

eEF-2K Deficiency Boosts the Virus-Specific Effector CD8+ T Cell Responses During Viral Infection

 , and
, and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Virus Preparation and Titration
2.3. Viral Infection
2.4. Tissue Processing
2.5. Flow Cytometry and Tetramer Staining
2.6. Sorting of VACV-Specific CD8⁺ T Cells
2.7. RNA Sequencing and Data Analysis
2.8. Intracellular Cytokine and Protein Staining
2.9. RNA Extraction and Quantitative PCR (qPCR)
- Traf3 Forward: GTGAACCTGCTGAAGGAGTGGA
- Traf3 Reverse: TTCGGAGCATCTCCTTCTGCCT
- Gapdh Forward: GTTGTCTCCTGCGACTTCA
- Gapdh Reverse: GGTGGTCCAGGGTTTCTTA
2.10. Illustration and Figures
2.11. Statistical Analysis
3. Results
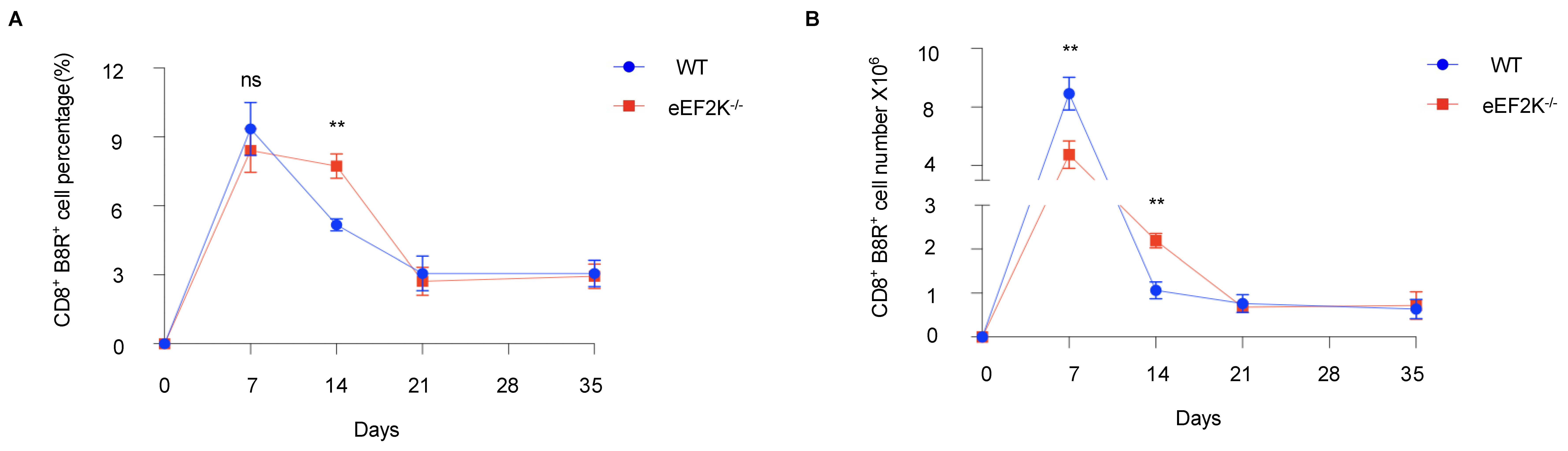
3.1. eEF-2K⁻/⁻ Mice Show Enhanced Response to VACV Infection During the Effector Stage
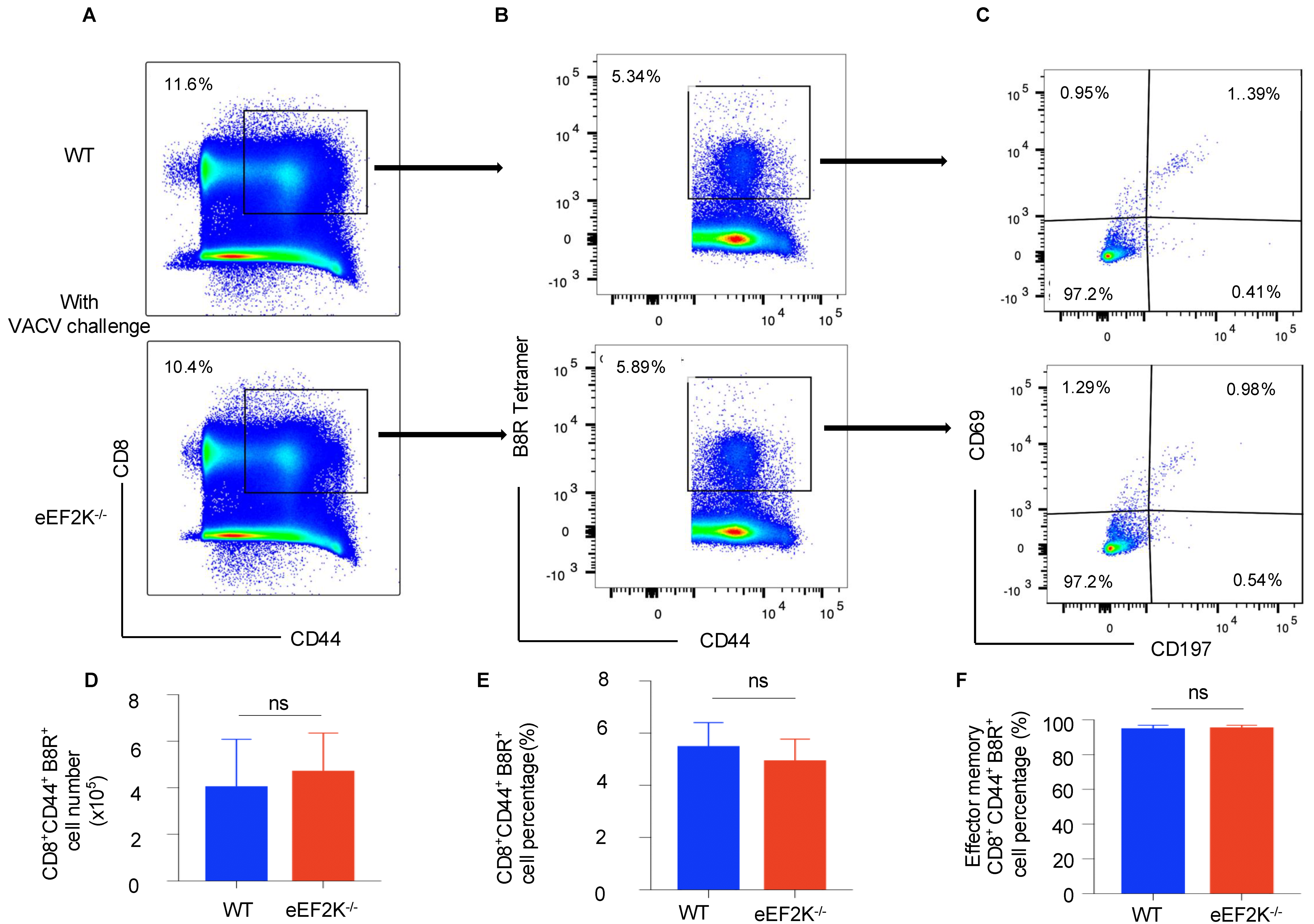
3.2. eEF-2K Deficiency Does Not Influence VACV-Specific Memory T Cell Formation
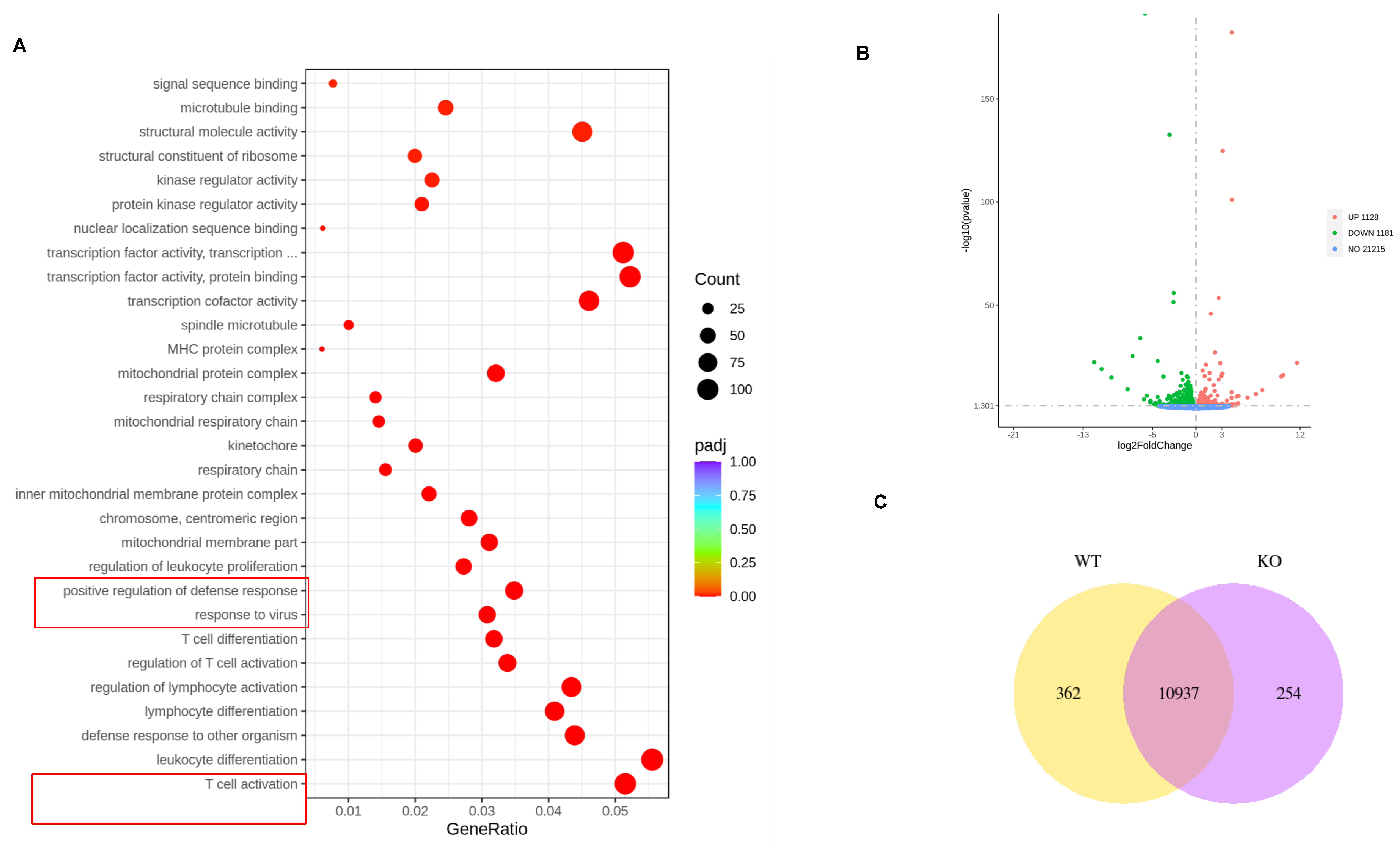
3.3. Involvement of Transcriptional Alteration in the Augmented Response of VACV-Specific eEF-2K⁻/⁻ Effector CD8⁺ T Cells to Viral Infection
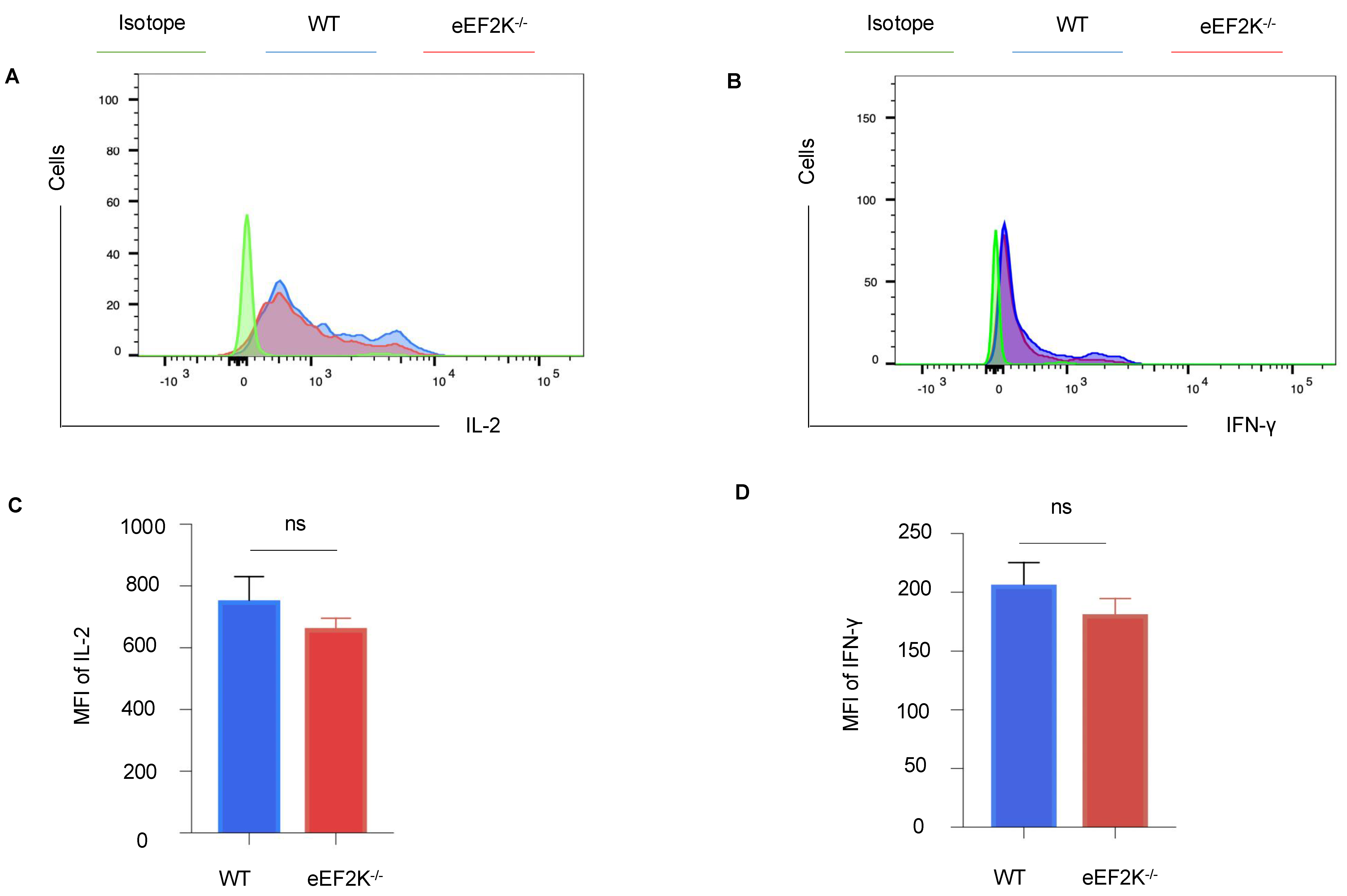
3.4. Functional Competence of VACV-Specific eEF-2K⁻/⁻ Effector CD8⁺ T Cells
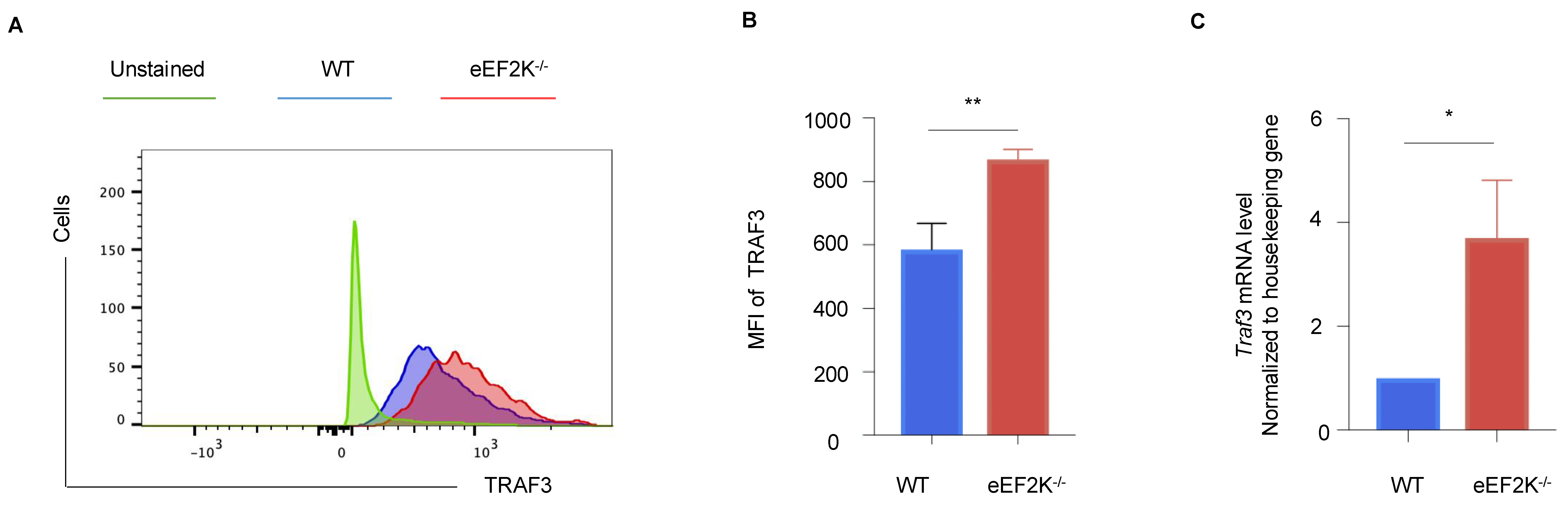
3.5. TRAF3 Mediates the Enhanced Antiviral Response in eEF-2K⁻/⁻ Effector CD8⁺ T Cells
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teque, F.; Wegehaupt, A.; Roufs, E.; Killian, M.S. CD8+ Lymphocytes from Healthy Blood Donors Secrete Antiviral Levels of Interferon-Alpha. Viruses 2023, 15, 894. [Google Scholar] [CrossRef]
- Wang, L.; Peng, H.Y.; Pham, A.; Villazana, E.; Ballard, D.J.; Das, J.K.; Kumar, A.; Xiong, X.; Song, J. T Cell Response to SARS-CoV-2 Coinfection and Comorbidities. Pathogens 2023, 12, 321. [Google Scholar] [CrossRef]
- Kaech, S.M.; Wherry, E.J.; Ahmed, R. Effector and memory T-cell differentiation: Implications for vaccine development. Nat. Rev. Immunol. 2002, 2, 251–262. [Google Scholar] [CrossRef]
- Meineke, R.; Rimmelzwaan, G.F.; Elbahesh, H. Influenza Virus Infections and Cellular Kinases. Viruses 2019, 11, 171. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Kumar, A.; Das, J.K.; Ren, Y.; Peng, H.Y.; Ballard, D.J.; Xiong, X.; Davis, J.R.; Ren, X.; Yang, J.M.; et al. Expression of NAC1 Restrains the Memory Formation of CD8(+) T Cells during Viral Infection. Viruses 2022, 14, 1713. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Peng, H.Y.; Kishore Das, J.; Kumar, A.; Ren, Y.; Ballard, D.J.; Xiong, X.; Yang, W.; Ren, X.; de Figueiredo, P.; et al. NAC1 confines virus-specific memory formation of CD4(+) T cells through the ROCK1-mediated pathway. J. Med. Virol. 2023, 95, e28957. [Google Scholar] [CrossRef] [PubMed]
- Salek-Ardakani, S.; Song, J.; Halteman, B.S.; Jember, A.G.; Akiba, H.; Yagita, H.; Croft, M. OX40 (CD134) controls memory T helper 2 cells that drive lung inflammation. J. Exp. Med. 2003, 198, 315–324. [Google Scholar] [CrossRef]
- Ryazanov, A.G.; Ward, M.D.; Mendola, C.E.; Pavur, K.S.; Dorovkov, M.V.; Wiedmann, M.; Erdjument-Bromage, H.; Tempst, P.; Parmer, T.G.; Prostko, C.R.; et al. Identification of a new class of protein kinases represented by eukaryotic elongation factor-2 kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 4884–4889. [Google Scholar] [CrossRef]
- Wang, H.; Jin, W.; Li, Z.; Guo, C.; Zhang, L.; Fu, L. Targeting eukaryotic elongation factor 2 kinase (eEF2K) with small-molecule inhibitors for cancer therapy. Drug Discov. Today 2024, 29, 104155. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Zeng, F.; He, Y.; Meng, Y.; Sun, H.; Su, J.; Zhao, S.; Cheng, Y.; Chen, X.; Yin, M. EEF2K silencing inhibits tumour progression through repressing SPP1 and synergises with BET inhibitors in melanoma. Clin. Transl. Med. 2022, 12, e722. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Ben-David, Y.; Rotin, D.; Zacksenhaus, E. Inhibition of eEF2K synergizes with glutaminase inhibitors or 4EBP1 depletion to suppress growth of triple-negative breast cancer cells. Sci. Rep. 2021, 11, 9181. [Google Scholar] [CrossRef]
- Wu, Y.; Xie, J.; Jin, X.; Lenchine, R.V.; Wang, X.; Fang, D.M.; Nassar, Z.D.; Butler, L.M.; Li, J.; Proud, C.G. eEF2K enhances expression of PD-L1 by promoting the translation of its mRNA. Biochem. J. 2020, 477, 4367–4381. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Song, H.; Chen, G.; Yang, X.; Liu, J.; Ge, Y.; Lu, J.; Qin, Q.; Zhang, C.; Xu, L.; et al. eEF2K promotes progression and radioresistance of esophageal squamous cell carcinoma. Radiother. Oncol. 2017, 124, 439–447. [Google Scholar] [CrossRef] [PubMed]
- De Gassart, A.; Demaria, O.; Panes, R.; Zaffalon, L.; Ryazanov, A.G.; Gilliet, M.; Martinon, F. Pharmacological eEF2K activation promotes cell death and inhibits cancer progression. EMBO Rep. 2016, 17, 1471–1484. [Google Scholar] [CrossRef]
- Cheng, Y.; Ren, X.; Zhang, Y.; Shan, Y.; Huber-Keener, K.J.; Zhang, L.; Kimball, S.R.; Harvey, H.; Jefferson, L.S.; Yang, J.M. Integrated regulation of autophagy and apoptosis by EEF2K controls cellular fate and modulates the efficacy of curcumin and velcade against tumor cells. Autophagy 2013, 9, 208–219. [Google Scholar] [CrossRef]
- Ma, T. Roles of eukaryotic elongation factor 2 kinase (eEF2K) in neuronal plasticity, cognition, and Alzheimer disease. J. Neurochem. 2023, 166, 47–57. [Google Scholar] [CrossRef]
- Smith, P.R.; Loerch, S.; Kunder, N.; Stanowick, A.D.; Lou, T.F.; Campbell, Z.T. Functionally distinct roles for eEF2K in the control of ribosome availability and p-body abundance. Nat. Commun. 2021, 12, 6789. [Google Scholar] [CrossRef] [PubMed]
- Bianco, C.; Thompson, L.; Mohr, I. Repression of eEF2K transcription by NF-kappaB tunes translation elongation to inflammation and dsDNA-sensing. Proc. Natl. Acad. Sci. USA 2019, 116, 22583–22590. [Google Scholar] [CrossRef] [PubMed]
- Versele, M.M.C.; Proud, C.G.; Rockx, C.; IWeyer, K.; Baelen, K.; Blencke, S.; Wanndinge, S.; Diels, G.; Berthelot, D.; Viellevoye, M.; et al. Managing stress: Discovery of inhibitors of the atypical kinase eEF2K and the class III PI3K, VPS34. In AACR Annual Meeting 2014; American Association for Cancer Research: San Diego, CA, USA, 2014. [Google Scholar]
- Chu, H.P.; Liao, Y.; Novak, J.S.; Hu, Z.; Merkin, J.J.; Shymkiv, Y.; Braeckman, B.P.; Dorovkov, M.V.; Nguyen, A.; Clifford, P.M.; et al. Germline quality control: eEF2K stands guard to eliminate defective oocytes. Dev. Cell 2014, 28, 561–572. [Google Scholar] [CrossRef]
- Das, J.K.; Ren, Y.; Kumar, A.; Peng, H.Y.; Wang, L.; Xiong, X.; Alaniz, R.C.; de Figueiredo, P.; Ren, X.; Liu, X.; et al. Elongation factor-2 kinase is a critical determinant of the fate and antitumor immunity of CD8(+) T cells. Sci. Adv. 2022, 8, eabl9783. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, F.; Tian, Y.; Zeng, G.; Lei, G.; Bai, Z.; Wang, Y.; Ge, X.; Wang, J.; Xiao, C.; et al. Enhanced NK cell activation via eEF2K-mediated potentiation of the cGAS-STING pathway in hepatocellular carcinoma. Int. Immunopharmacol. 2024, 129, 111628. [Google Scholar] [CrossRef]
- Peng, H.Y.; Wang, L.; Das, J.K.; Kumar, A.; Ballard, D.J.; Ren, Y.; Xiong, X.; de Figueiredo, P.; Yang, J.M.; Song, J. Control of CD4(+) T cells to restrain inflammatory diseases via eukaryotic elongation factor 2 kinase. Signal Transduct. Target. Ther. 2023, 8, 415. [Google Scholar] [CrossRef] [PubMed]
- Cotter, C.A.; Earl, P.L.; Wyatt, L.S.; Moss, B. Preparation of Cell Cultures and Vaccinia Virus Stocks. Curr. Protoc. Mol. Biol. 2017, 117, 16 16 1–16 16 18. [Google Scholar] [CrossRef]
- Haque, M.; Song, J.; Fino, K.; Wang, Y.; Sandhu, P.; Song, X.; Norbury, C.; Ni, B.; Fang, D.; Salek-Ardakani, S.; et al. C-Myc regulation by costimulatory signals modulates the generation of CD8+ memory T cells during viral infection. Open Biol. 2016, 6, 150208. [Google Scholar] [CrossRef]
- Hornick, E.L.; Wallis, A.M.; Bishop, G.A. TRAF3 enhances type I interferon receptor signaling in T cells by modulating the phosphatase PTPN22. Sci. Signal 2022, 15, eabn5507. [Google Scholar] [CrossRef]
- Chen, F.; Chen, L.; Li, Y.; Sang, H.; Zhang, C.; Yuan, S.; Yang, J. TRAF3 Positively Regulates Host Innate Immune Resistance to Influenza A Virus Infection. Front. Cell Infect. Microbiol. 2022, 12, 839625. [Google Scholar] [CrossRef] [PubMed]
- Hacker, H.; Tseng, P.H.; Karin, M. Expanding TRAF function: TRAF3 as a tri-faced immune regulator. Nat. Rev. Immunol. 2011, 11, 457–468. [Google Scholar] [CrossRef]
- Zhao, X.; Zhong, C.; Zhu, R.; Gong, R.; Liu, B.; He, L.; Tian, S.; Jin, J.; Jiang, T.; Chen, J.L.; et al. Structure-Activity Relationship Studies of Substituted 2-Phenyl-1,2,4-triazine-3,5(2H,4H)-dione Analogues: Development of Potent eEF2K Degraders against Triple-Negative Breast Cancer. J. Med. Chem. 2024, 67, 15837–15861. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Zhu, R.; Jiang, T.; Tian, S.; Zhao, X.; Wan, X.; Jiang, S.; Chen, Z.; Gong, R.; He, L.; et al. Design and Characterization of a Novel eEF2K Degrader with Potent Therapeutic Efficacy Against Triple-Negative Breast Cancer. Adv. Sci. 2024, 11, e2305035. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Color | Company & Cat NO. |
|---|---|---|
| IL-2 | BV421 | BioLegend 503825 |
| IFN-y | FITC | BioLegend 505806 |
| CD8 | PE | BioLegend 100708 |
| CD44 | FITC | BioLegend 103005 |
| CD69 | BV711 | BioLegend 104537 |
| CD197 | BV421 | BioLegend 120119 |
| BCL-2 | PE | Miltenyi Biotec 130-118-688 |
| VACV tetramer | APC | NIH Tetramer core B8R |
| TRAF3 | Alex Fluor 488 | Proteintech CL488-66310 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Song, B.S.; Poojary, R.; Xiong, X.; Ren, X.; Yang, J.-M.; Song, J. eEF-2K Deficiency Boosts the Virus-Specific Effector CD8+ T Cell Responses During Viral Infection. Viruses 2025, 17, 26. https://doi.org/10.3390/v17010026
Wang L, Song BS, Poojary R, Xiong X, Ren X, Yang J-M, Song J. eEF-2K Deficiency Boosts the Virus-Specific Effector CD8+ T Cell Responses During Viral Infection. Viruses. 2025; 17(1):26. https://doi.org/10.3390/v17010026
Chicago/Turabian StyleWang, Liqing, Benny Shone Song, Rayansh Poojary, Xiaofang Xiong, Xingcong Ren, Jin-Ming Yang, and Jianxun Song. 2025. "eEF-2K Deficiency Boosts the Virus-Specific Effector CD8+ T Cell Responses During Viral Infection" Viruses 17, no. 1: 26. https://doi.org/10.3390/v17010026
APA StyleWang, L., Song, B. S., Poojary, R., Xiong, X., Ren, X., Yang, J.-M., & Song, J. (2025). eEF-2K Deficiency Boosts the Virus-Specific Effector CD8+ T Cell Responses During Viral Infection. Viruses, 17(1), 26. https://doi.org/10.3390/v17010026