A VSV-G Pseudotyped Last Generation Lentiviral Vector Mediates High Level and Persistent Gene Transfer in Models of Airway Epithelium In Vitro and In Vivo


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
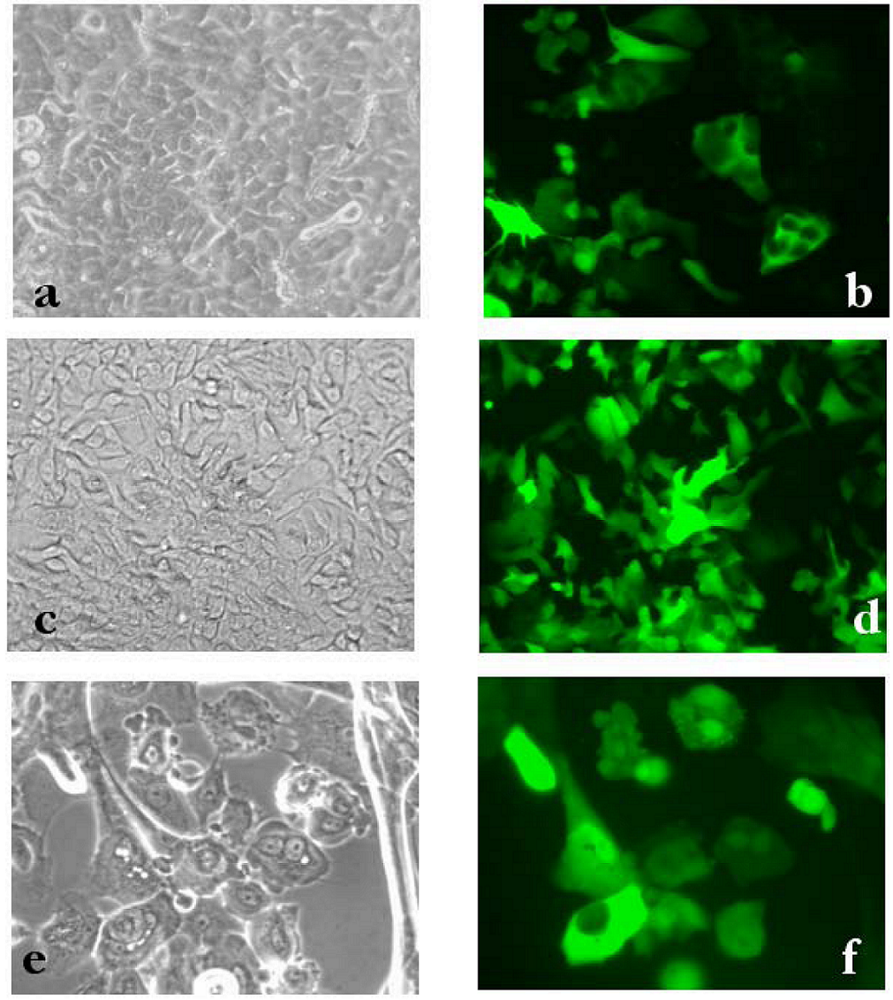
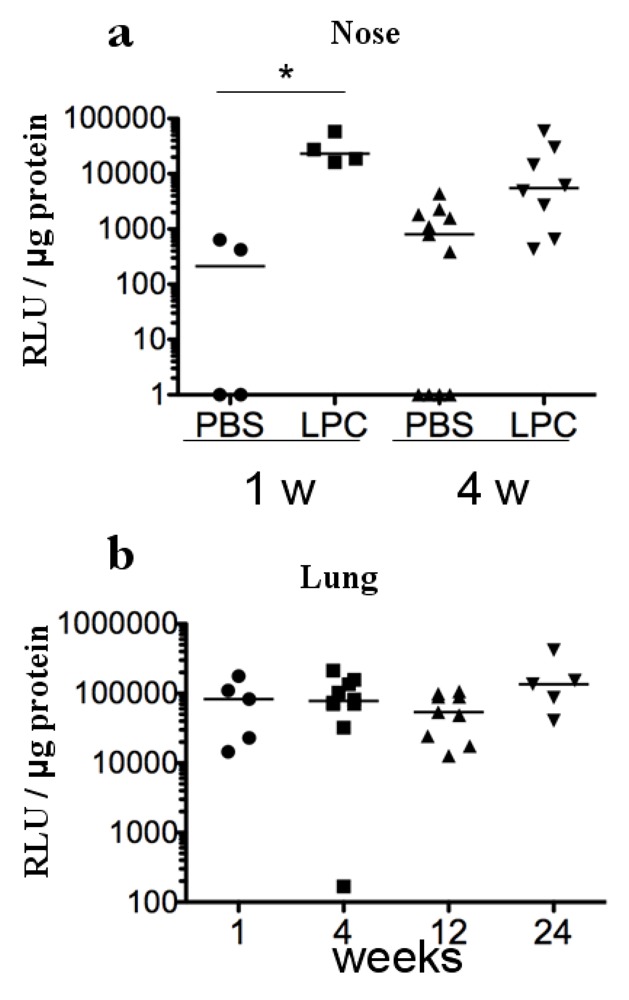
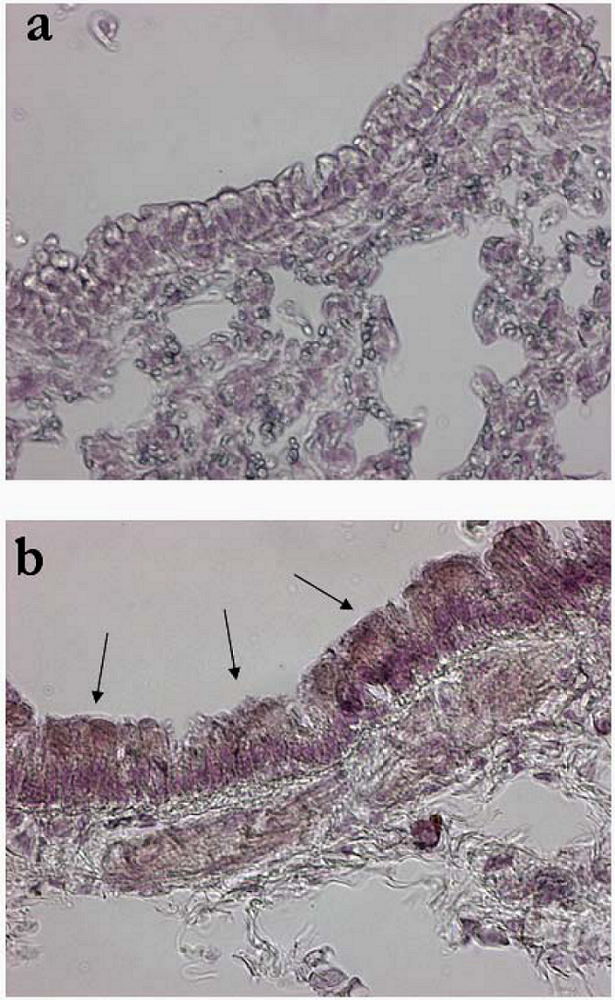
2. Results and Discussion
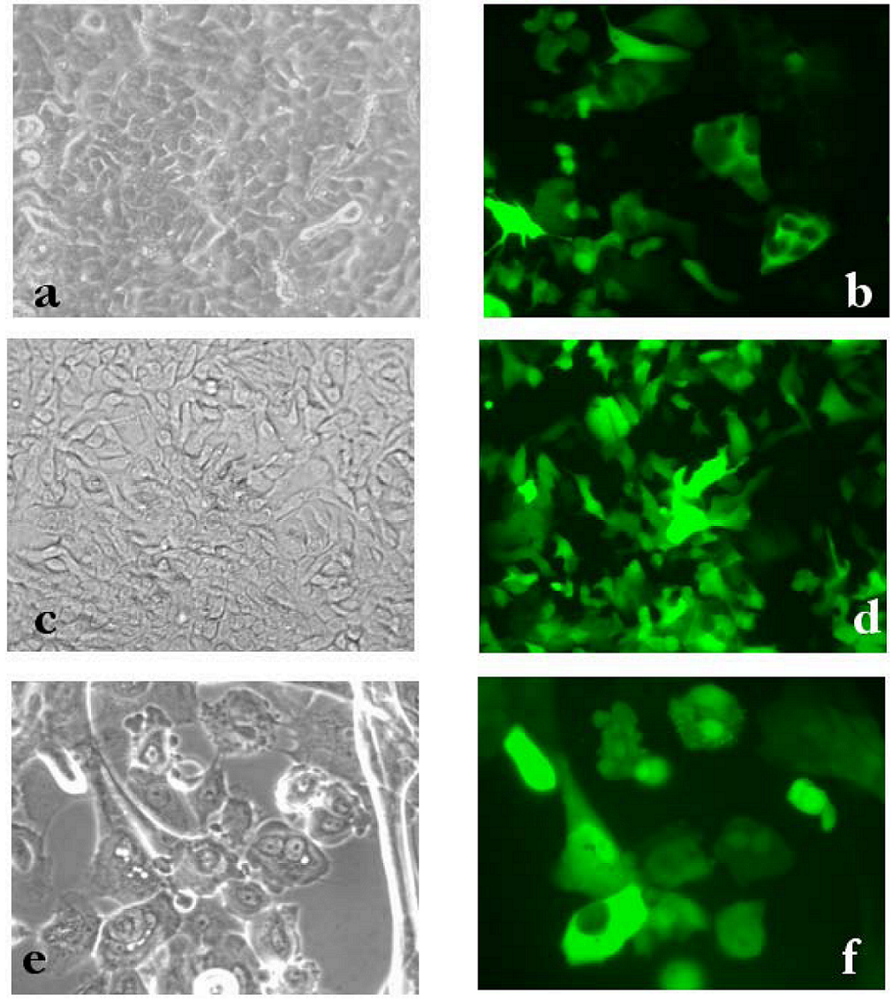
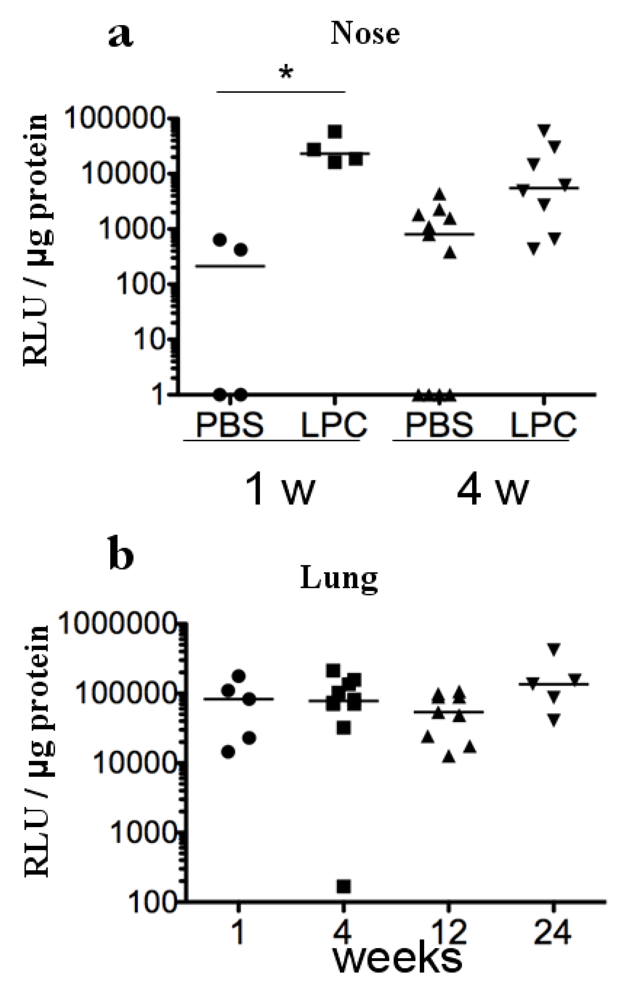
3 .Experimental Section
4. Conclusions
Acknowledgments
References
- Griesenbach, U.; Alton, E.W.F.W. Cystic fibrosis gene therapy: successes, failures and hopes for the future. Exp. Rev. Resp. Med. 2009, 3, 363–371. [Google Scholar] [CrossRef]
- Limberis, M.; Anson, D.S.; Fuller, M.; Parsons, D.W. Recovery of airway cystic fibrosis transmembrane conductance regulator function in mice with cystic fibrosis after single-dose lentivirus-mediated gene transfer. Hum. Gene Ther. 2002, 13, 1961–1970. [Google Scholar] [PubMed]
- Johnson, L.G.; Olsen, J.C.; Naldini, L.; Boucher, R. Pseudotyped human lentiviral vector-mediated gene transfer to airway epithelia in vivo. Gene Ther. 2000, 7, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Goldman, M.J.; Lee, P.S.; Yang, J.S.; Wilson, J.M. Lentiviral vectors for gene therapy of cystic fibrosis. Hum. Gene Ther. 1997, 8, 2261–2268. [Google Scholar] [CrossRef]
- McKay, T.; Patel, M.; Pickles, R.J.; Johnson, L.G.; Olsen, J.C. Influenza M2 envelope protein augments avian influenza hemagglutinin pseudotyping of lentiviral vectors. Gene Ther. 2006, 13, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Slepushkin, V.; Zabner, J.; Keshavjee, S.; Johnston, J.C.; Sauter, S.L.; Jolly, D.J.; Dubensky Jr., T.W.; Davidson, B.L.; McCray Jr., P.B. Feline immunodeficiency virus vectors persistently transduce nondividing airway epithelia and correct the cystic fibrosis defect . J. Clin. Invest. 1999, 104, R55–R62. [Google Scholar] [CrossRef] [PubMed]
- Sersale, G.; Carpani, D.; Casotti, V.; Livraghi, A.; Carrabino, S.; Di Cicco, M.; Assael, B.M.; Giunta, A.; Conese, M. Inhibition of nonviral cationic liposome-mediated gene transfer into primary human respiratory cells by interferon-gamma. J. Mol. Med. 2002, 80, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Sersale, G.; Casotti, V.; Di Cicco, M.; Carpani, D.; Muggia, A.; Calori, G.; Assael, B M.; Conese, M. Human respiratory cells from nasal polyps as a model for gene transfer by non-viral cationic vectors . Acta Otolaryngologica 2001, 121, 76–82. [Google Scholar] [CrossRef]
- Di Gioia, S.; Rejman, J.; Carrabino, S.; De Fino, I.; Rudolph, C.; Doherty, A.; Hyndman, L.; Di Cicco, M.; Copreni, E.; Bragonzi, A.; Colombo, C.; Boyd, A.C.; Conese, M. Role of biophysical parameters on ex vivo and in vivo gene transfer to the airway epithelium by polyethylenimine/albumin complexes. Biomacromolecules 2008, 9, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Castellani, C.; Conese, M. Lentiviral Vectors and Cystic Fibrosis Gene Therapy. Viruses 2010, 2, 395–412. [Google Scholar] [CrossRef]
- Follenzi, A.; Ailles, L.E.; Bakovic, S.; Geuna, M.; Naldini, L. Gene transfer by lentiviral vectors is limited by nuclear translocation and rescued by HIV-1 pol sequences. Nat. Genet. 2000, 25, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Castellani, S.; Di Gioia, S.; Trotta, T.; Maffione, A.B.; Conese, M. Impact of lentiviral vector-mediated transduction on the tightness of a polarized model of airway epithelium and effect of the cationic polymer polyethylenimine. J. Biomed. Biotechnol. 2010, 2010, 1–11. [Google Scholar] [CrossRef]
- Copreni, E.; Penzo, M.; Carrabino, S.; Conese, M. Lentivirus-mediated gene transfer to the respiratory epithelium: a promising approach to gene therapy of cystic fibrosis . Gene Ther. 2004, 11 (Suppl. 1), S67–S75. [Google Scholar] [CrossRef] [PubMed]
- Sinn, P.L.; Arias, A.C.; Brogden, K.A.; McCray Jr., P.B. Lentivirus vector can be readministered to nasal epithelia without blocking immune responses . J. Virol. 2008, 82, 10684–10692. [Google Scholar] [CrossRef] [PubMed]
- Sinn, PL.; Burnight, E.R.; Hickey, M.A.; Blissard, G.W.; McCray Jr., P.B. Persistent gene expression in mouse nasal epithelia following feline immunodeficiency virus-based vector gene transfer . J. Virol. 2005, 79, 12818–12827. [Google Scholar] [CrossRef] [PubMed]
- Kremer, K.L.; Dunning, K.R.; Parsons, D.W.; Anson, D.S. Gene delivery to airway epithelial cells in vivo: a direct comparison of apical and basolateral transduction strategies using pseudotyped lentivirus vectors. J. Gene Med. 2007, 9, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Stocker, A.G.; Kremer, K.L.; Koldej, R.; Miller, D.S.; Anson, D.S.; Parsons, D.W. Single-dose lentiviral gene transfer for lifetime airway gene expression. J. Gene Med. 2009, 11, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Rejman, J.; Di Gioia, S.; Bragonzi, A.; Conese, M. Pseudomonas aeruginosa infection destroys the barrier function of lung epithelium and enhances polyplex-mediated transfection. Hum. Gene Ther. 2007, 18, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Limberis, M.P.; Bell, C.L.; Heath, J.; Wilson, J.M. Activation of transgene-specific T cells following lentivirus-mediated gene delivery to mouse lung. Mol. Ther. 2010, 18, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Buckley, S.M.; Howe, S.J.; Sheard, V.; Ward, N.J.; Coutelle, C.; Thrasher, A.J.; Waddington, S.N.; McKay, T.R. Lentiviral transduction of the murine lung provides efficient pseudotype and developmental stage-dependent cell-specific transgene expression. Gene Ther. 2008, 15, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Tarantal, A.F.; Lee, C.I.; Ekert, J.E.; McDonald, R.; Kohn, D.B.; Plopper, C.G.; Case, S.S.; Bunnell, B.A. Lentiviral vector gene transfer into fetal rhesus monkeys (Macaca mulatta): lung-targeting approaches. Mol. Ther. 2001, 4, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.Y.; McKay, K.; van Asperen, P.; Zheng, M.; Fleming, J.; Ginn, S.L.; Kizana, E.; Latham, M.; Feneley, M.P.; Kirkland, P.D.; Rowe, P.B.; Lumbers, E.R.; Alexander, I.E. Lentivirus vector-mediated gene transfer to the developing bronchiolar airway epithelium in the fetal lamb. J. Gene Med. 2007, 9, 429–439. [Google Scholar] [CrossRef]
- Parsons, D.; Martella, T.; Hopkins, P.; Limberis, M.; Martin, J. Lysophosphatidylcholine (LPC) provides greater enhancement of in-vivo adenoviral airway gene transfer than polidocanol . Respirology 1999, 4 (Suppl. 1), A11. [Google Scholar] [PubMed]
- Borok, Z.; Harboe-Schmidt, J.E.; Brody, S.L.; You, Y.; Zhou, B.; Li, X.; Cannon, P.M.; Kim, K. J.; Crandall, E.D.; Kasahara, N. Vesicular stomatitis virus G-pseudotyped lentivirus vectors mediate efficient apical transduction of polarized quiescent primary alveolar epithelial cells. J. Virol. 2001, 75, 11747–11754. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, D.W.; Shahbazian, M.; Krantz, Q.T.; Dorin, J.R.; Randell, S.H. Evidence for stem-cell niches in the tracheal epithelium. Am. J. Respir. Cell Mol. Biol. 2001, 24, 662–670. [Google Scholar] [PubMed]
- Rawlins, E.L.; Hogan, B.L. Ciliated epithelial cell lifespan in the mouse trachea and lung . Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L231–L234. [Google Scholar] [CrossRef] [PubMed]
- Copreni, E.; Nicolis, E.; Tamanini, A.; Bezzerri, V.; Castellani, S.; Palmieri, L.; Giri, M.G.; Vella, A.; Colombatti, M.; Rizzotti, P.; Conese, M.; Cabrini, G. Late generation lentiviral vectors: evaluation of inflammatory potential in human airway epithelial cells. Virus Res. 2009, 144, 8–17. [Google Scholar] [CrossRef]
- Copreni, E.; Conese, M. Institute for the Experimental Treatment of Cystic Fibrosis, 20132 Milan, Italy . 2010, Unpublished work. [Google Scholar]
- Copreni, E.; Castellani, S.; Palmieri, L.; Penzo, M.; Conese, M. Involvement of glycosaminoglycans in vesicular stomatitis virus G glycoprotein pseudotyped lentiviral vector-mediated gene transfer into airway epithelial cells. J. Gene Med. 2008, 10, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Bragonzi, A.; Dina, G.; Villa, A.; Calori, G.; Biffi, A.; Bordignon, C.; Assael, B.M.; Conese, M. Biodistribution and transgene expression with nonviral cationic vector/DNA complexes in the lungs. Gene Ther. 2000, 7, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Copreni, E.; Palmieri, L.; Castellani, S.; Conese, M. A VSV-G Pseudotyped Last Generation Lentiviral Vector Mediates High Level and Persistent Gene Transfer in Models of Airway Epithelium In Vitro and In Vivo. Viruses 2010, 2, 1577-1588. https://doi.org/10.3390/v2081577
Copreni E, Palmieri L, Castellani S, Conese M. A VSV-G Pseudotyped Last Generation Lentiviral Vector Mediates High Level and Persistent Gene Transfer in Models of Airway Epithelium In Vitro and In Vivo. Viruses. 2010; 2(8):1577-1588. https://doi.org/10.3390/v2081577
Chicago/Turabian StyleCopreni, Elena, Lucia Palmieri, Stefano Castellani, and Massimo Conese. 2010. "A VSV-G Pseudotyped Last Generation Lentiviral Vector Mediates High Level and Persistent Gene Transfer in Models of Airway Epithelium In Vitro and In Vivo" Viruses 2, no. 8: 1577-1588. https://doi.org/10.3390/v2081577
APA StyleCopreni, E., Palmieri, L., Castellani, S., & Conese, M. (2010). A VSV-G Pseudotyped Last Generation Lentiviral Vector Mediates High Level and Persistent Gene Transfer in Models of Airway Epithelium In Vitro and In Vivo. Viruses, 2(8), 1577-1588. https://doi.org/10.3390/v2081577