

Establishment of a Dual-Antigen Indirect ELISA Based on p30 and pB602L to Detect Antibodies against African Swine Fever Virus

 ,
,
Abstract
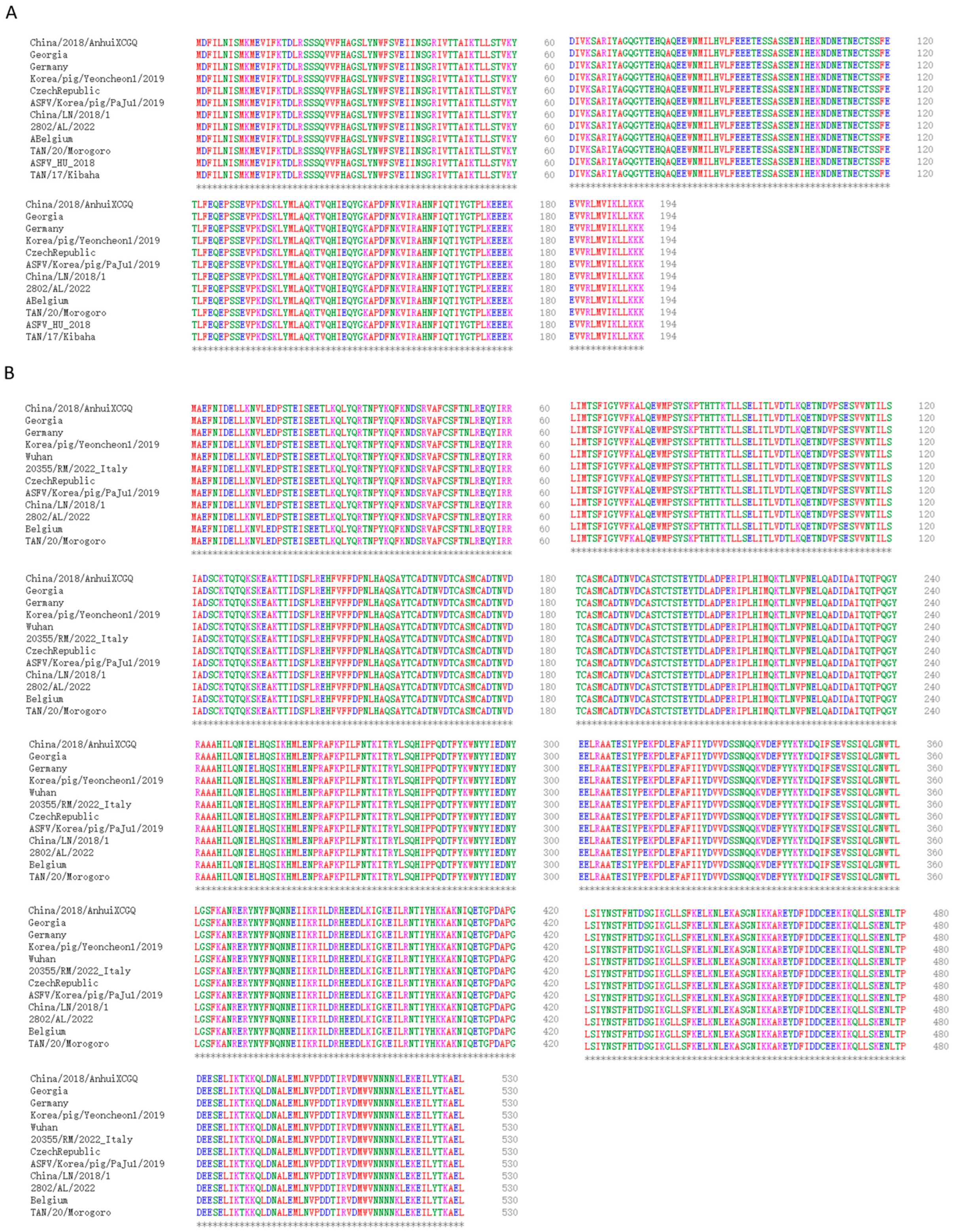
:1. Introduction
2. Materials and Methods
2.1. Serum Samples
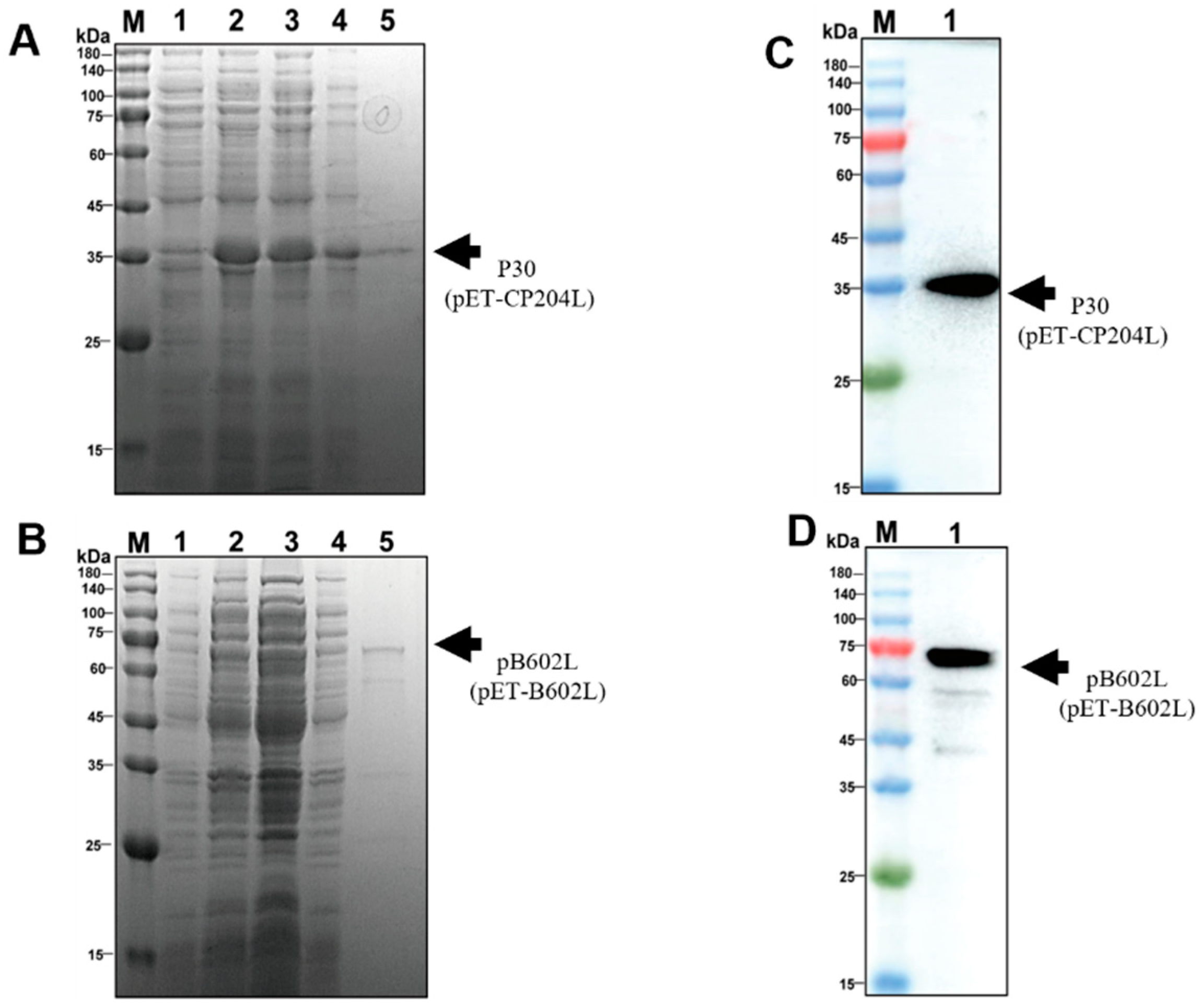
2.2. Expression and Purification of p30 and pB602L
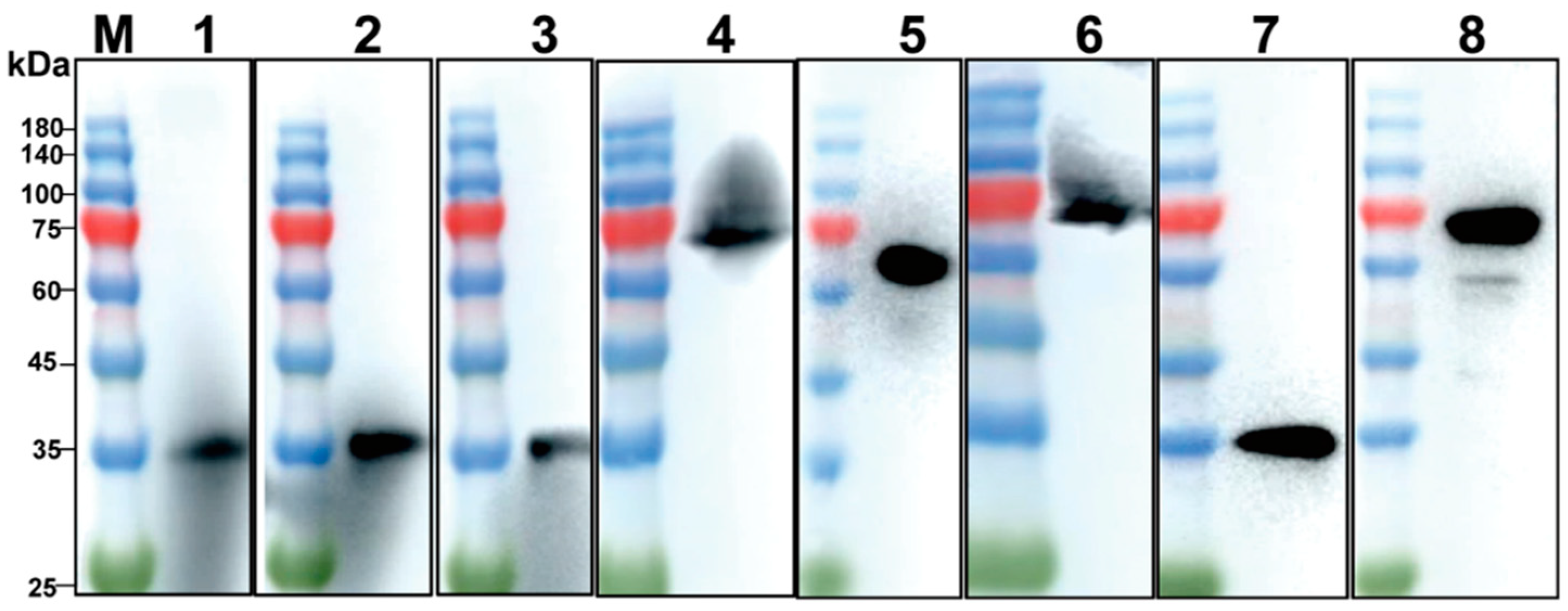
2.3. Western Blotting
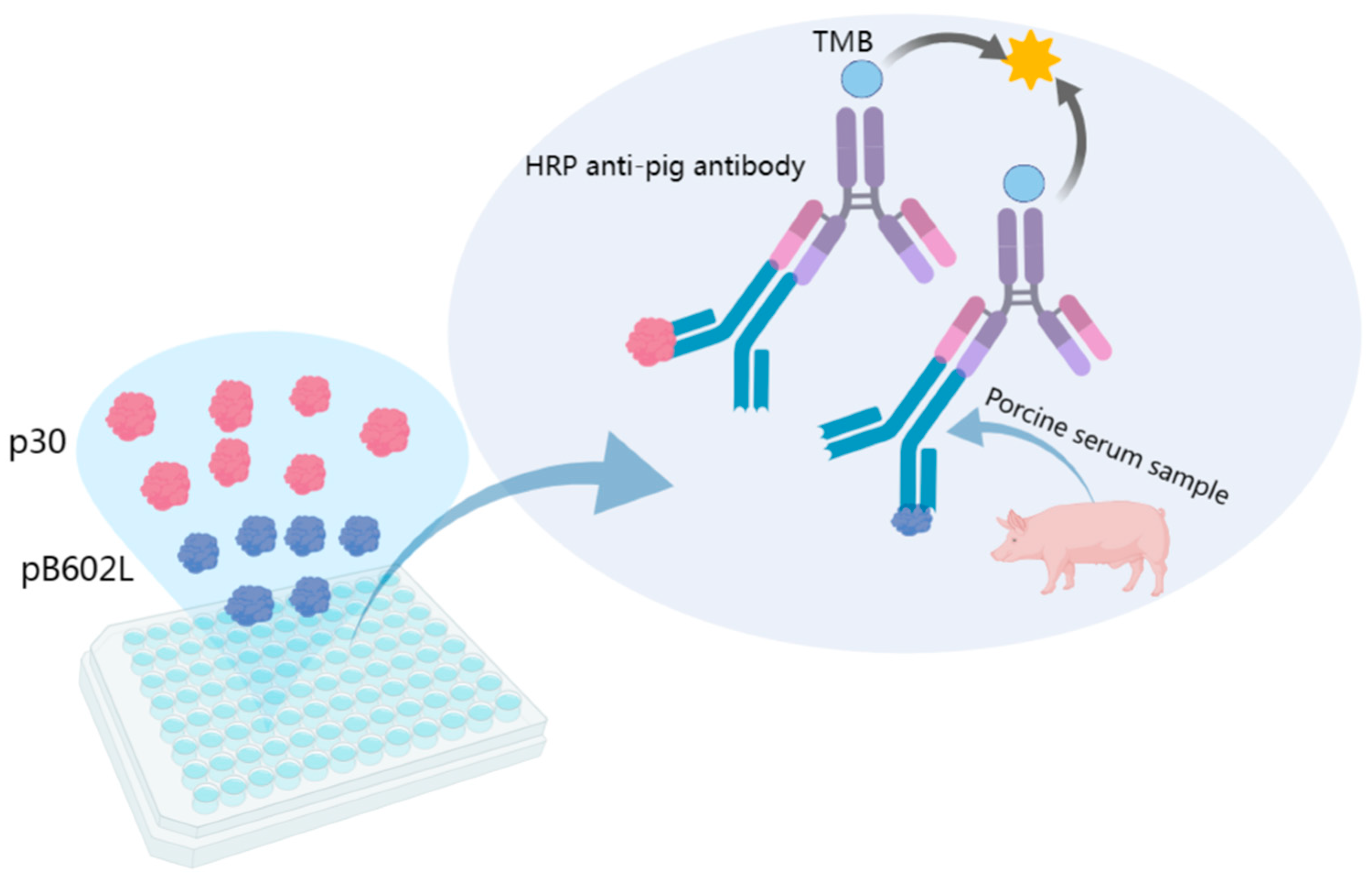
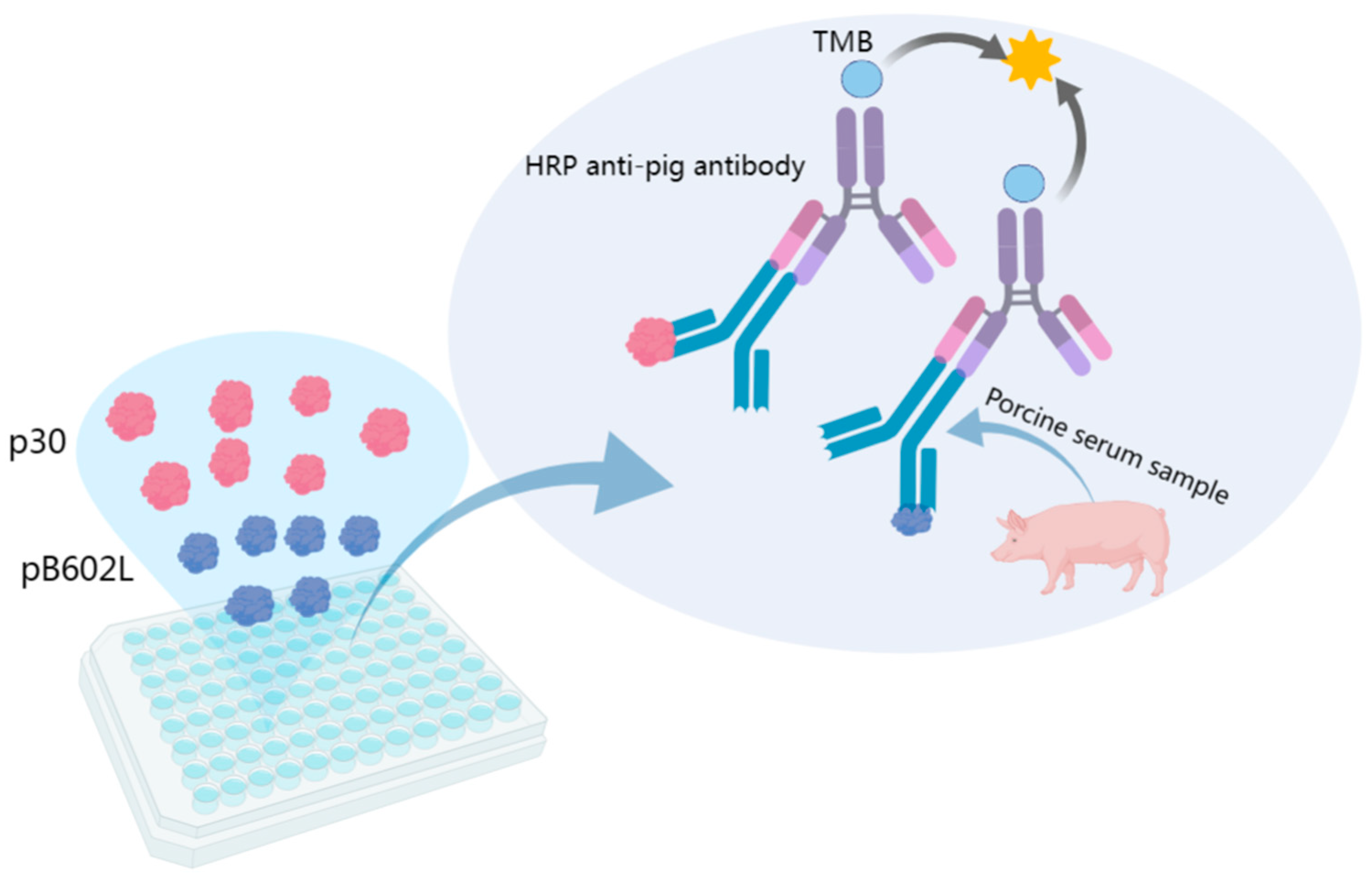
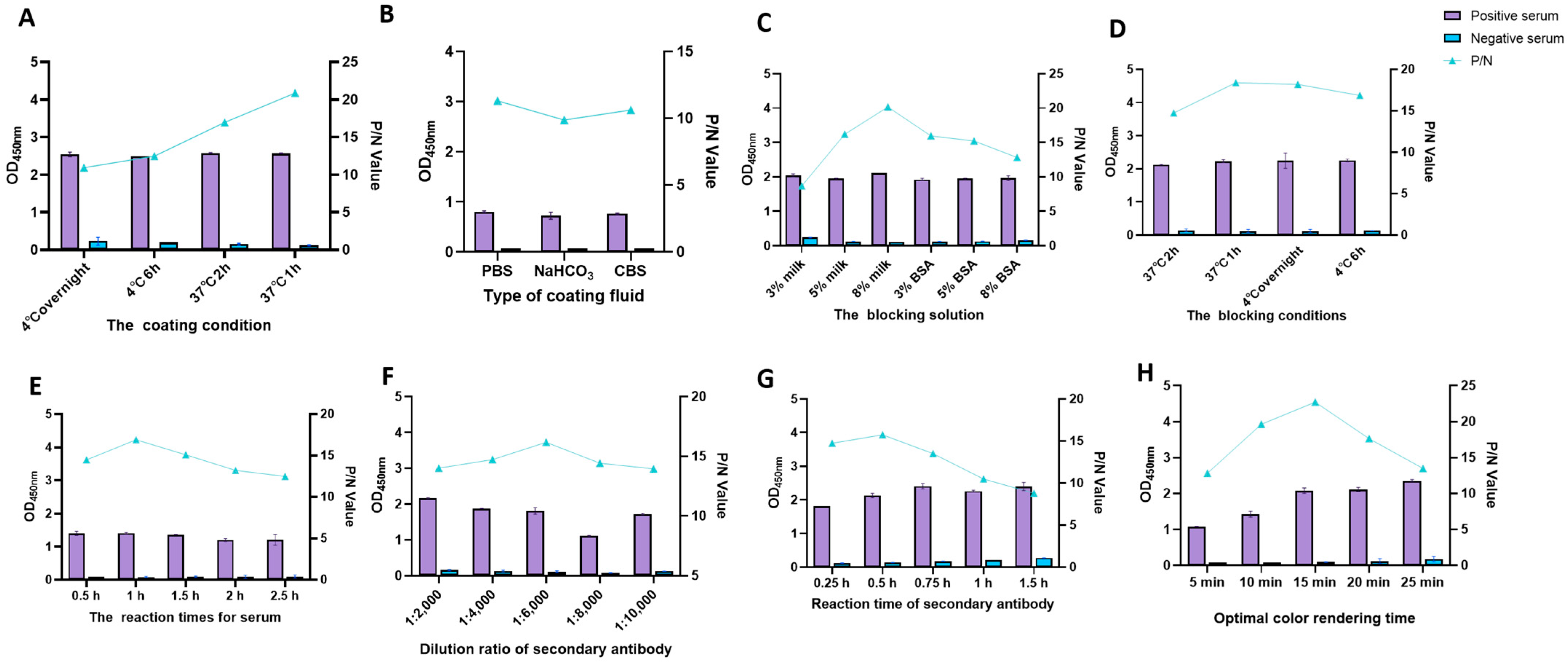
2.4. Establishment of Indirect ELISA Method Based on p30 and pB602L Dual-Antigen
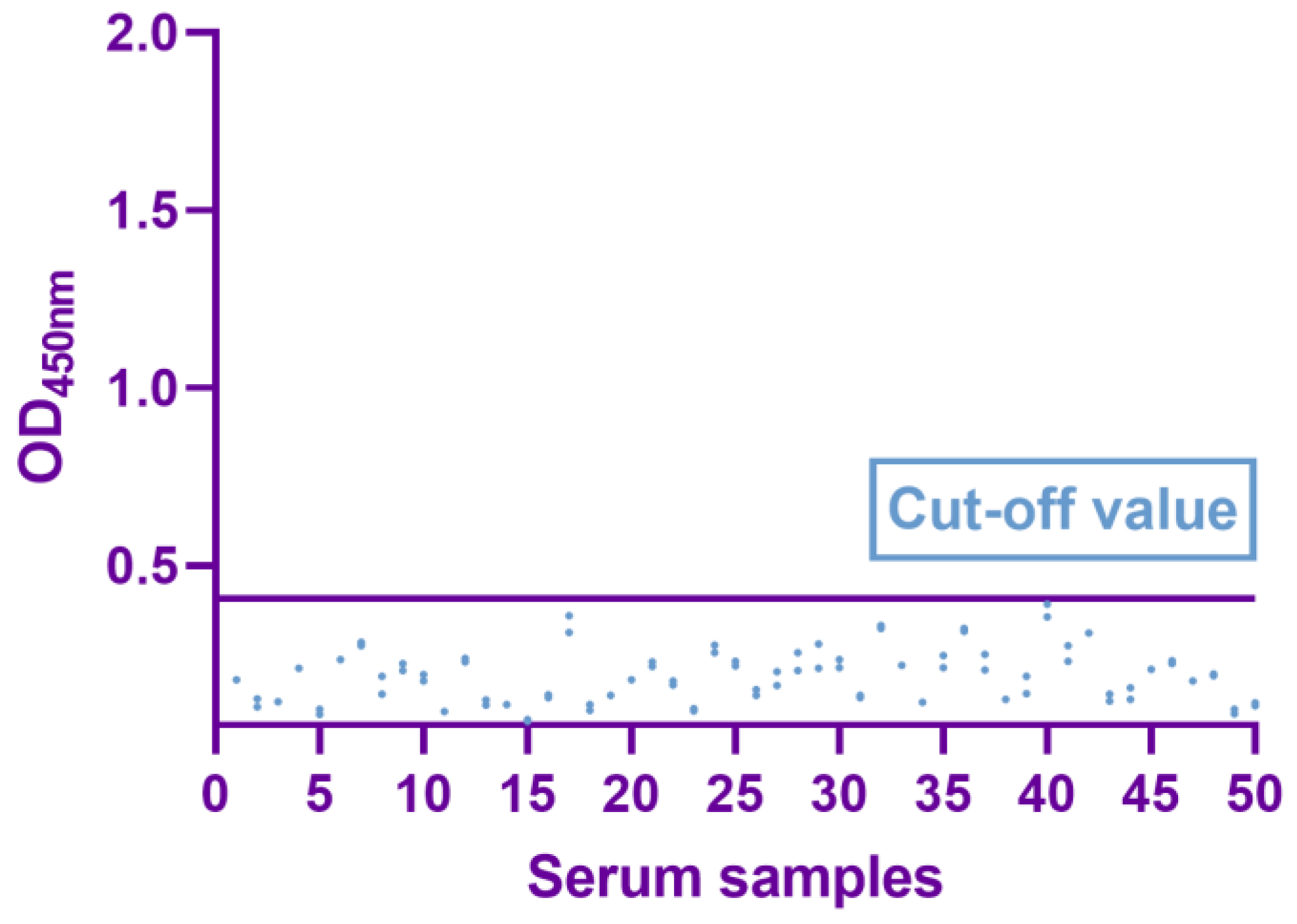
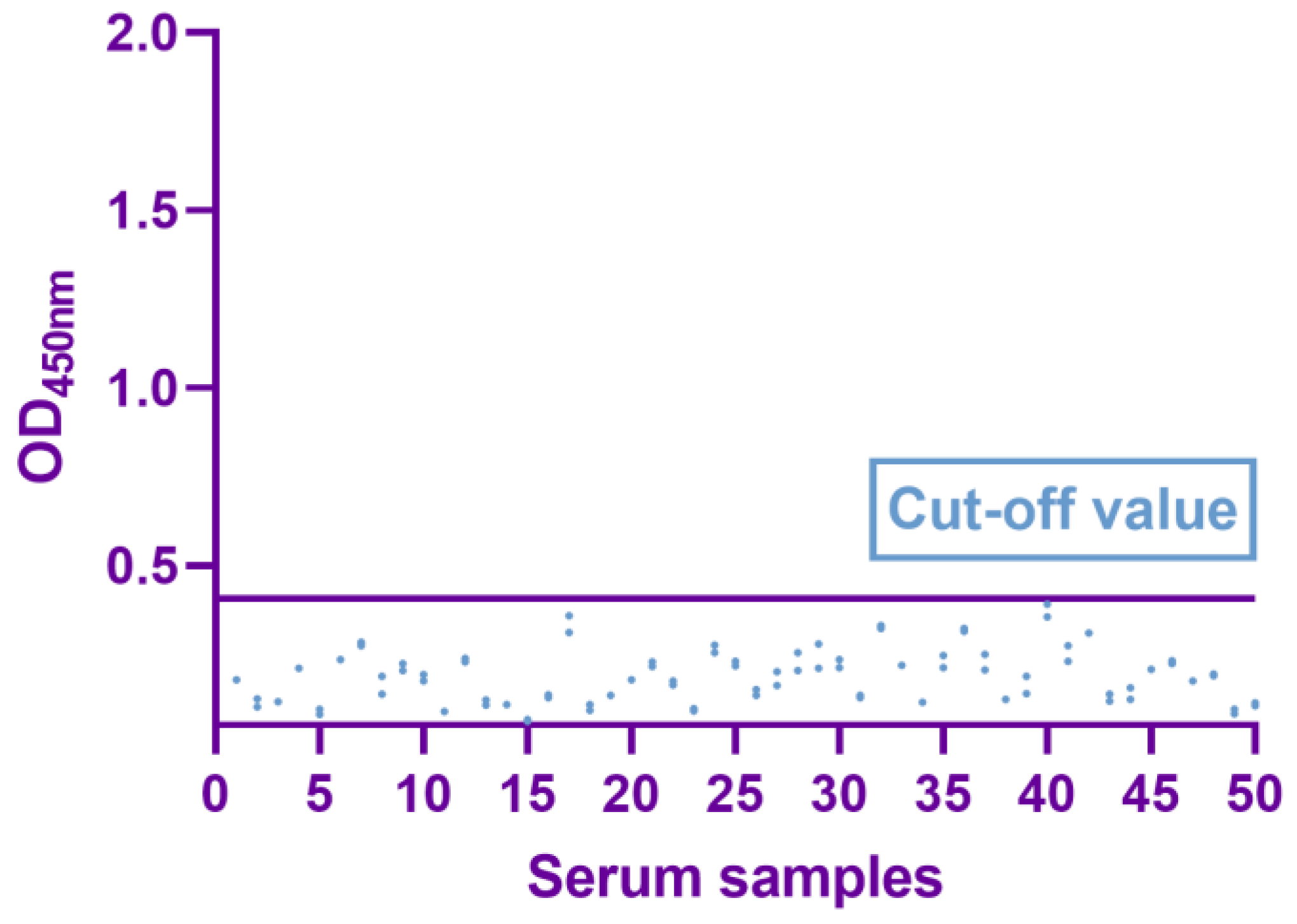
2.5. Determination of the Cutoff Value
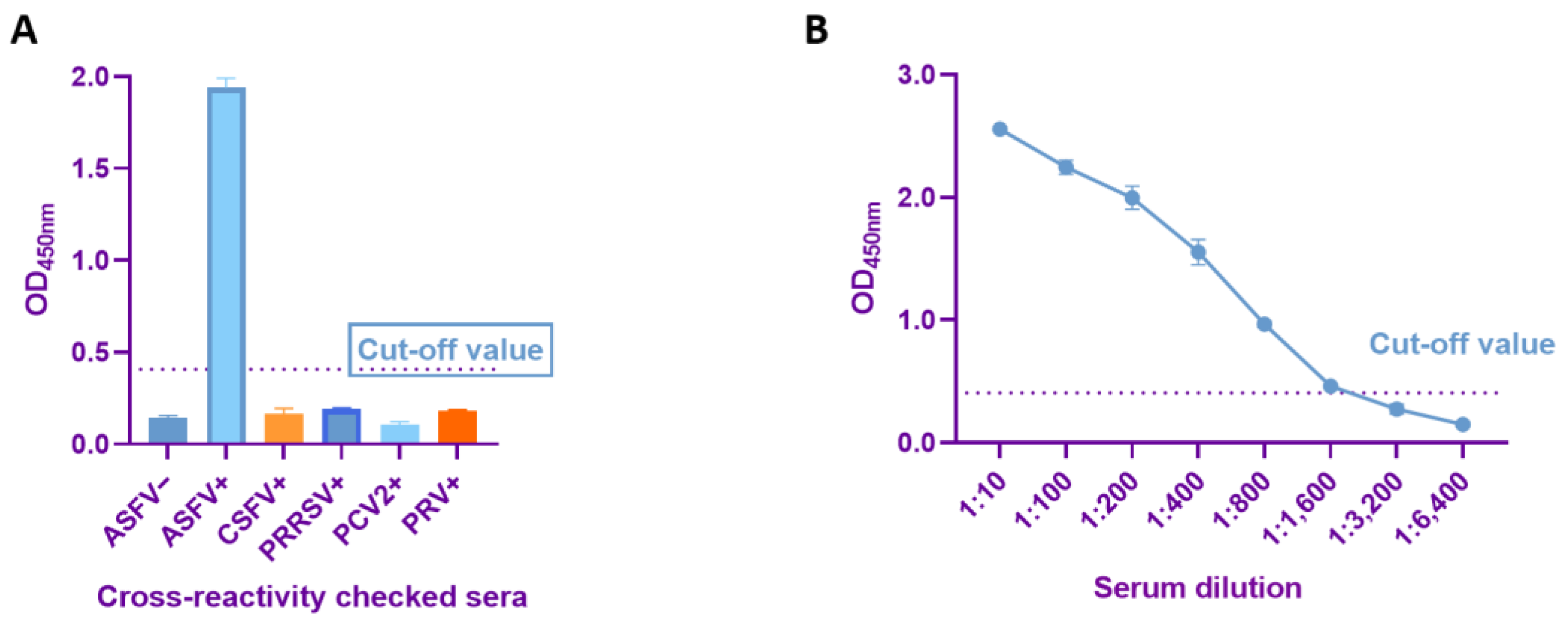
2.6. Sensitivity and Specificity Determination
2.7. Reproducibility of Indirect ELISA with Dual-Antigen
2.8. Comparison of Dual-Antigen Indirect ELISA with Commercial Kits
2.9. Statistical Analysis
3. Results
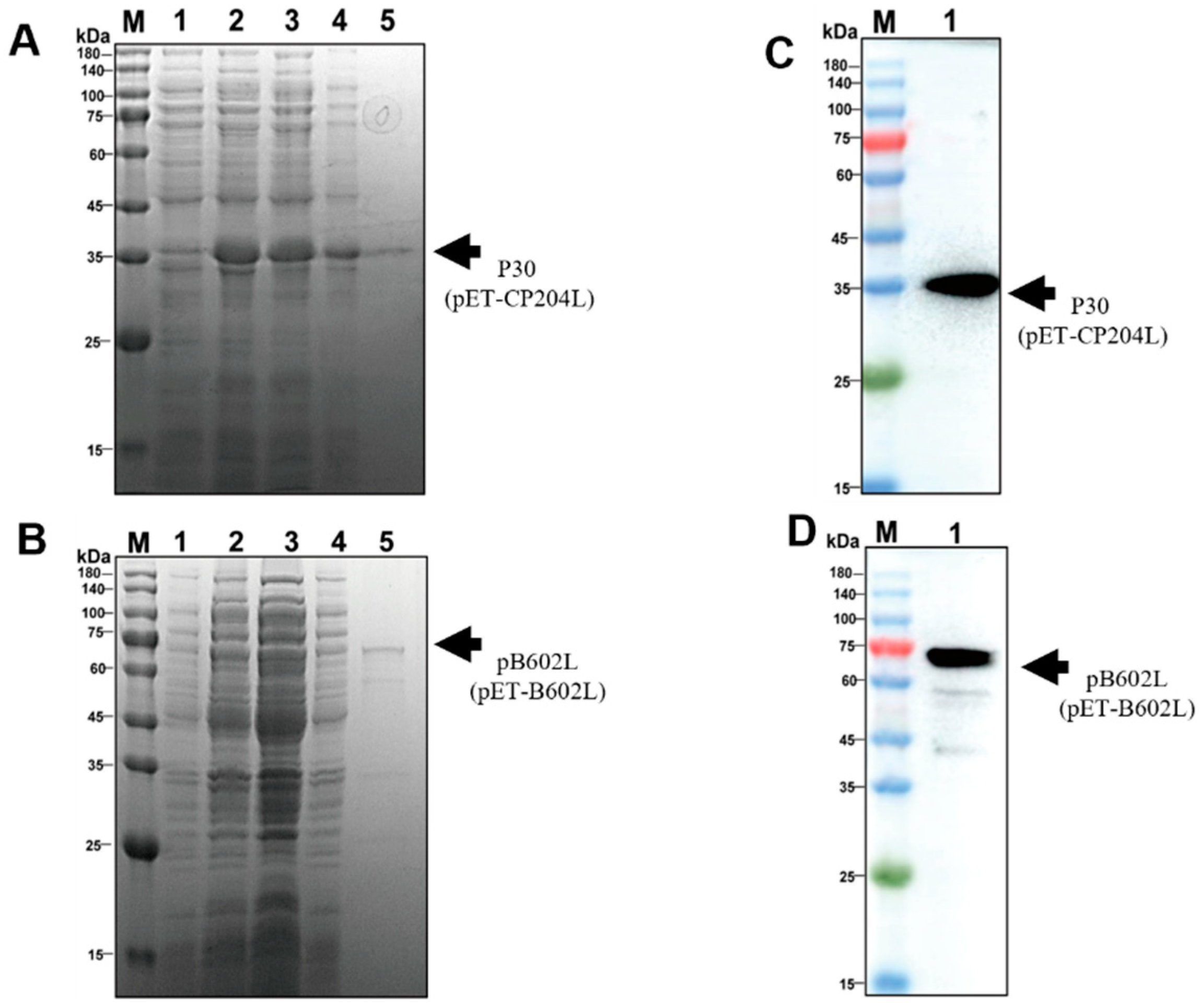
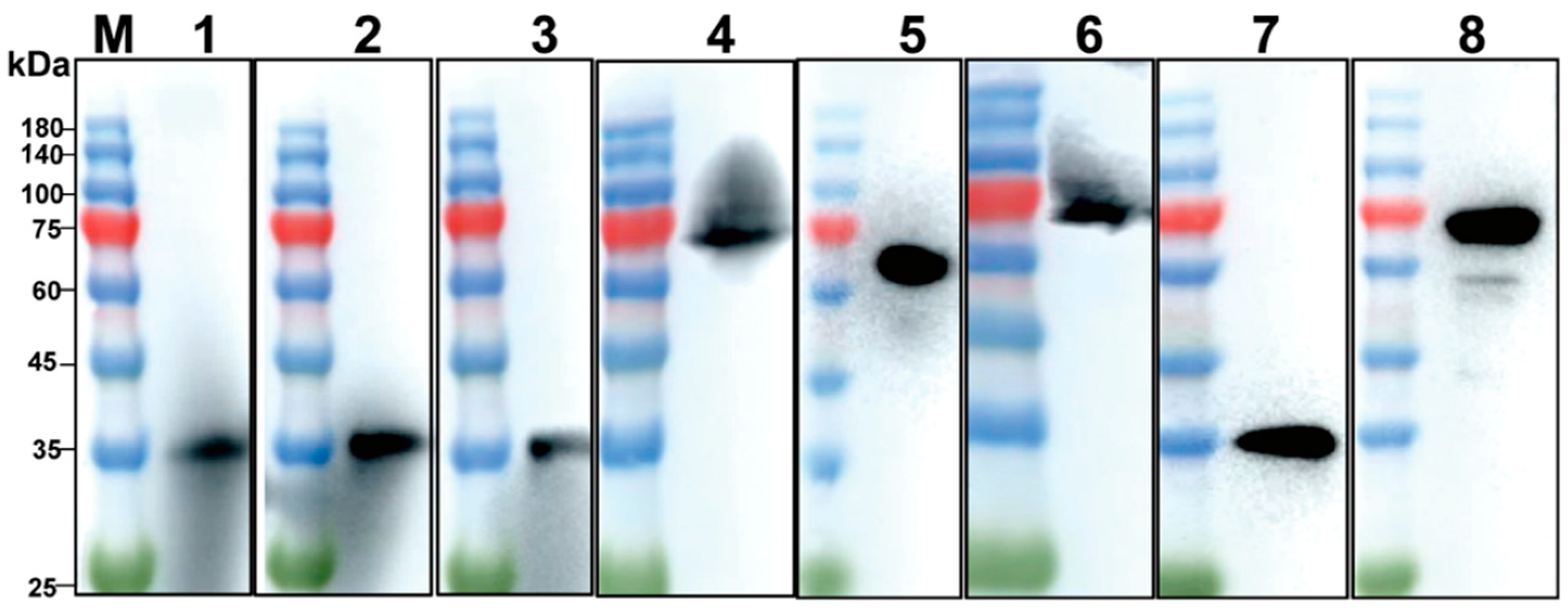
3.1. Expression and Purification of p30 and pB602L
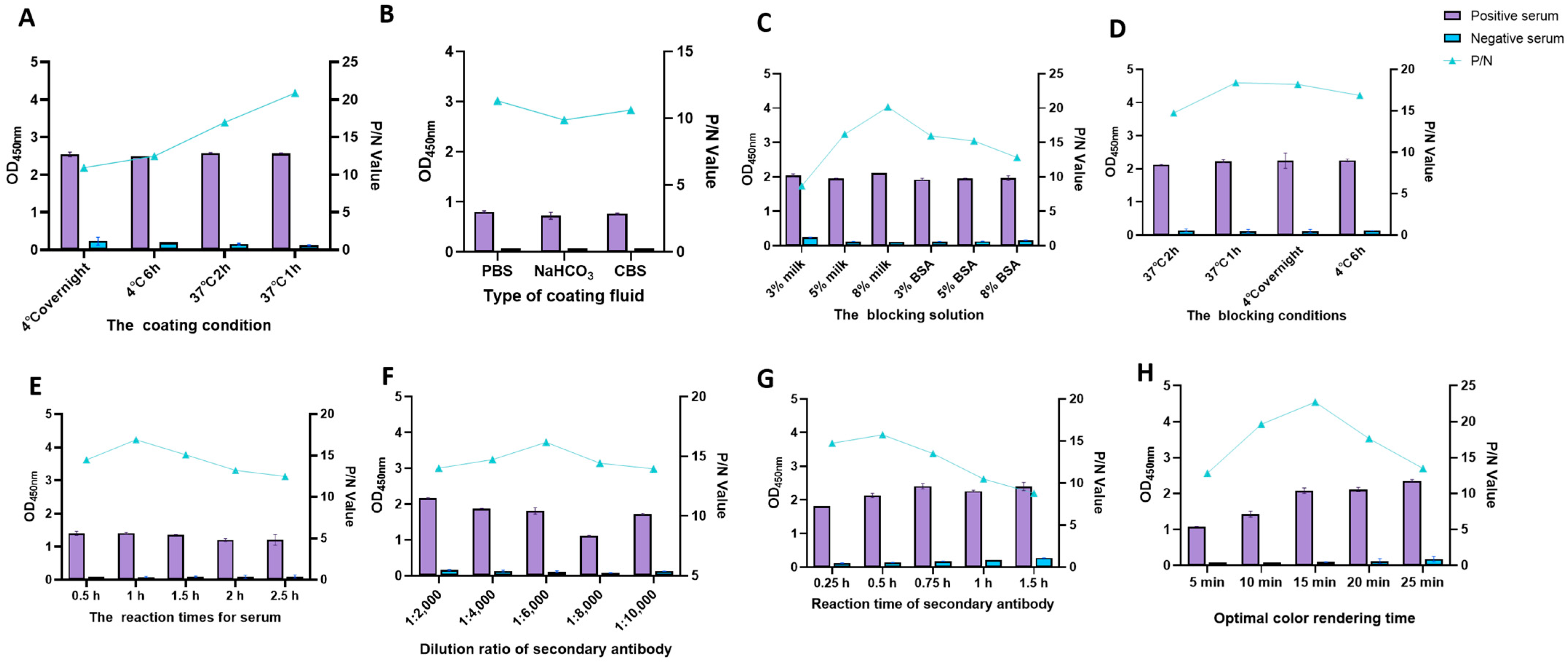
3.2. Optimization of Optimal Conditions for Indirect ELISA with Dual-Antigen
3.3. Determination of Cutoff Values
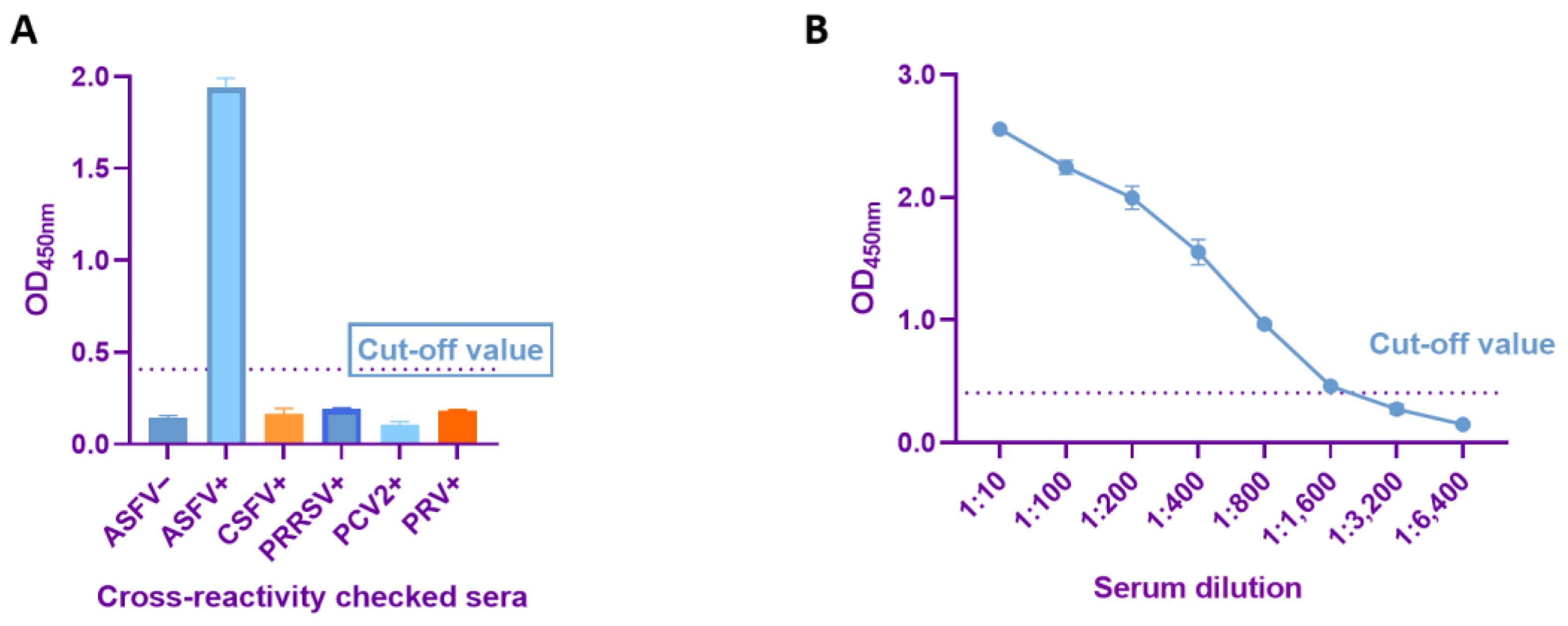
3.4. Sensitivity and Specificity Tests of Dual-Antigen Indirect ELISA
3.5. Repeatability Test
3.6. Detection of Clinical Serum Samples
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, G.; Xie, M.; Wu, W.; Chen, Z. Structures and functional diversities of asfv proteins. Viruses 2021, 13, 2124. [Google Scholar] [CrossRef]
- Sanchez, E.G.; Quintas, A.; Nogal, M.; Castello, A.; Revilla, Y. African swine fever virus controls the host transcription and cellular machinery of protein synthesis. Virus Res. 2013, 173, 58–75. [Google Scholar] [CrossRef]
- Wang, N.; Zhao, D.; Wang, J.; Zhang, Y.; Wang, M.; Gao, Y.; Li, F.; Wang, J.; Bu, Z.; Rao, Z.; et al. Architecture of african swine fever virus and implications for viral assembly. Science 2019, 366, 640–644. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African swine fever virus: An emerging DNA arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef]
- Li, Z.; Chen, W.; Qiu, Z.; Li, Y.; Fan, J.; Wu, K.; Li, X.; Zhao, M.; Ding, H.; Fan, S.; et al. African swine fever virus: A review. Life 2022, 12, 1255. [Google Scholar] [CrossRef]
- Duan, X.; Ru, Y.; Yang, W.; Ren, J.; Hao, R.; Qin, X.; Li, D.; Zheng, H. Research progress on the proteins involved in african swine fever virus infection and replication. Front. Immunol. 2022, 13, 947180. [Google Scholar] [CrossRef] [PubMed]
- Alejo, A.; Matamoros, T.; Guerra, M.; Andres, G. A proteomic atlas of the african swine fever virus particle. J. Virol. 2018, 92, e01293-18. [Google Scholar] [CrossRef] [PubMed]
- Kazakova, A.S.; Imatdinov, I.R.; Dubrovskaya, O.A.; Imatdinov, A.R.; Sidlik, M.V.; Balyshev, V.M.; Krasochko, P.A.; Sereda, A.D. Recombinant protein p30 for serological diagnosis of african swine fever by immunoblotting assay. Transbound. Emerg. Dis. 2017, 64, 1479–1492. [Google Scholar] [CrossRef]
- Wang, M.; Song, J.; Sun, J.; Du, Y.; Qin, X.; Xia, L.; Wu, Y.; Zhang, G. Development of an effective double antigen sandwich elisa based on p30 protein to detect antibodies against african swine fever virus. Viruses 2022, 14, 2170. [Google Scholar] [CrossRef] [PubMed]
- Gan, S.D.; Patel, K.R. Enzyme immunoassay and enzyme-linked immunosorbent assay. J. Investig. Dermatol. 2013, 133, e12. [Google Scholar] [CrossRef] [PubMed]
- Awosanya, E.J.; Olugasa, B.O.; Gimba, F.I.; Sabri, M.Y.; Ogundipe, G.A. Detection of african swine fever virus in pigs in southwest nigeria. Vet. World 2021, 14, 1840–1845. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.L.; Andres, G. African swine fever virus morphogenesis. Virus Res. 2013, 173, 29–41. [Google Scholar] [CrossRef]
- Wang, P.; Liu, C.; Wang, S.; Wen, L.; Shi, Z.; Chi, Y.; Wang, M.; Liu, Z.; Sun, Z.; Wei, L.; et al. Production and application of mouse monoclonal antibodies targeting linear epitopes in pb602l of african swine fever virus. Arch. Virol. 2022, 167, 415–424. [Google Scholar] [CrossRef]
- Onzere, C.K.; Bastos, A.D.; Okoth, E.A.; Lichoti, J.K.; Bochere, E.N.; Owido, M.G.; Ndambuki, G.; Bronsvoort, M.; Bishop, R.P. Multi-locus sequence typing of african swine fever viruses from endemic regions of kenya and eastern uganda (2011–2013) reveals rapid b602l central variable region evolution. Virus Genes 2018, 54, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xia, Q.; Sun, Q.; Zhang, Y.; Li, Y.; Ma, X.; Guan, Z.; Zhang, J.; Li, Z.; Liu, K.; et al. Detection of african swine fever virus antibodies in serum using a pb602l protein-based indirect elisa. Front. Vet. Sci. 2022, 9, 971841. [Google Scholar] [CrossRef]
- Zhong, K.; Zhu, M.; Yuan, Q.; Deng, Z.; Feng, S.; Liu, D.; Yuan, X. Development of an indirect elisa to detect african swine fever virus pp62 protein-specific antibodies. Front. Vet. Sci. 2021, 8, 798559. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, C.; Gomez-Sebastian, S.; Moreno, N.; Nunez, M.C.; Mulumba-Mfumu, L.K.; Quembo, C.J.; Heath, L.; Etter, E.M.; Jori, F.; Escribano, J.M.; et al. African swine fever virus serodiagnosis: A general review with a focus on the analyses of african serum samples. Virus Res. 2013, 173, 159–167. [Google Scholar] [CrossRef]
- Li, J.; Jiao, J.; Liu, N.; Ren, S.; Zeng, H.; Peng, J.; Zhang, Y.; Guo, L.; Liu, F.; Lv, T.; et al. Novel p22 and p30 dual-proteins combination based indirect elisa for detecting antibodies against african swine fever virus. Front. Vet. Sci. 2023, 10, 1093440. [Google Scholar] [CrossRef]
- Bao, J.; Wang, Q.; Lin, P.; Liu, C.; Li, L.; Wu, X.; Chi, T.; Xu, T.; Ge, S.; Liu, Y.; et al. Genome comparison of african swine fever virus china/2018/anhuixcgq strain and related european p72 genotype ii strains. Transbound. Emerg. Dis. 2019, 66, 1167–1176. [Google Scholar] [CrossRef]
- Wang, T.; Sun, Y.; Qiu, H.J. African swine fever: An unprecedented disaster and challenge to china. Infect. Dis. Poverty 2018, 7, 111. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, H.; Hou, L.; Yang, C.; Wen, Y. Advance of african swine fever virus in recent years. Res. Vet. Sci. 2021, 136, 535–539. [Google Scholar] [CrossRef]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernandez-Pinero, J.; Sanchez-Vizcaino, J.M. Gaps in african swine fever: Analysis and priorities. Transbound. Emerg. Dis. 2018, 65 (Suppl. S1), 235–247. [Google Scholar] [CrossRef]
- Alcaraz, C.; Rodriguez, F.; Oviedo, J.M.; Eiras, A.; De Diego, M.; Alonso, C.; Escribano, J.M. Highly specific confirmatory western blot test for african swine fever virus antibody detection using the recombinant virus protein p54. J. Virol. Methods 1995, 52, 111–119. [Google Scholar] [CrossRef]
- Gallardo, C.; Blanco, E.; Rodriguez, J.M.; Carrascosa, A.L.; Sanchez-Vizcaino, J.M. Antigenic properties and diagnostic potential of african swine fever virus protein pp62 expressed in insect cells. J. Clin. Microbiol. 2006, 44, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Sastre, P.; Perez, T.; Costa, S.; Yang, X.; Raber, A.; Blome, S.; Goller, K.V.; Gallardo, C.; Tapia, I.; Garcia, J.; et al. Development of a duplex lateral flow assay for simultaneous detection of antibodies against african and classical swine fever viruses. J. Vet. Diagn. Investig. 2016, 28, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Achenbach, J.E.; Gallardo, C.; Nieto-Pelegrin, E.; Rivera-Arroyo, B.; Degefa-Negi, T.; Arias, M.; Jenberie, S.; Mulisa, D.D.; Gizaw, D.; Gelaye, E.; et al. Identification of a new genotype of african swine fever virus in domestic pigs from ethiopia. Transbound. Emerg. Dis. 2017, 64, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Soler, A.; Nieto, R.; Carrascosa, A.L.; De Mia, G.M.; Bishop, R.P.; Martins, C.; Fasina, F.O.; Couacy-Hymman, E.; Heath, L.; et al. Comparative evaluation of novel african swine fever virus (asf) antibody detection techniques derived from specific asf viral genotypes with the oie internationally prescribed serological tests. Vet. Microbiol. 2013, 162, 32–43. [Google Scholar] [CrossRef]
- Le, V.P.; Jeong, D.G.; Yoon, S.W.; Kwon, H.M.; Trinh, T.B.N.; Nguyen, T.L.; Bui, T.T.N.; Oh, J.; Kim, J.B.; Cheong, K.M.; et al. Outbreak of african swine fever, vietnam, 2019. Emerg. Infect. Dis. 2019, 25, 1433–1435. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernandez-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simon, A.; et al. Assessment of african swine fever diagnostic techniques as a response to the epidemic outbreaks in eastern european union countries: How to improve surveillance and control programs. J. Clin. Microbiol. 2015, 53, 2555–2565. [Google Scholar] [CrossRef]
- Oviedo, J.; Rodríguez, F.; Gómez-Puertas, P.; Brun, A.; Gómez, N.; Alonso, C.; Escribano, J. High level expression of the major antigenic african swine fever virus proteins p54 and p30 in baculovirus and their potential use as diagnostic reagents. J. Virol. Methods 1997, 64, 27–35. [Google Scholar] [CrossRef]
- Nah, J.J.; Kwon, O.K.; Choi, J.D.; Jang, S.H.; Lee, H.J.; Ahn, D.G.; Lee, K.; Kang, B.; Hae-Eun, K.; Shin, Y.K. Development of an indirect elisa against african swine fever virus using two recombinant antigens, partial p22 and p30. J. Virol. Methods 2022, 309, 114611. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Lowe, A.D.; Rodriguez, Y.Y.; Murgia, M.V.; Dodd, K.A.; Rowland, R.R.; Jia, W. Antigenic regions of african swine fever virus phosphoprotein p30. Transbound. Emerg. Dis. 2020; online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Lokhandwala, S.; Waghela, S.D.; Bray, J.; Sangewar, N.; Charendoff, C.; Martin, C.L.; Hassan, W.S.; Koynarski, T.; Gabbert, L.; Burrage, T.G.; et al. Adenovirus-vectored novel african swine fever virus antigens elicit robust immune responses in swine. PLoS ONE 2017, 12, e0177007. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Castaneda, B.; Reis, A.L.; Corteyn, A.; Parkhouse, R.M.; Kollnberger, S. Expression, cellular localization and antibody responses of the african swine fever virus genes b602l and k205r. Arch. Virol. 2008, 153, 2303–2306. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Primers Sequences |
|---|---|
| CP204L-F | AGCTTGTCGACGGAGCTCGAATTCTTATTTTTTTTTTAA |
| CP204L-R | CAAGGCCATGGCTGATATCGGATCCATGGATTTTATTTTA |
| B602L-F | TGGTGGTGCTCGAGTGCGGCCGCCAATTCTGCTTT |
| B602L-R | ATGGGTCGCGGATCCGAATTCATGGCAGAATTTAA |
| Dilution of Sera | Antigen at Different Concentrations (µg/mL) | ||||||
|---|---|---|---|---|---|---|---|
| 0.1 | 0.2 | 0.4 | 0.6 | 0.8 | 1 | ||
| 1:10 | P | 0.847 | 1.273 | 1.691 | 1.961 | 2.168 | 2.105 |
| N | 0.159 | 0.180 | 0.362 | 0.249 | 0.236 | 0.251 | |
| P/N | 5.333 | 7.084 | 4.669 | 7.876 | 9.208 | 8.394 | |
| 1:50 | P | 0.503 | 0.813 | 1.413 | 1.840 | 1.905 | 2.031 |
| N | 0.103 | 0.084 | 0.086 | 0.104 | 0.114 | 0.140 | |
| P/N | 4.871 | 9.729 | 16.527 | 17.680 | 16.686 | 14.562 | |
| 1:100 | P | 0.381 | 0.691 | 1.337 | 1.724 | 1.780 | 1.851 |
| N | 0.060 | 0.171 | 0.082 | 0.076 | 0.081 | 0.096 | |
| P/N | 6.304 | 4.038 | 16.263 | 22.571 | 22.100 | 19.376 | |
| 1:200 | P | 0.308 | 0.603 | 1.013 | 1.621 | 1.546 | 1.567 |
| N | 0.057 | 0.063 | 0.057 | 0.066 | 0.071 | 0.072 | |
| P/N | 5.385 | 9.579 | 17.744 | 24.732 | 21.935 | 21.729 | |
| 1:400 | P | 0.226 | 0.349 | 0.616 | 1.141 | 1.094 | 1.096 |
| N | 0.066 | 0.094 | 0.056 | 0.084 | 0.066 | 0.084 | |
| P/N | 3.406 | 3.697 | 11.012 | 13.523 | 16.683 | 12.991 | |
| 1:600 | P | 0.179 | 0.313 | 0.528 | 0.940 | 0.984 | 1.067 |
| N | 0.056 | 0.064 | 0.058 | 0.057 | 0.060 | 0.068 | |
| P/N | 3.219 | 4.900 | 9.038 | 16.447 | 16.388 | 15.721 | |
| 1:800 | P | 0.175 | 0.305 | 0.443 | 0.950 | 0.921 | 0.952 |
| N | 0.056 | 0.058 | 0.064 | 0.062 | 0.060 | 0.061 | |
| P/N | 3.113 | 5.287 | 6.972 | 15.277 | 15.284 | 15.518 | |
| Volume Ratio of p30 to pB602L | 5:1 | 4:1 | 3:1 | 2:1 | 1:1 | 1:2 | 1:3 | 1:4 | 1:5 |
|---|---|---|---|---|---|---|---|---|---|
| P | 1.061 | 1.024 | 0.914 | 0.834 | 0.675 | 0.500 | 0.502 | 0.424 | 0.367 |
| N | 0.090 | 0.056 | 0.059 | 0.055 | 0.084 | 0.057 | 0.080 | 0.066 | 0.063 |
| P/N | 11.783 | 18.204 | 15.564 | 15.231 | 8.045 | 8.713 | 6.274 | 6.442 | 5.800 |
| Sample No. | Intra-Assay CV (%) | Inter-Assay CV (%) | |||
|---|---|---|---|---|---|
| X ± SD | CV (%) | X ± SD | CV (%) | ||
| Positive samples | 1 | 1.689 ± 0.070 | 4.14 | 1.492 ± 0.064 | 4.32 |
| 2 | 1.842 ± 0.019 | 1.02 | 1.424 ± 0.043 | 3.04 | |
| 3 | 1.571 ± 0.057 | 3.65 | 1.455 ± 0.069 | 4.71 | |
| 4 | 1.918 ± 0.047 | 2.44 | 1.493 ± 0.063 | 4.21 | |
| 5 | 1.962 ± 0.040 | 2.04 | 1.458 ± 0.062 | 4.24 | |
| Negative samples | 6 | 0.107 ± 0.005 | 4.96 | 0.110 ± 0.006 | 5.87 |
| 7 | 0.073 ± 0.003 | 4.39 | 0.068 ± 0.003 | 5.11 | |
| 8 | 0.085 ± 0.003 | 3.95 | 0.091 ± 0.006 | 6.98 | |
| 9 | 0.087 ± 0.002 | 2.28 | 0.070 ± 0.006 | 8.98 | |
| 10 | 0.083 ± 0.004 | 4.44 | 0.071 ± 0.005 | 7.05 | |
| No. of Clinical Samples | Dual-Antigen Indirect ELISA | Commercial Kits | ||
|---|---|---|---|---|
| No. of Positive | Positive Rate (%) | No. of Positive | Positive Rate (%) | |
| 106 | 17 | 16.1% | 14 | 13.3% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Song, J.; Wang, M.; Sun, Z.; Sun, J.; Tian, P.; Zhuang, G.; Zhang, A.; Wu, Y.; Zhang, G. Establishment of a Dual-Antigen Indirect ELISA Based on p30 and pB602L to Detect Antibodies against African Swine Fever Virus. Viruses 2023, 15, 1845. https://doi.org/10.3390/v15091845
Zhou L, Song J, Wang M, Sun Z, Sun J, Tian P, Zhuang G, Zhang A, Wu Y, Zhang G. Establishment of a Dual-Antigen Indirect ELISA Based on p30 and pB602L to Detect Antibodies against African Swine Fever Virus. Viruses. 2023; 15(9):1845. https://doi.org/10.3390/v15091845
Chicago/Turabian StyleZhou, Lei, Jinxing Song, Mengxiang Wang, Zhuoya Sun, Junru Sun, Panpan Tian, Guoqing Zhuang, Angke Zhang, Yanan Wu, and Gaiping Zhang. 2023. "Establishment of a Dual-Antigen Indirect ELISA Based on p30 and pB602L to Detect Antibodies against African Swine Fever Virus" Viruses 15, no. 9: 1845. https://doi.org/10.3390/v15091845
APA StyleZhou, L., Song, J., Wang, M., Sun, Z., Sun, J., Tian, P., Zhuang, G., Zhang, A., Wu, Y., & Zhang, G. (2023). Establishment of a Dual-Antigen Indirect ELISA Based on p30 and pB602L to Detect Antibodies against African Swine Fever Virus. Viruses, 15(9), 1845. https://doi.org/10.3390/v15091845