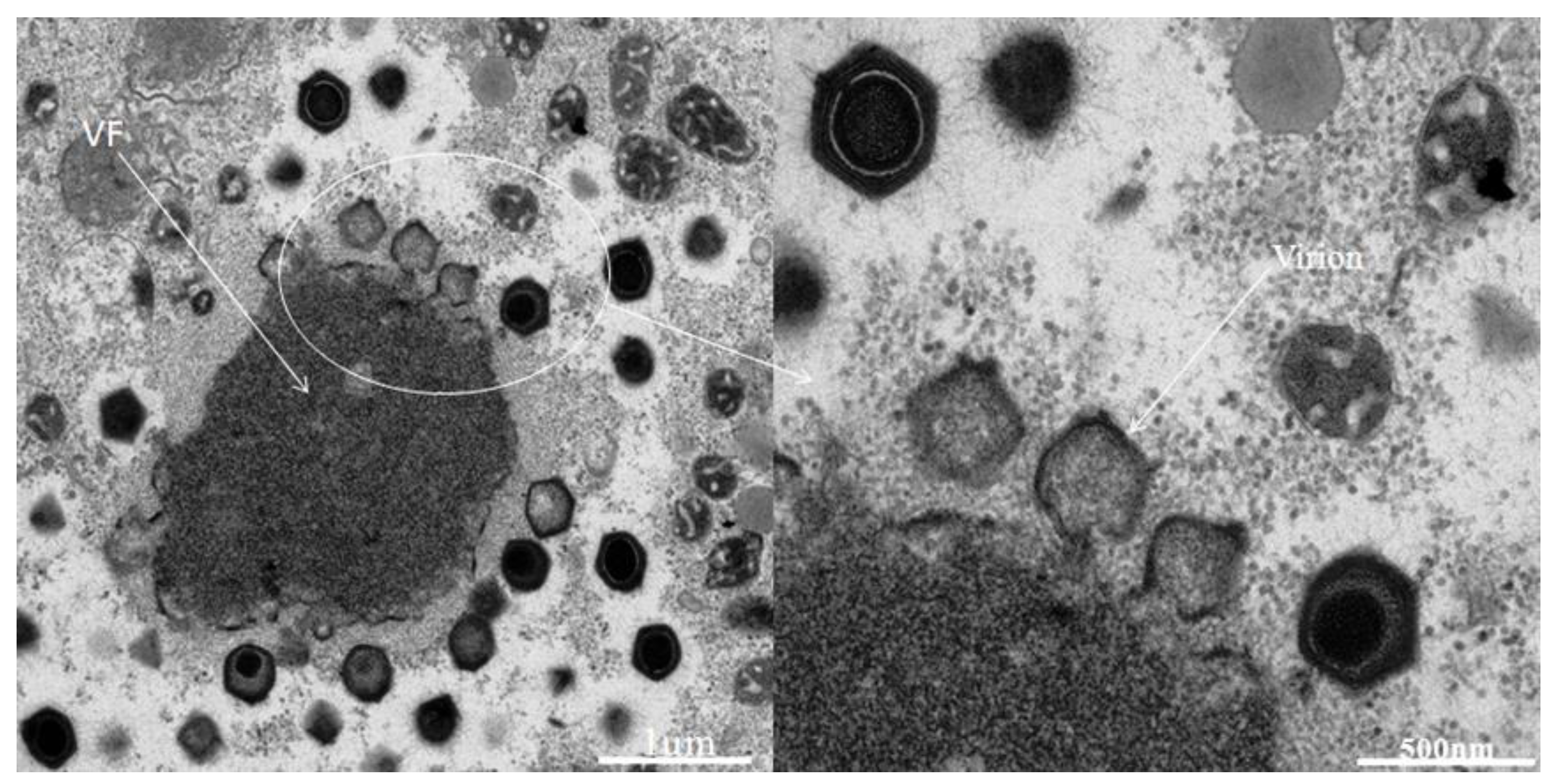
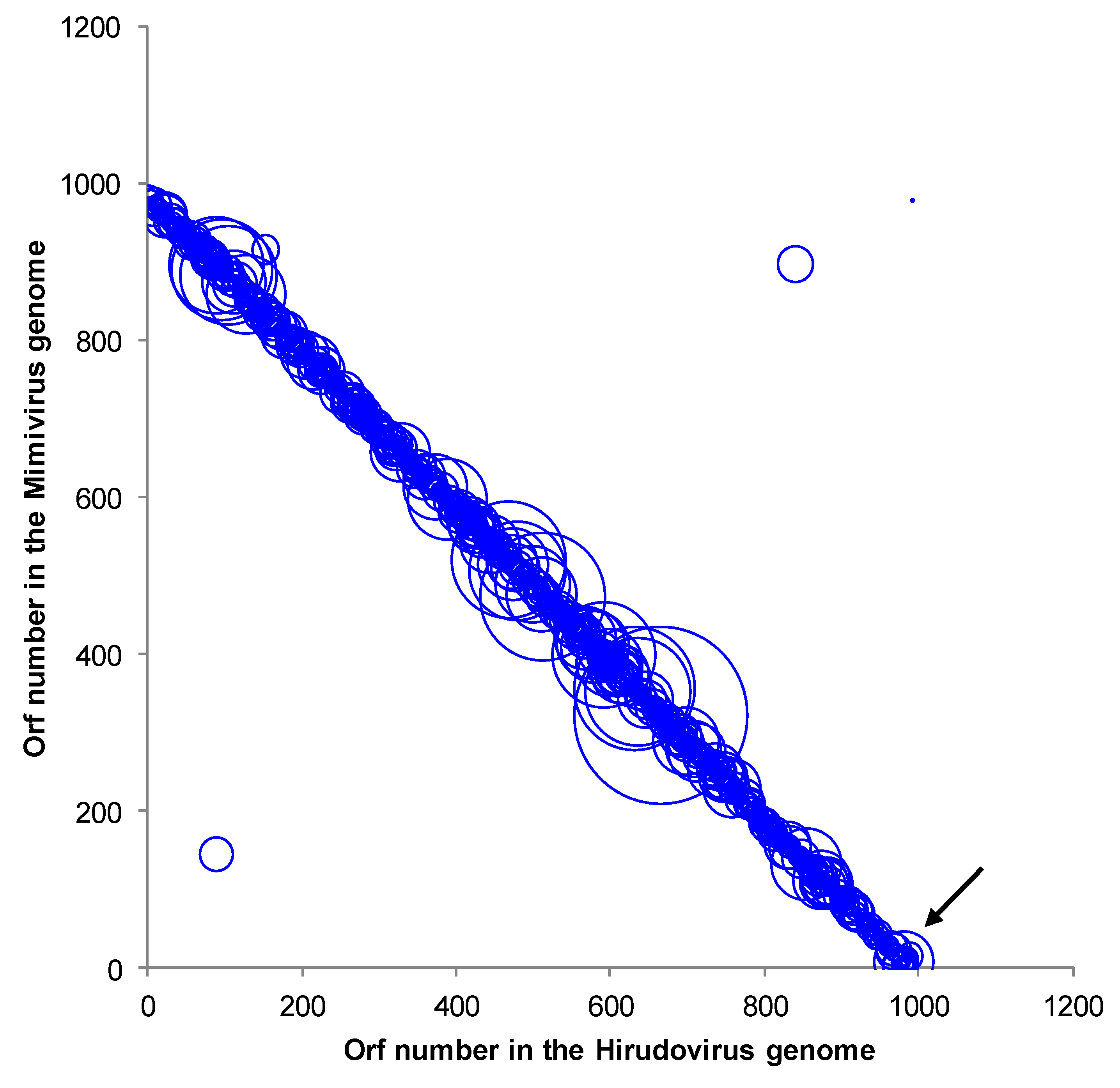
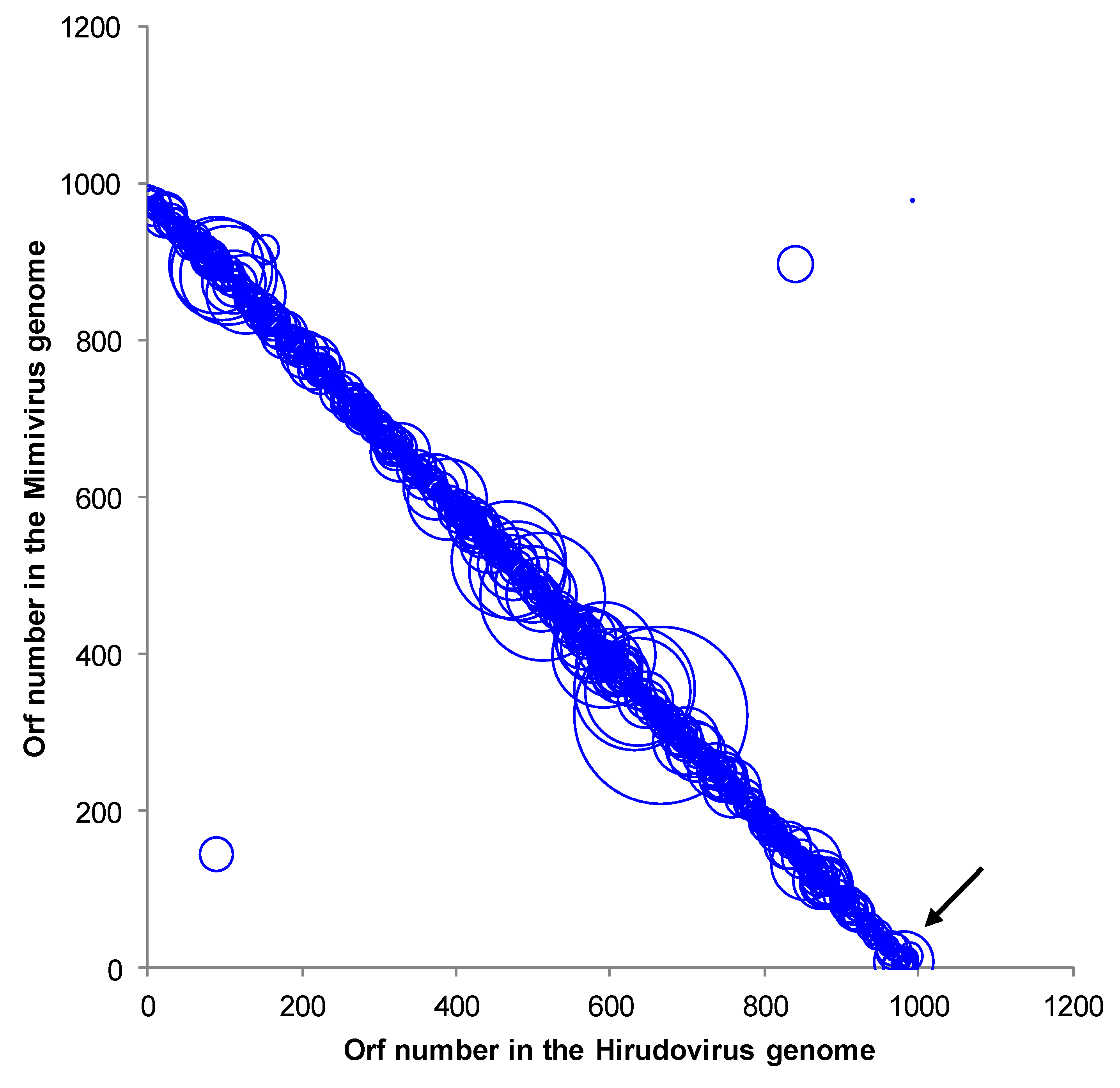
2.2. Genome sequencing, annotation and comparative genomics
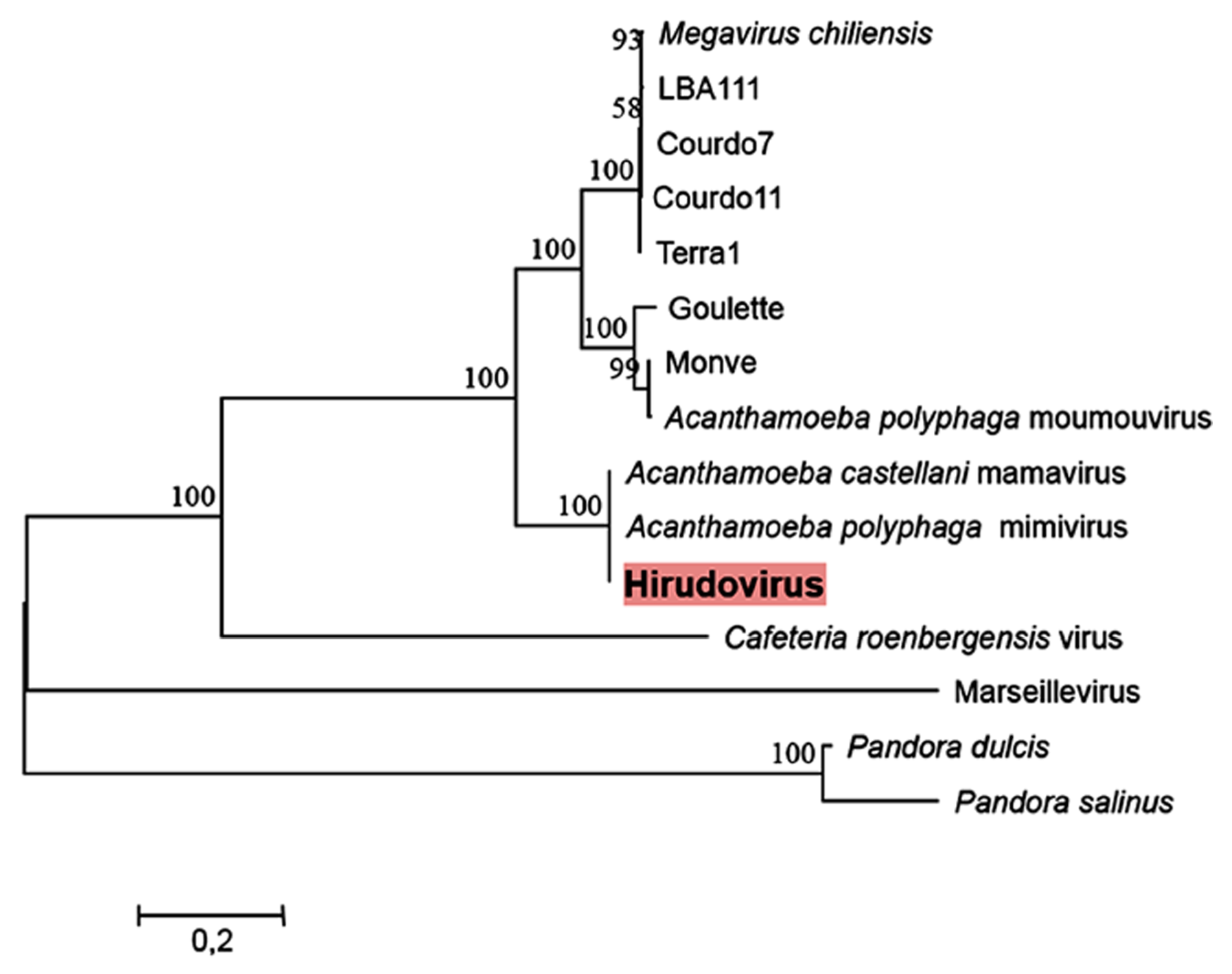
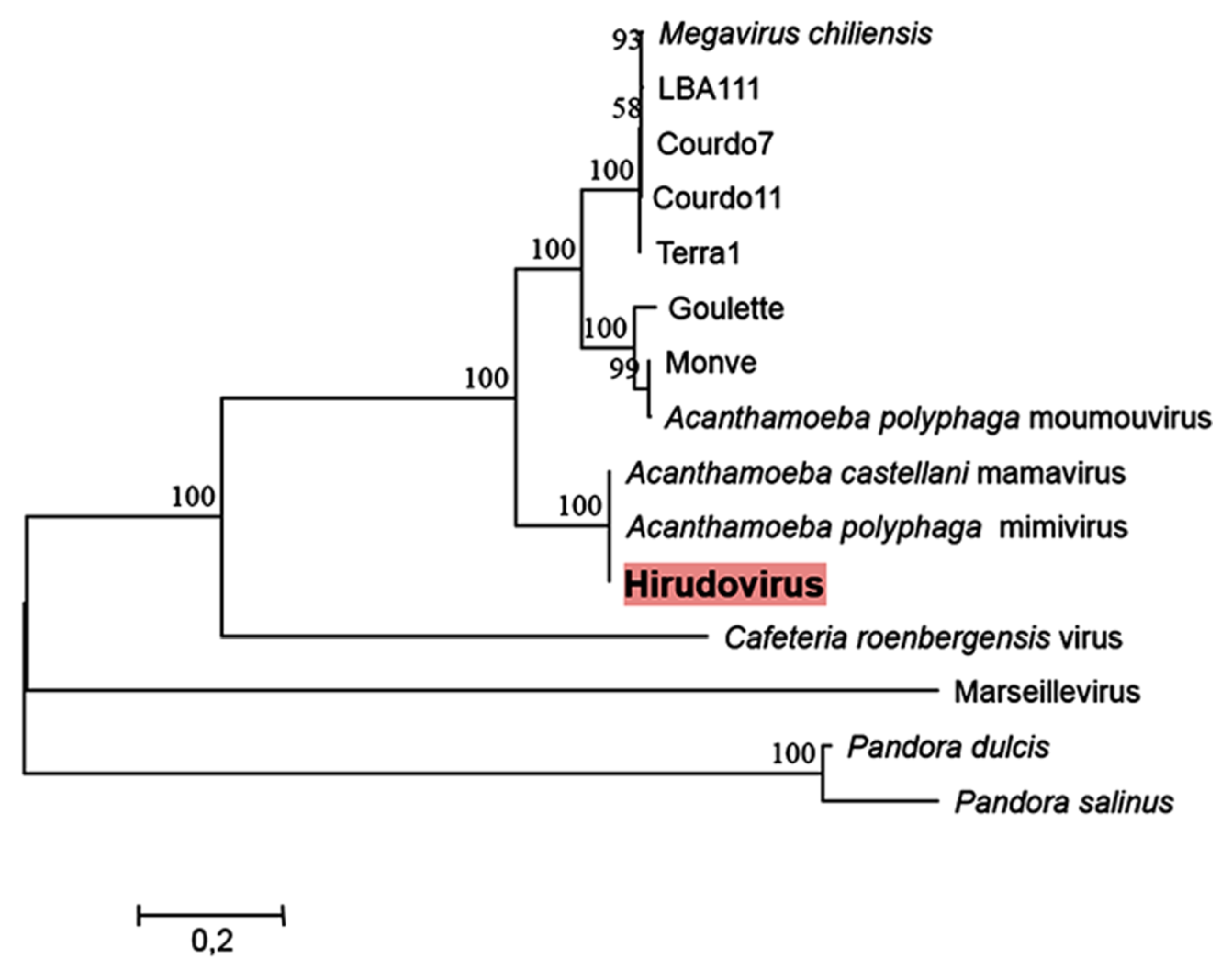
2.3. Phylogeny
3. Discussion
4. Materials and methods
Conflicts of Interest
References
- Fields, W.S. The history of leeching and hirudin. Haemostasis 1991, 21, 3–10. [Google Scholar]
- Baillie, J.E.M.; Hilton-Taylor, C.; Stuart, S.N. A Global Species Assessment; IUCN: Gland, Switzerland and Cambridge, UK; Red List of Threatened Species, Ed.; 2004; pp. 1–191. [Google Scholar]
- Nelson, M.C.; Graf, J. Bacterial symbioses of the medicinal leech Hirudo verbana. Gut. Microbe. 2012, 3, 322–331. [Google Scholar] [CrossRef]
- Whitaker, I.S.; Oboumarzouk, O.; Rozen, W.M.; Naderi, N.; Balasubramanian, S.P.; Azzopardi, E.A.; Kon, M. The efficacy of medicinal leeches in plastic and reconstructive surgery: a systematic review of 277 reported clinical cases. Microsurgery 2012, 32, 240–249. [Google Scholar] [CrossRef]
- Maetz, B.; Abbou, R.; Andreoletti, J.B.; Bruant-Rodier, C. Infections following the application of leeches: two case reports and review of the literature. J. Med. Case Rep. 2012, 25, 364. [Google Scholar]
- Sartor, C.; Bornet, C.; Guinard, D.; Fournier, P.E. Transmission of Aeromonas hydrophila by leeches. Lancet 2013, 11, 1686. [Google Scholar]
- Kikuchi, Y.; Bomar, L.; Graf, J. Stratified bacterial community in the bladder of the medicinal leech, Hirudo verbana. Environ. Microbiol. 2009, 11, 2758–2770. [Google Scholar] [CrossRef]
- Bomar, L.; Maltz, M.; Colston, S.; Graf, J. Directed culturing of microorganisms using metatranscriptomics. MBio. 2011, 5, 2. [Google Scholar]
- Shope, R.E. The leech as a potential virus reservoir. J. Exp. Med. 1957, 105, 373–382. [Google Scholar]
- Al-Khleif, A.; Roth, M.; Menge, C.; Heuser, J.; Baljer, G; Herbst, W. Tenacity of mammalian viruses in the gut of leeches fed with porcine blood. J. Med. Microbiol. 2011, 60, 787–792. [Google Scholar] [CrossRef]
- Nehili, M.; Ilk, C.; Mehlhorn, H.; Ruhnau, K.; Dick, W.; Njayou, M. Experiments on the possible role of leeches as vectors of animal and human pathogens: a light and electron microscopy study. Parasitol. Res. 1994, 80, 277–290. [Google Scholar]
- Pagnier, I.; Ikanga Reteno, D.G.; Saadi, H.; Boughalmi, M.; Gaia, M.; Slimani, M.; Nto, T.; Bekliz, M.; Colson, P.; Raoult, D.; La Scola, B. A decade of improvements in Mimiviridae and Marseilleviridae isolation from amoeba. Intervirology 2013, (in press). [Google Scholar]
- Colson, P.; de Lamballerie, X.; Fournous, G.; Raoult, D. Reclassification of giant viruses composing a fourth domain of life in the new order Megavirales. Intervirology 2012, 55, 321–332. [Google Scholar] [CrossRef]
- Raoult, D.; Audic, S.; Robert, C.; Abergel, C.; Renesto, P.; Ogata, H.; La Scola, B.; Suzan, M.; Claverie, J.M. The 1.2-megabase genome sequence of Mimivirus. Science 2004, 306, 1344–1350. [Google Scholar]
- Colson, P.; Yutin, N.; Shabalina, S.A.; Robert, C.; Fournous, G.; La Scola, B.; Raoult, D.; Koonin, E.V. Viruses with more than 1,000 genes: Mamavirus, a new Acanthamoeba polyphaga mimivirus strain, and reannotation of Mimivirus genes. Genome Biol. Evol. 2011, 3, 737–742. [Google Scholar] [CrossRef]
- Arslan, D.; Legendre, M.; Seltzer, V.; Abergel, C.; Claverie, J.M. Distant Mimivirus relative with a larger genome highlights the fundamental features of Megaviridae. Proc. Natl. Acad. Sci. U.S.A. 2011, 108, 17486–17491. [Google Scholar]
- Saadi, H.; Pagnier, I.; Colson, P.; Cherif, J.K.; Beji, M.; Boughalmi, M.; Azza, S.; Armstrong, N.; Robert, C.; Fournous, G.; La Scola, B.; Raoult, D. First isolation of mimivirus in a patient with pneumonia. Clin. Infect. Dis. 2013, 57, 127–134. [Google Scholar]
- Yoosuf, N.; Yutin, N.; Colson, P.; Shabalina, S.A.; Pagnier, I.; Robert, C.; Azza, S.; Klose, T.; Wong, J.; Rossmann, M.G.; La Scola, B.; Raoult, D.; Koonin, E.V. Related giant viruses in distant locations and different habitats: Acanthamoeba polyphaga moumouvirus represents a third lineage of the Mimiviridae that is close to the megavirus lineage. Genome. Biol. Evol. 2012 4, 1324–1330.
- Cohen, G.; Hoffart, L.; La Scola, B.; Raoult, D.; Drancourt, M. Ameba-associated keratitis, France. Emerg. Infect. Dis. 2011, 17, 1306–1308. [Google Scholar]
- Boughalmi, M.; Pagnier, I.; Aherfi, S.; Colson, P.; Raoult, D.; La Scola, B. First isolation of a giant virus, Insectomime, in the Diptera Syrphidae Eristalis tenax. Intervirology 2013, (in press). [Google Scholar]
- Boughalmi, M.; Saadi, H.; Pagnier, I.; Colson, P.; Fournous, G.; Raoult, D.; La Scola, B. High-throughput isolation of giant viruses of the Mimiviridae and Marseilleviridae families in the Tunisian environment. Environ. Microbiol. 2013, 15, 2000–2007. [Google Scholar]
- Boughlami, M.; Pagnier, I.; Aherfi, S.; Colson, P.; Raoult, D.; La Scola, B. First isolation of Marseillevirus in the diptera Syrphidae Eristalis tenax. Intervirology 2013, 56, 386–394. [Google Scholar]
- Boyer, M.; Yutin, N.; Pagnier, I.; Barrassi, L.; Fournous, G.; Espinosa, L.; Robert, C.; Azza, S.; Sun, S.; Rossmann, M.G.; Suzan-Monti, M.; La Scola, B.; Koonin, E.V.; Raoult, D. Giant Marseillevirus highlights the role of amoebae as a melting pot in emergence of chimeric microorganisms. Proc. Natl. Acad. Sci. U.S.A. 2009, 106, 21848–21853. [Google Scholar]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.J.; Chen, Z.; Dewell, S.B.; Du, L.; Fierro, J.M.; Gomes, X.V.; Godwin, B.C.; He, W.; Helgesen, S.; Ho, C.H.; Irzyk, G.P.; Jando, S.C.; Alenquer, M.L.; Jarvie, T.P.; Jirage, K.B.; Kim, J.B.; Knight, J.R.; Lanza, J.R.; Leamon, J.H.; Lefkowitz, S.M.; Lei, M.; Li, J.; Lohman, K.L.; Lu, H.; Makhijani, V.B.; McDade, K.E.; McKenna, M.P.; Myers, E.W.; Nickerson, E.; Nobile, J.R.; Plant, R.; Puc, B.P.; Ronan, M.T.; Roth, G.T.; Sarkis, G.J.; Simons, J.F.; Simpson, J.W.; Srinivasan, M.; Tartaro, K.R.; Tomasz, A.; Vogt, K.A.; Volkmer, G.A.; Wang, S.H.; Wang, Y.; Weiner, M.P.; Yu, P.; Begley, R.F.; Rothberg, J.M. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar]
- Besemer, J.; Borodovsky, M. GeneMark: web software for gene finding in prokaryotes, eukaryotes and viruses. Nucleic. Acids. Res. 2005, 33, 451–454. [Google Scholar] [CrossRef]
- La Scola, B.; Audic, S.; Robert, C.; Jungang, L.; de Lamballerie, X.; Drancourt, M.; Birtles, R.; Claverie, J.M.; Raoult, D. A giant virus in amoebae. Science 2003, 299, 2033. [Google Scholar]
- Philippe, N.; Legendre, M.; Doutre, G.; Couté, Y.; Poirot, O.; Lescot, M.; Arslan, D.; Seltzer, V.; Bertaux, L.; Bruley, C.; Garin, J.; Claverie, J.M.; Abergel, C. Pandoraviruses: amoeba viruses with genomes up to 2.5 Mb reaching that of parasitic eukaryotes. Science 2013, 341, 281–286. [Google Scholar] [CrossRef]
- Lechner, M.; Findeiss, S.; Steiner, L.; Marz, M.; Stadler, P.F.; Prohaska, S.J. Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinformatics 2011, 12, 124. [Google Scholar]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic. Acids. Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic. Acids. Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Boughalmi, M.; Pagnier, I.; Aherfi, S.; Colson, P.; Raoult, D.; La Scola, B. First Isolation of a Giant Virus from Wild Hirudo medicinalis Leech: Mimiviridae isolation in Hirudo medicinalis. Viruses 2013, 5, 2920-2930. https://doi.org/10.3390/v5122920
Boughalmi M, Pagnier I, Aherfi S, Colson P, Raoult D, La Scola B. First Isolation of a Giant Virus from Wild Hirudo medicinalis Leech: Mimiviridae isolation in Hirudo medicinalis. Viruses. 2013; 5(12):2920-2930. https://doi.org/10.3390/v5122920
Chicago/Turabian StyleBoughalmi, Mondher, Isabelle Pagnier, Sarah Aherfi, Philippe Colson, Didier Raoult, and Bernard La Scola. 2013. "First Isolation of a Giant Virus from Wild Hirudo medicinalis Leech: Mimiviridae isolation in Hirudo medicinalis" Viruses 5, no. 12: 2920-2930. https://doi.org/10.3390/v5122920
APA StyleBoughalmi, M., Pagnier, I., Aherfi, S., Colson, P., Raoult, D., & La Scola, B. (2013). First Isolation of a Giant Virus from Wild Hirudo medicinalis Leech: Mimiviridae isolation in Hirudo medicinalis. Viruses, 5(12), 2920-2930. https://doi.org/10.3390/v5122920