
Pteropus vampyrus TRIM40 Is an Interferon-Stimulated Gene That Antagonizes RIG-I-like Receptors


Abstract
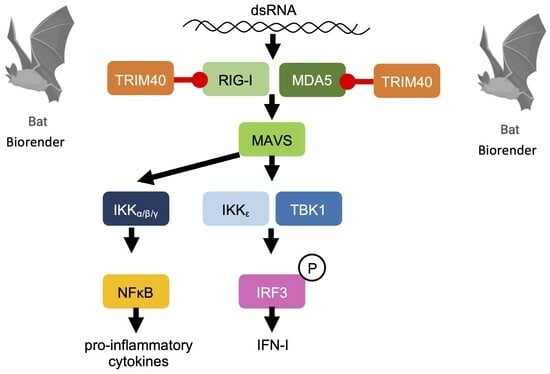
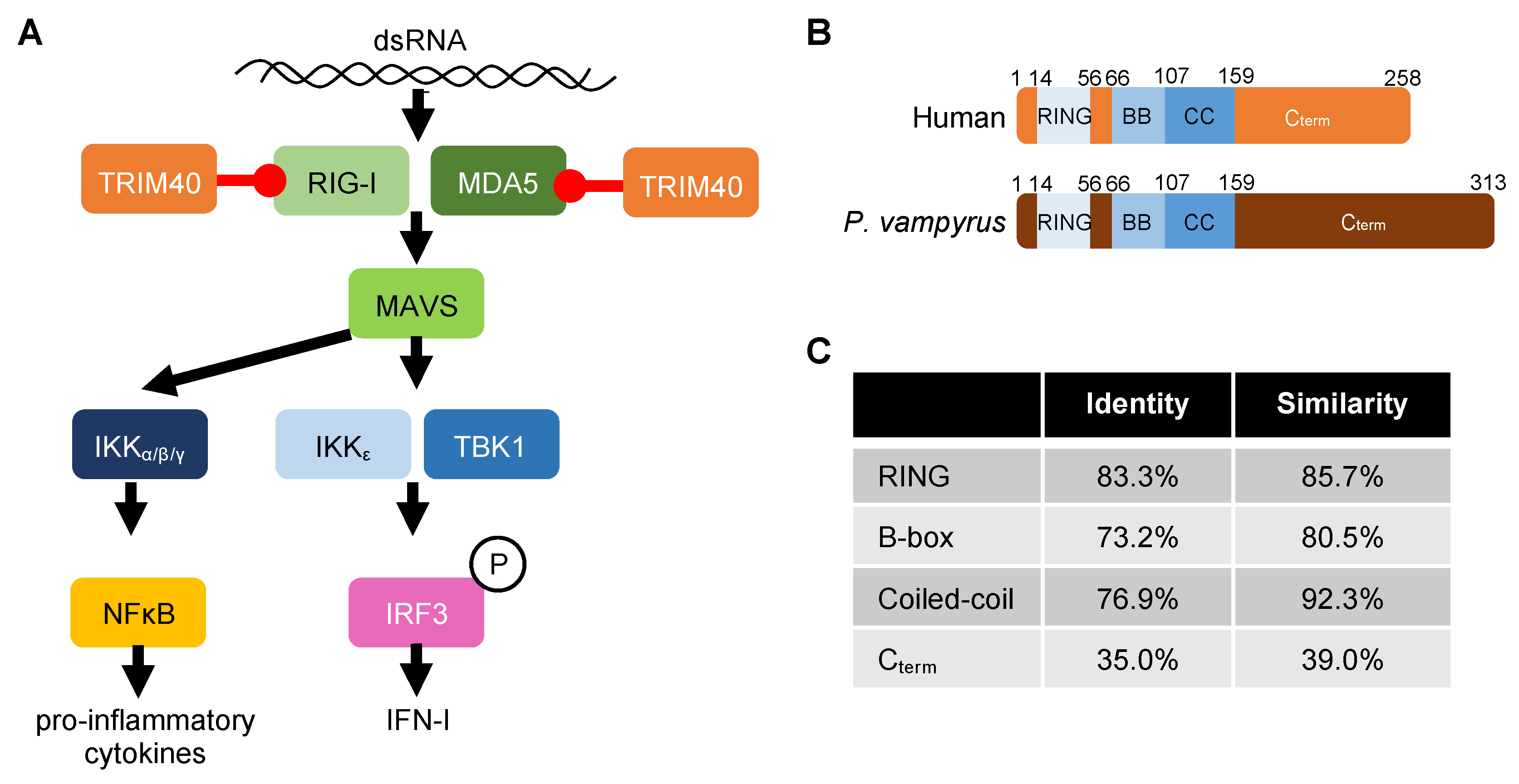
:
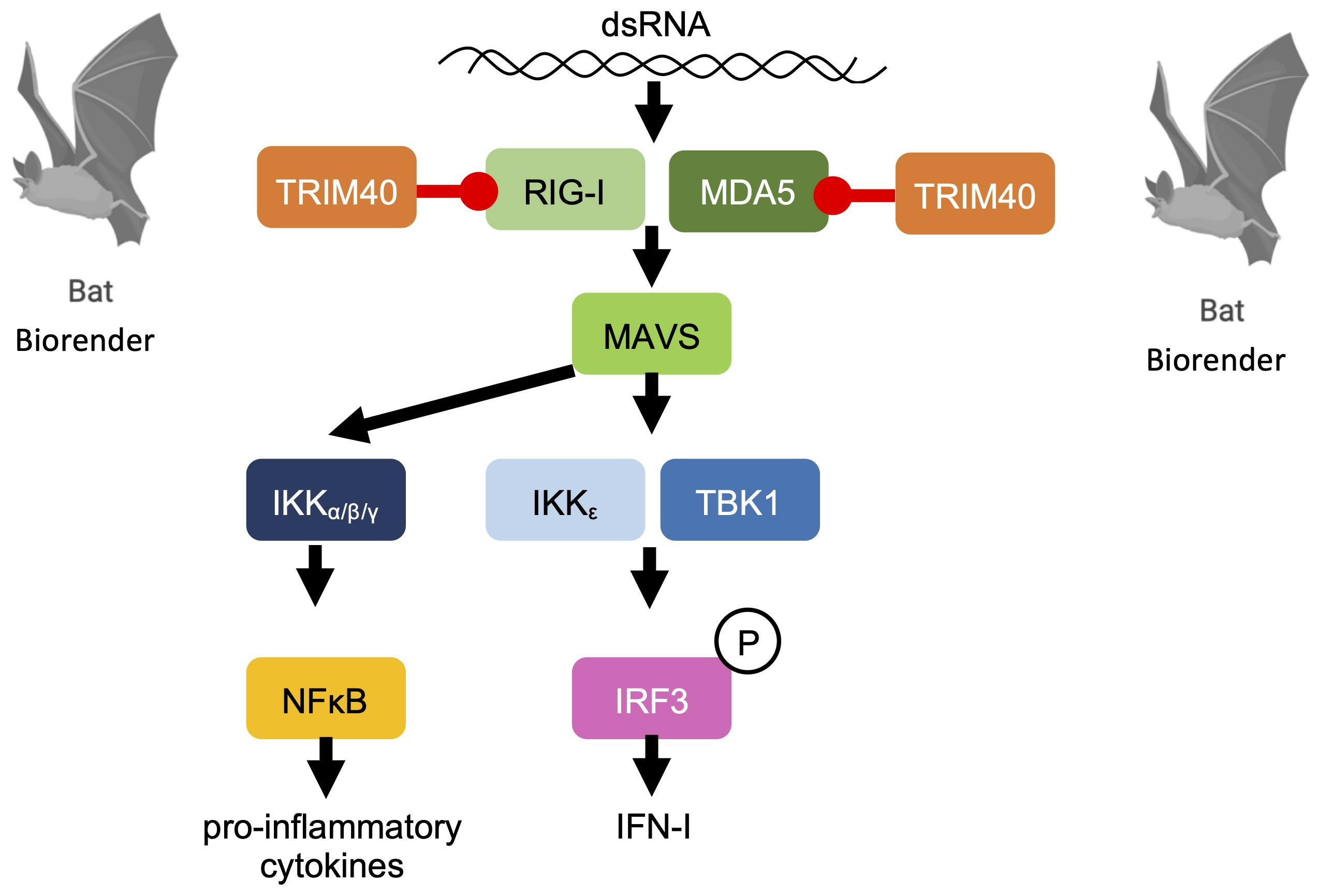
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Plasmids
2.3. Viruses
2.4. Virus Infections and Plaque Assays
2.5. Transfections and Immunoprecipitations
2.6. Protein Purification
2.7. Immune Stimulations
2.8. Western Blots
2.9. RNA Extractions and Quantitative PCR
2.10. Knockdown Experiments
2.11. Quantification and Statistical Analysis
3. Results
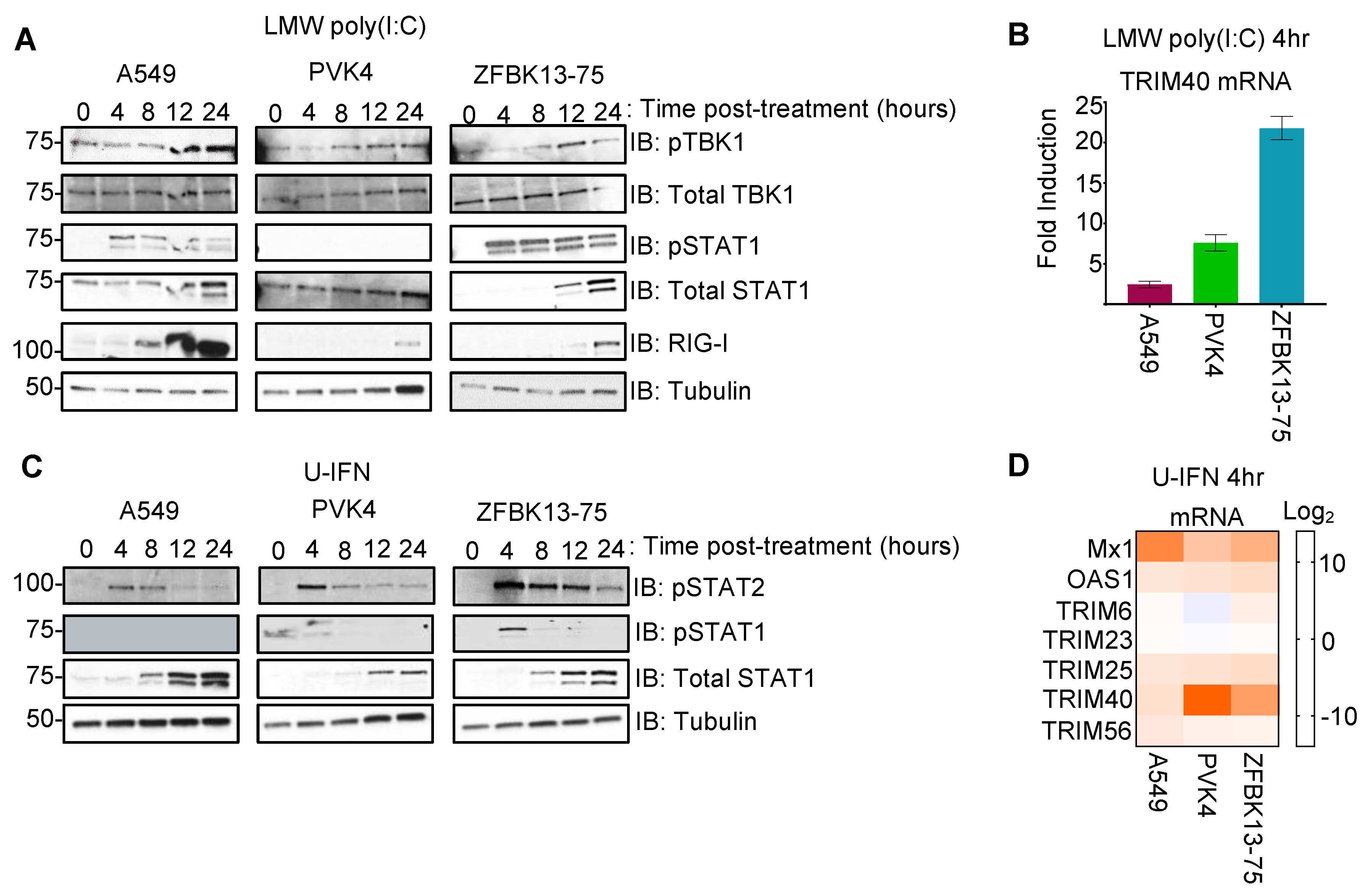
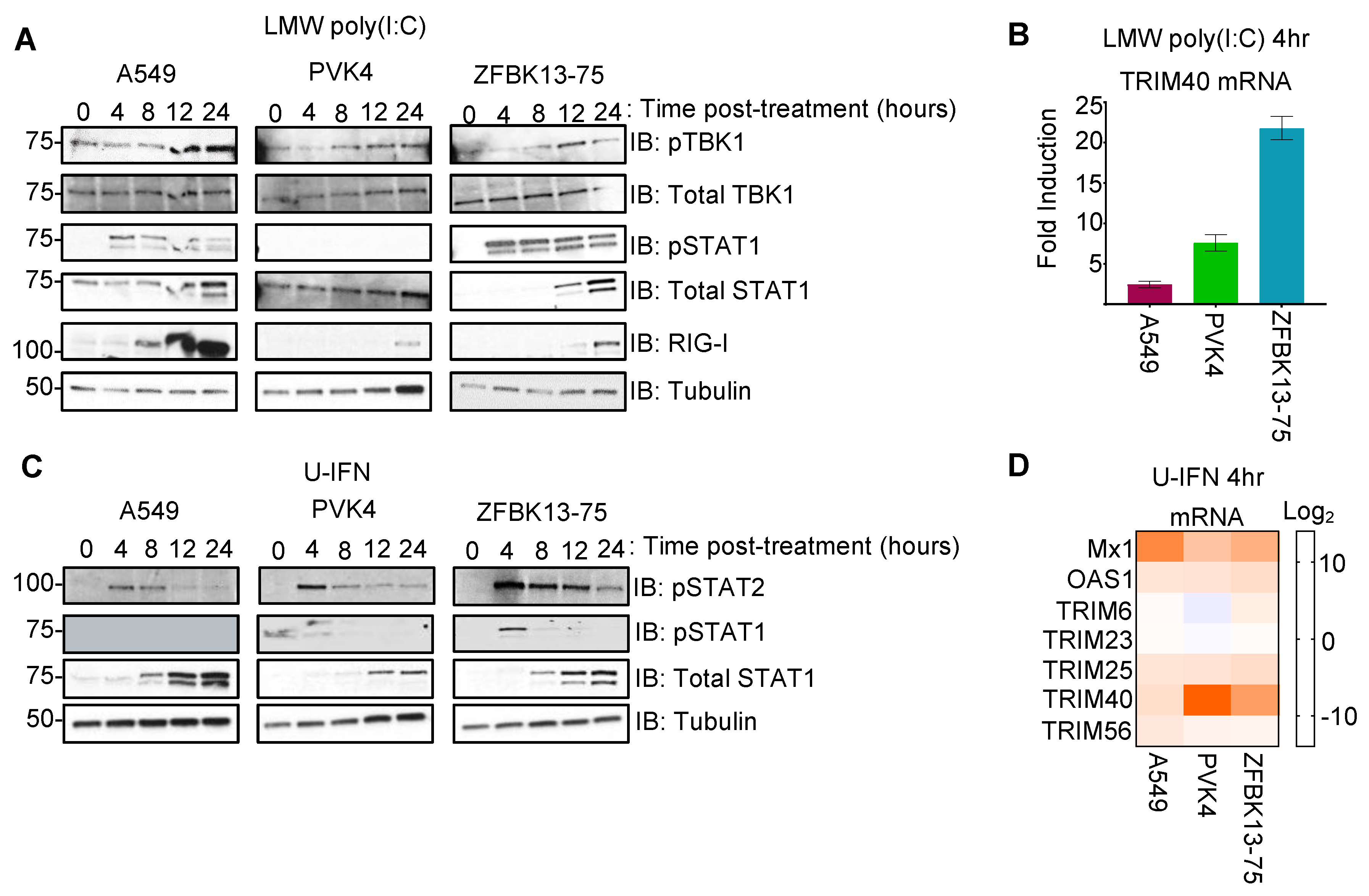
3.1. Pteropodid TRIM40 Is an IFN-Stimulated Gene (ISG)
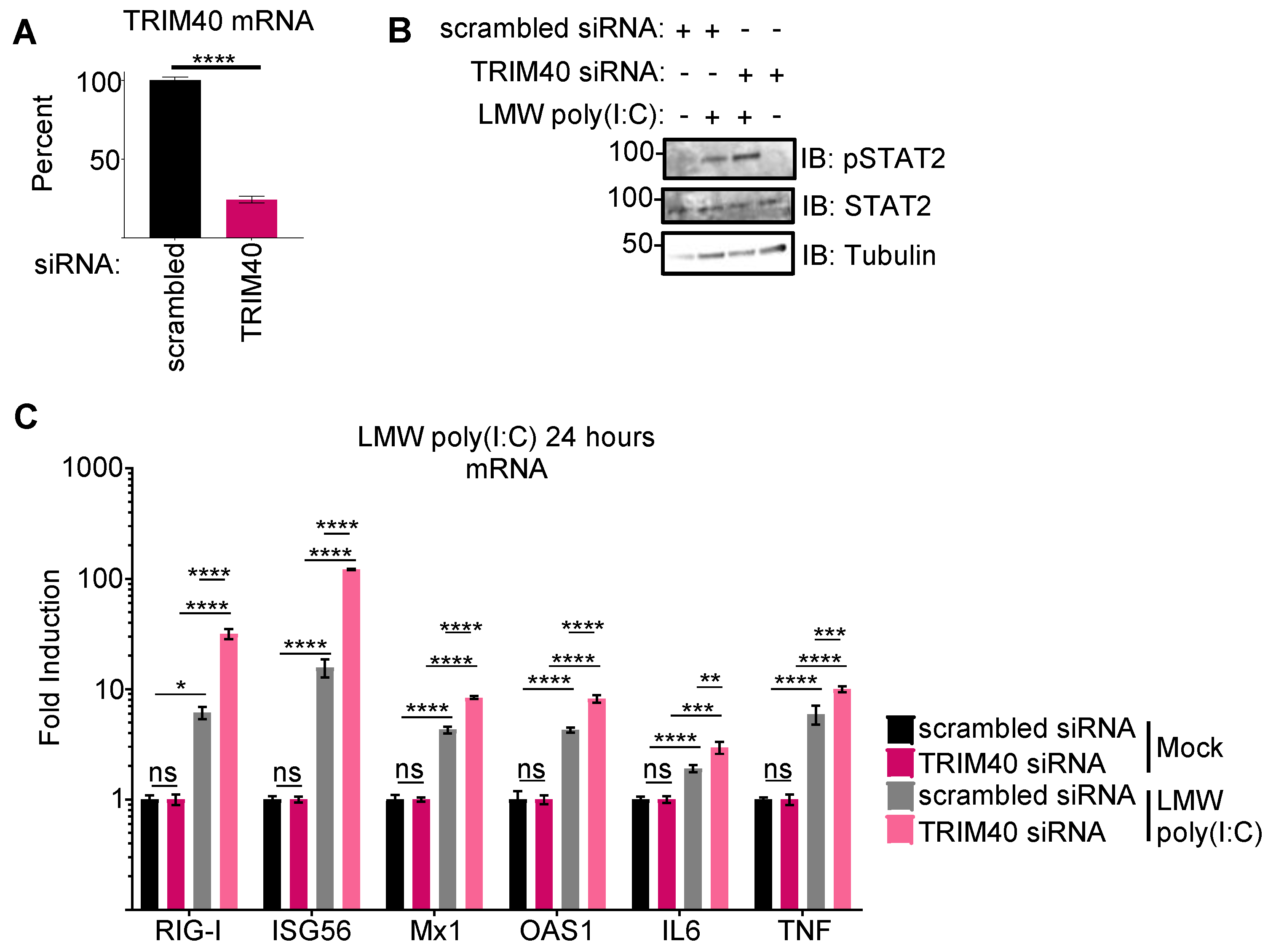
3.2. Pteropus vampyrus TRIM40 Antagonizes Type I Interferon and Pro-Inflammatory Cytokine Gene Expression
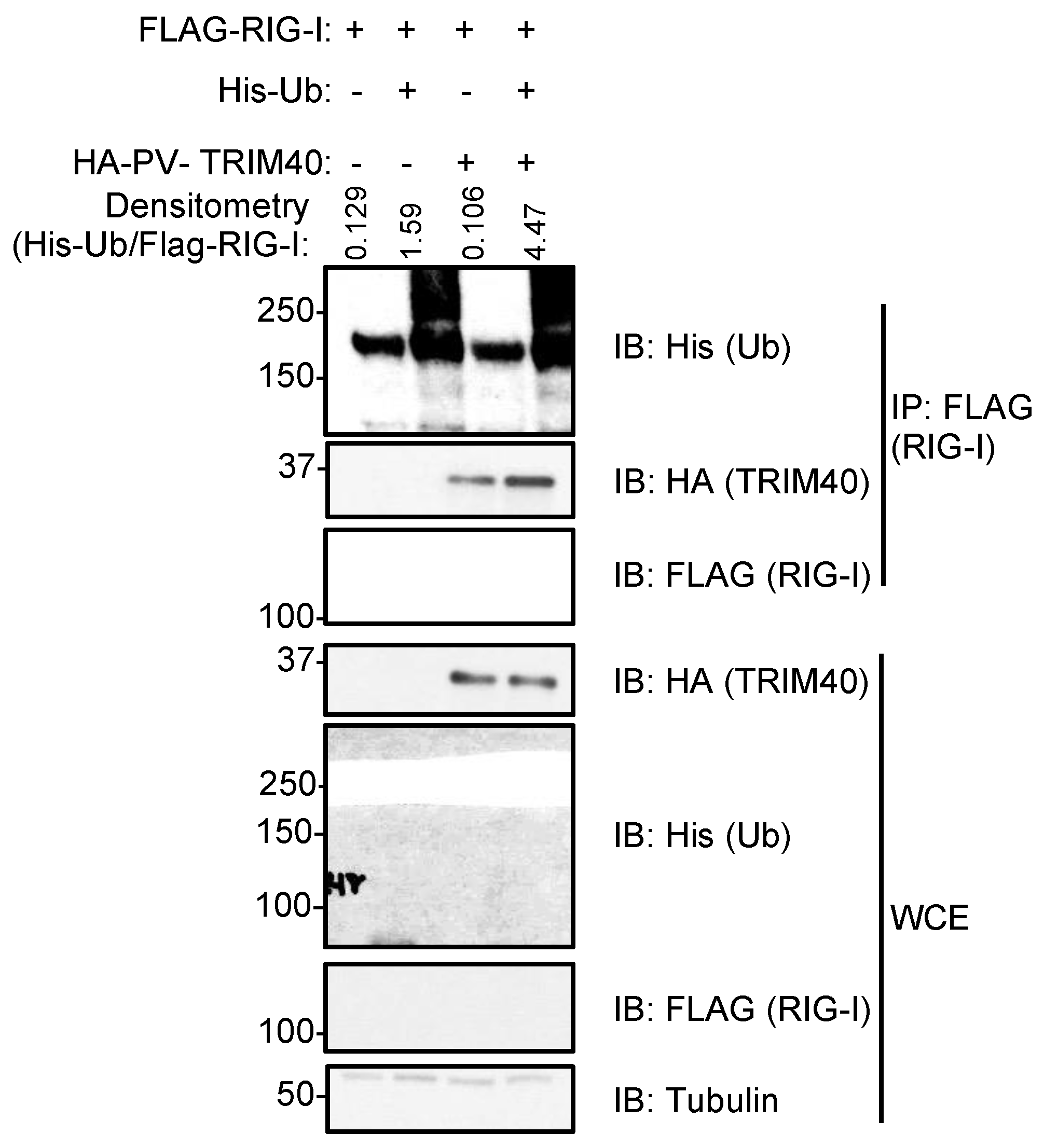
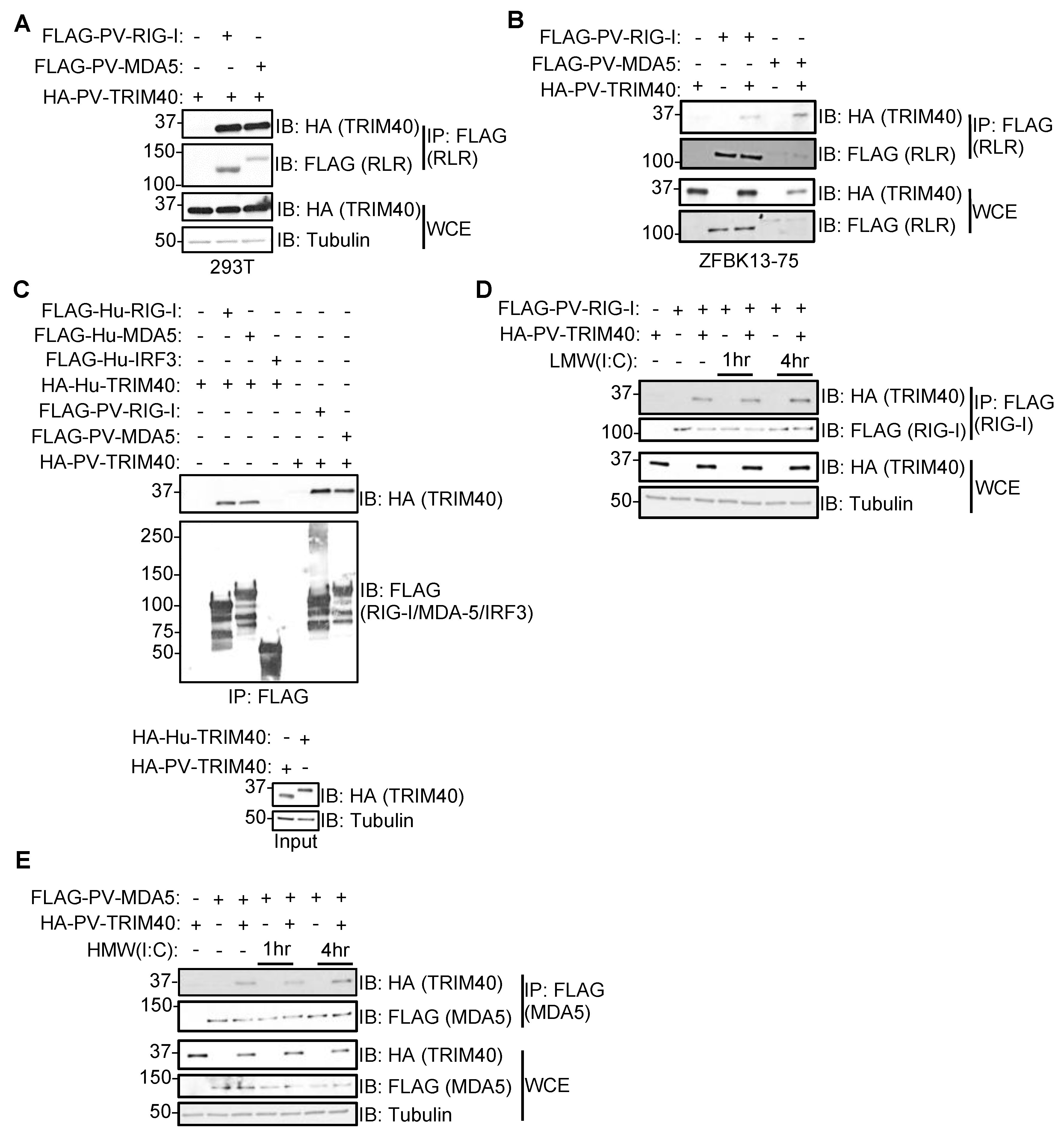
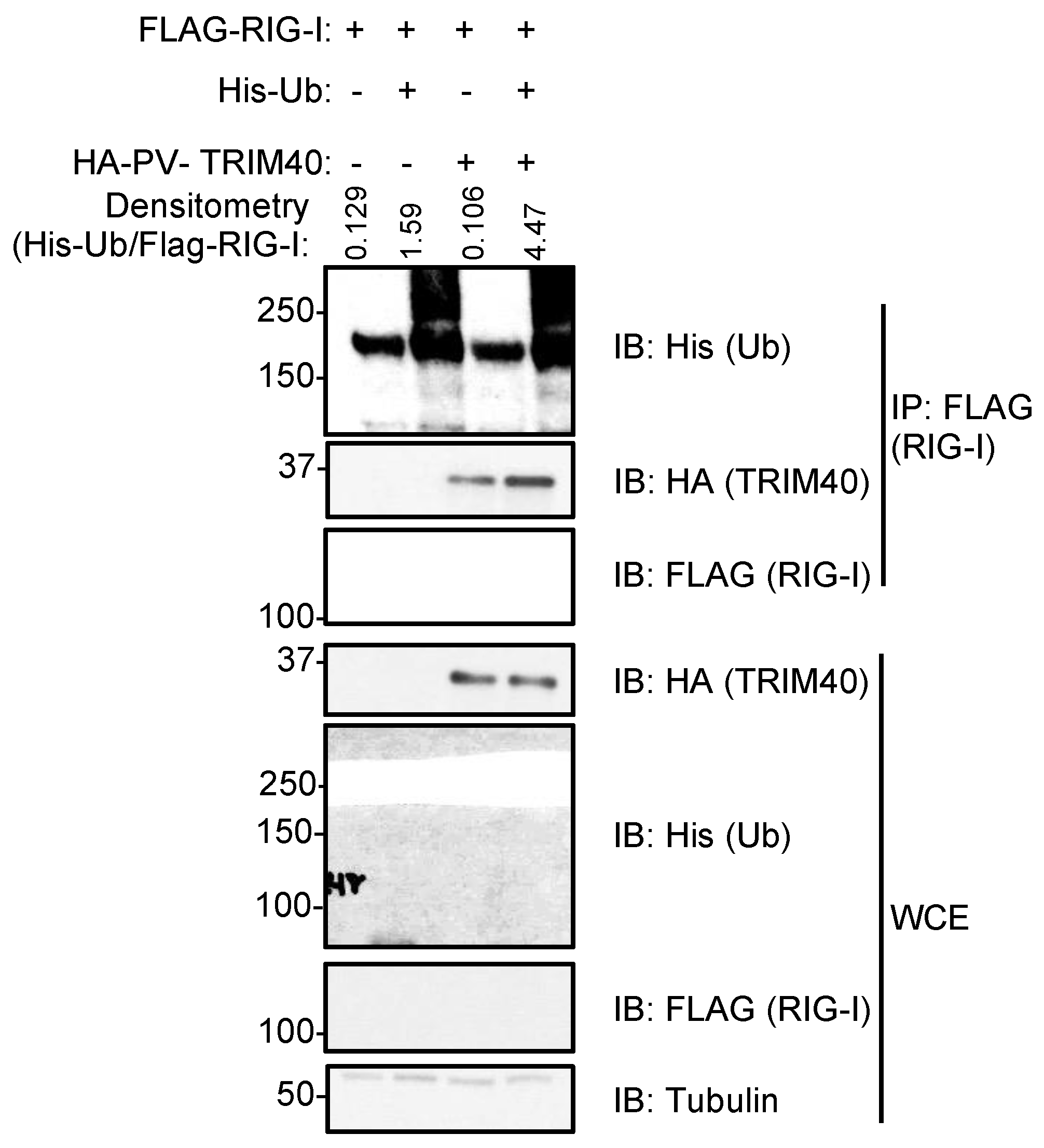
3.3. Pteropus vampyrus TRIM40 Interacts with and Ubiquitinates RIG-I-like Receptors RIG-I and MDA5
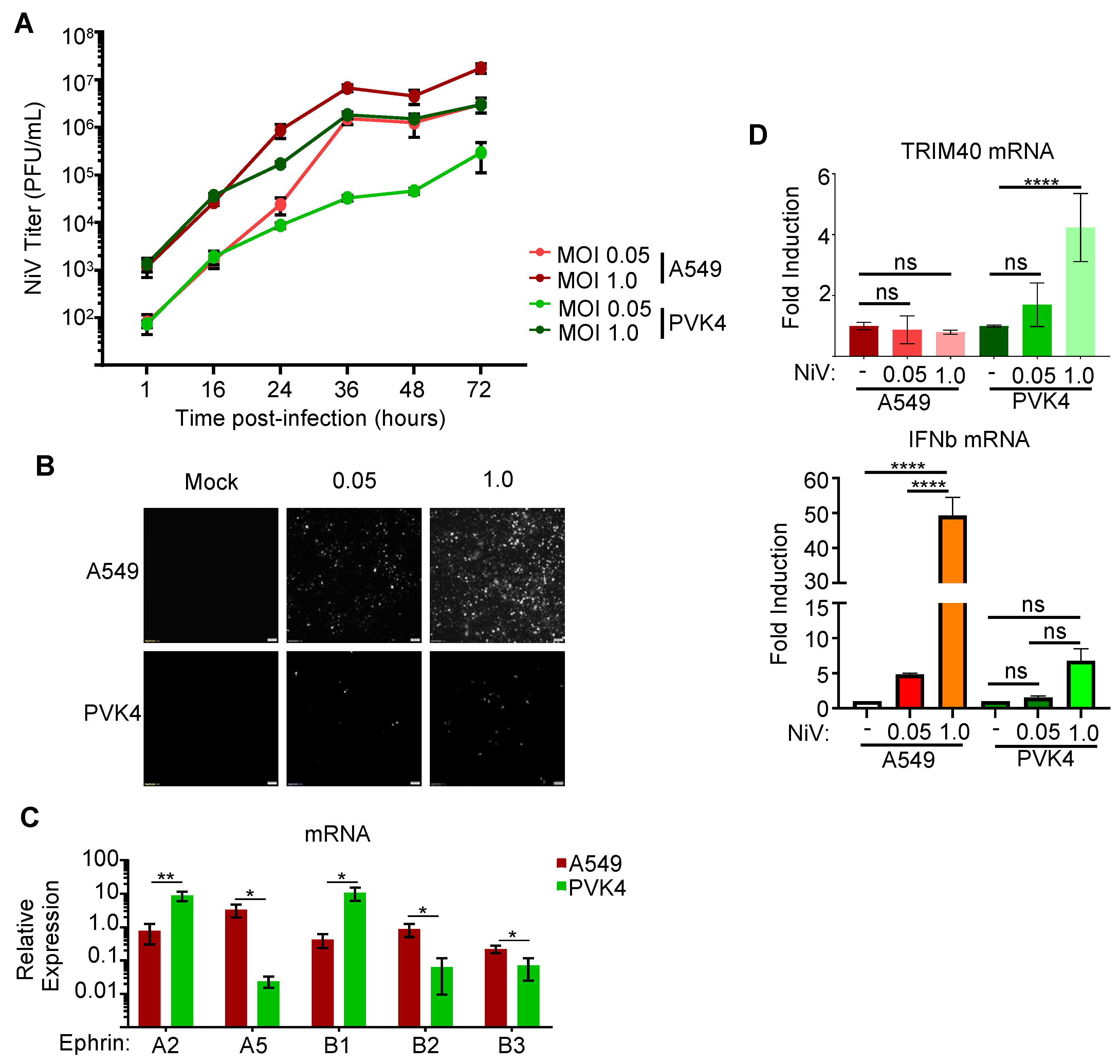
3.4. Nipah Virus Infection Induces TRIM40 Expression in Pteropus vampyrus Cells
3.5. TRIM40 Promotes Virus Replication in Bat and Human Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guth, S.; Mollentze, N.; Renault, K.; Streicker, D.G.; Visher, E.; Boots, M.; Brook, C.E. Bats host the most virulent-but not the most dangerous-zoonotic viruses. Proc. Natl. Acad. Sci. USA 2022, 119, e2113628119. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.J.; Morrissy, C.J.; van der Heide, B.M.; Russell, G.M.; Braun, M.A.; Westbury, H.A.; Halpin, K.; Daniels, P.W. Experimental Nipah virus infection in pteropid bats (Pteropus poliocephalus). J. Comp. Pathol. 2007, 136, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Halpin, K.; Hyatt, A.D.; Fogarty, R.; Middleton, D.; Bingham, J.; Epstein, J.H.; Rahman, S.A.; Hughes, T.; Smith, C.; Field, H.E.; et al. Pteropid bats are confirmed as the reservoir hosts of henipaviruses: A comprehensive experimental study of virus transmission. Am. J. Trop. Med. Hyg. 2011, 85, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Woon, A.; Boyd, V.; Todd, S.; Smith, I.; Klein, R.; Woodhouse, I.; Riddell, S.; Crameri, G.; Bingham, J.; Wang, L.; et al. Acute experimental infection of bats and ferrets with Hendra virus: Insights into the early host response of the reservoir host and susceptible model species. PLoS Pathog. 2020, 16, e1008412. [Google Scholar] [CrossRef]
- Chua, K.B.; Bellini, W.J.; Rota, P.A.; Harcourt, B.H.; Tamin, A.; Lam, S.K.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Shieh, W.; et al. Nipah virus: A recently emergent deadly paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef] [PubMed]
- Selvey, L.A.; Wells, R.M.; McCormack, J.G.; Ansford, A.J.; Murray, K.; Rogers, R.J.; Lavercombe, P.S.; Selleck, P.; Sheridan, J.W. Infection of humans and horses by a newly described morbillivirus. Med. J. Aust. 1995, 162, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, B.A.; Cross, R.W.; Fenton, K.A.; Agans, K.N.; Basler, C.F.; Geisbert, T.W.; Mire, C.E. The immunomodulating V and W proteins of Nipah virus determine disease course. Nat. Commun. 2015, 6, 7483. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, B.; Weingartl, H.M. Nipah virus infects specific subsets of porcine peripheral blood mononuclear cells. PLoS ONE 2012, 7, e30855. [Google Scholar] [CrossRef]
- Pelissier, R.; Iampietro, M.; Horvat, B. Recent advances in the understanding of Nipah virus immunopathogenesis and anti-viral approaches. F1000Research 2019, 8, 1763. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Bobb, K.; Borisevich, V.; Geisbert, J.B.; Agans, K.N.; Cross, R.W.; Prasad, A.N.; Fenton, K.A.; Yu, H.; Fouts, T.R.; et al. A single dose investigational subunit vaccine for human use against Nipah virus and Hendra virus. Npj Vaccines 2021, 6, 23. [Google Scholar] [CrossRef]
- Dong, J.; Cross, R.W.; Doyle, M.P.; Kose, N.; Mousa, J.J.; Annand, E.J.; Borisevich, V.; Agans, K.N.; Sutton, R.; Nargi, R.; et al. Potent Henipavirus Neutralization by Antibodies Recognizing Diverse Sites on Hendra and Nipah Virus Receptor Binding Protein. Cell 2020, 183, 1536–1550.e17. [Google Scholar] [CrossRef]
- Amaya, M.; Broder, C.C. Vaccines to Emerging Viruses: Nipah and Hendra. Annu. Rev. Virol. 2020, 7, 447–473. [Google Scholar] [CrossRef] [PubMed]
- Burroughs, A.L.; Durr, P.A.; Boyd, V.; Graham, K.; White, J.R.; Todd, S.; Barr, J.; Smith, I.; Baverstock, G.; Meers, J.; et al. Hendra Virus Infection Dynamics in the Grey-Headed Flying Fox (Pteropus poliocephalus) at the Southern-Most Extent of Its Range: Further Evidence This Species Does Not Readily Transmit the Virus to Horses. PLoS ONE 2016, 11, e0155252. [Google Scholar] [CrossRef] [PubMed]
- Halpin, K.; Young, P.L.; Field, H.E.; Mackenzie, J.S. Isolation of Hendra virus from pteropid bats: A natural reservoir of Hendra virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar] [CrossRef]
- Chua, K.B.; Koh, C.L.; Hooi, P.S.; Wee, K.F.; Khong, J.H.; Chua, B.H.; Chan, Y.P.; Lim, M.E.; Lam, S.K. Isolation of Nipah virus from Malaysian Island flying-foxes. Microbes Infect. 2002, 4, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Goldspink, L.K.; Edson, D.W.; Vidgen, M.E.; Bingham, J.; Field, H.E.; Smith, C.S. Natural Hendra Virus Infection in Flying-Foxes—Tissue Tropism and Risk Factors. PLoS ONE 2015, 10, e0128835. [Google Scholar] [CrossRef]
- Reynes, J.M.; Counor, D.; Ong, S.; Faure, C.; Seng, V.; Molia, S.; Walston, J.; Georges-Courbot, M.C.; Deubel, V.; Sarthou, J.L. Nipah virus in Lyle’s flying foxes, Cambodia. Emerg. Infect. Dis. 2005, 11, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Sazzad, H.M.; Satter, S.M.; Sultana, S.; Hossain, M.J.; Hasan, M.; Rahman, M.; Campbell, S.; Cannon, D.L.; Stroher, U.; et al. Nipah Virus Transmission from Bats to Humans Associated with Drinking Traditional Liquor Made from Date Palm Sap, Bangladesh, 2011–2014. Emerg. Infect. Dis. 2016, 22, 664–670. [Google Scholar] [CrossRef]
- Plowright, R.K.; Field, H.E.; Smith, C.; Divljan, A.; Palmer, C.; Tabor, G.; Daszak, P.; Foley, J.E. Reproduction and nutritional stress are risk factors for Hendra virus infection in little red flying foxes (Pteropus scapulatus). Proc. Biol. Sci. 2008, 275, 861–869. [Google Scholar] [CrossRef]
- Hahn, M.B.; Epstein, J.H.; Gurley, E.S.; Islam, M.S.; Luby, S.P.; Daszak, P.; Patz, J.A. Roosting behaviour and habitat selection of Pteropus giganteusreveal potential links to Nipah virus epidemiology. J. Appl. Ecol. 2014, 51, 376–387. [Google Scholar] [CrossRef]
- Edson, D.; Field, H.; McMichael, L.; Vidgen, M.; Goldspink, L.; Broos, A.; Melville, D.; Kristoffersen, J.; de Jong, C.; McLaughlin, A.; et al. Routes of Hendra Virus Excretion in Naturally-Infected Flying-Foxes: Implications for Viral Transmission and Spillover Risk. PLoS ONE 2015, 10, e0140670. [Google Scholar] [CrossRef]
- Zhou, P.; Tachedjian, M.; Wynne, J.W.; Boyd, V.; Cui, J.; Smith, I.; Cowled, C.; Ng, J.H.; Mok, L.; Michalski, W.P.; et al. Contraction of the type I IFN locus and unusual constitutive expression of IFN-α in bats. Proc. Natl. Acad. Sci. USA 2016, 113, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Anderson, D.E.; Zhang, Q.; Tan, C.W.; Lim, B.L.; Luko, K.; Wen, M.; Chia, W.N.; Mani, S.; Wang, L.C.; et al. Dampened NLRP3-mediated inflammation in bats and implications for a special viral reservoir host. Nat. Microbiol. 2019, 4, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Li, Y.; Shen, X.; Goh, G.; Zhu, Y.; Cui, J.; Wang, L.F.; Shi, Z.L.; Zhou, P. Dampened STING-Dependent Interferon Activation in Bats. Cell Host Microbe 2018, 23, 297–301.e294. [Google Scholar] [CrossRef] [PubMed]
- Ahn, M.; Cui, J.; Irving, A.T.; Wang, L.F. Unique Loss of the PYHIN Gene Family in Bats Amongst Mammals: Implications for Inflammasome Sensing. Sci. Rep. 2016, 6, 21722. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Baker, M.L.; Butler, J.; Munster, V. Immunological Control of Viral Infections in Bats and the Emergence of Viruses Highly Pathogenic to Humans. Front. Immunol. 2017, 8, 1098. [Google Scholar] [CrossRef] [PubMed]
- Pavlovich, S.S.; Lovett, S.P.; Koroleva, G.; Guito, J.C.; Arnold, C.E.; Nagle, E.R.; Kulcsar, K.; Lee, A.; Thibaud-Nissen, F.; Hume, A.J.; et al. The Egyptian Rousette Genome Reveals Unexpected Features of Bat Antiviral Immunity. Cell 2018, 173, 1098–1110.e18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Cowled, C.; Shi, Z.; Huang, Z.; Bishop-Lilly, K.A.; Fang, X.; Wynne, J.W.; Xiong, Z.; Baker, M.L.; Zhao, W.; et al. Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 2013, 339, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Jebb, D.; Huang, Z.; Pippel, M.; Hughes, G.M.; Lavrichenko, K.; Devanna, P.; Winkler, S.; Jermiin, L.S.; Skirmuntt, E.C.; Katzourakis, A.; et al. Six reference-quality genomes reveal evolution of bat adaptations. Nature 2020, 583, 578–584. [Google Scholar] [CrossRef]
- Huang, Z.; Whelan, C.V.; Foley, N.M.; Jebb, D.; Touzalin, F.; Petit, E.J.; Puechmaille, S.J. Longitudinal comparative transcriptomics reveals unique mechanisms underlying extended healthspan in bats. Nat. Ecol. Evol. 2019, 3, 1110–1120. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Glennon, N.B.; Jabado, O.; Lo, M.K.; Shaw, M.L. Transcriptome Profiling of the Virus-Induced Innate Immune Response in Pteropus vampyrus and Its Attenuation by Nipah Virus Interferon Antagonist Functions. J. Virol. 2015, 89, 7550–7566. [Google Scholar] [CrossRef] [PubMed]
- Wynne, J.W.; Shiell, B.J.; Marsh, G.A.; Boyd, V.; Harper, J.A.; Heesom, K.; Monaghan, P.; Zhou, P.; Payne, J.; Klein, R.; et al. Proteomics informed by transcriptomics reveals Hendra virus sensitizes bat cells to TRAIL-mediated apoptosis. Genome Biol. 2014, 15, 532. [Google Scholar] [CrossRef]
- van Tol, S.; Hage, A.; Giraldo, M.I.; Bharaj, P.; Rajsbaum, R. The TRIMendous Role of TRIMs in Virus-Host Interactions. Vaccines 2017, 5, 23. [Google Scholar] [CrossRef]
- Versteeg, G.A.; Rajsbaum, R.; Sanchez-Aparicio, M.T.; Maestre, A.M.; Valdiviezo, J.; Shi, M.; Inn, K.S.; Fernandez-Sesma, A.; Jung, J.; Garcia-Sastre, A. The E3-ligase TRIM family of proteins regulates signaling pathways triggered by innate immune pattern-recognition receptors. Immunity 2013, 38, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Versteeg, G.A.; Benke, S.; Garcia-Sastre, A.; Rajsbaum, R. InTRIMsic immunity: Positive and negative regulation of immune signaling by tripartite motif proteins. Cytokine Growth Factor Rev. 2014, 25, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Bharaj, P.; Wang, Y.E.; Dawes, B.E.; Yun, T.E.; Park, A.; Yen, B.; Basler, C.F.; Freiberg, A.N.; Lee, B.; Rajsbaum, R. The Matrix Protein of Nipah Virus Targets the E3-Ubiquitin Ligase TRIM6 to Inhibit the IKKepsilon Kinase-Mediated Type-I IFN Antiviral Response. PLoS Pathog. 2016, 12, e1005880. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Aparicio, M.T.; Feinman, L.J.; Garcia-Sastre, A.; Shaw, M.L. Paramyxovirus V Proteins Interact with the RIG-I/TRIM25 Regulatory Complex and Inhibit RIG-I Signaling. J. Virol. 2018, 92, e01960-17. [Google Scholar] [CrossRef]
- Rajsbaum, R.; Versteeg, G.A.; Schmid, S.; Maestre, A.M.; Belicha-Villanueva, A.; Martinez-Romero, C.; Patel, J.R.; Morrison, J.; Pisanelli, G.; Miorin, L.; et al. Unanchored K48-linked polyubiquitin synthesized by the E3-ubiquitin ligase TRIM6 stimulates the interferon-IKKepsilon kinase-mediated antiviral response. Immunity 2014, 40, 880–895. [Google Scholar] [CrossRef]
- Shaw, A.E.; Hughes, J.; Gu, Q.; Behdenna, A.; Singer, J.B.; Dennis, T.; Orton, R.J.; Varela, M.; Gifford, R.J.; Wilson, S.J.; et al. Fundamental properties of the mammalian innate immune system revealed by multispecies comparison of type I interferon responses. PLoS Biol. 2017, 15, e2004086. [Google Scholar] [CrossRef]
- De La Cruz-Rivera, P.C.; Kanchwala, M.; Liang, H.; Kumar, A.; Wang, L.F.; Xing, C.; Schoggins, J.W. The IFN Response in Bats Displays Distinctive IFN-Stimulated Gene Expression Kinetics with Atypical RNASEL Induction. J. Immunol. 2018, 200, 209–217. [Google Scholar] [CrossRef]
- Pavlovich, S.S.; Darling, T.; Hume, A.J.; Davey, R.A.; Feng, F.; Mühlberger, E.; Kepler, T.B. Egyptian Rousette IFN-ω Subtypes Elicit Distinct Antiviral Effects and Transcriptional Responses in Conspecific Cells. Front. Immunol. 2020, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- Rusinova, I.; Forster, S.; Yu, S.; Kannan, A.; Masse, M.; Cumming, H.; Chapman, R.; Hertzog, P.J. INTERFEROME v2.0: An updated database of annotated interferon-regulated genes. Nucleic Acids Res. 2013, 41, D1040–D1046. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Jia, M.; Song, H.; Yu, Z.; Wang, W.; Li, Q.; Zhang, L.; Zhao, W.; Cao, X. The E3 Ubiquitin Ligase TRIM40 Attenuates Antiviral Immune Responses by Targeting MDA5 and RIG-I. Cell Rep. 2017, 21, 1613–1623. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Tang, K.; Chen, D.; Hong, M.; Sun, F.; Wang, S.; Ke, Y.; Wu, T.; Sun, R.; Qian, J.; et al. Riok3 inhibits the antiviral immune response by facilitating TRIM40-mediated RIG-I and MDA5 degradation. Cell Rep. 2021, 35, 109272. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Kajihara, M.; Nao, N.; Shigeno, A.; Fujikura, D.; Hang’ombe, B.M.; Mweene, A.S.; Mutemwa, A.; Squarre, D.; Yamada, M.; et al. Characterization of a Novel Bat Adenovirus Isolated from Straw-Colored Fruit Bat (Eidolon helvum). Viruses 2017, 9, 371. [Google Scholar] [CrossRef] [PubMed]
- Yun, T.; Park, A.; Hill, T.E.; Pernet, O.; Beaty, S.M.; Juelich, T.L.; Smith, J.K.; Zhang, L.; Wang, Y.E.; Vigant, F.; et al. Efficient reverse genetics reveals genetic determinants of budding and fusogenic differences between Nipah and Hendra viruses and enables real-time monitoring of viral spread in small animal models of henipavirus infection. J. Virol. 2015, 89, 1242–1253. [Google Scholar] [CrossRef] [PubMed]
- Mohd Nor, M.N.; Gan, C.H.; Ong, B.L. Nipah virus infection of pigs in peninsular Malaysia. Rev. Sci. Tech. 2000, 19, 160–165. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Feldmann, H.; Broder, C.C. Animal challenge models of henipavirus infection and pathogenesis. Curr. Top. Microbiol. Immunol. 2012, 359, 153–177. [Google Scholar] [CrossRef]
- Simmons, N.B.; Cirranello, A.L. Bat Species of the World: A taxonomic and geographic database. 2022. Available online: https://batnames.org/ (accessed on 8 September 2023).
- Gonzalez, V.; Banerjee, A. Molecular, ecological, and behavioral drivers of the bat-virus relationship. iScience 2022, 25, 104779. [Google Scholar] [CrossRef]
- Short, K.M.; Cox, T.C. Subclassification of the RBCC/TRIM superfamily reveals a novel motif necessary for microtubule binding. J. Biol. Chem. 2006, 281, 8970–8980. [Google Scholar] [CrossRef] [PubMed]
- Reymond, A.; Meroni, G.; Fantozzi, A.; Merla, G.; Cairo, S.; Luzi, L.; Riganelli, D.; Zanaria, E.; Messali, S.; Cainarca, S.; et al. The tripartite motif family identifies cell compartments. EMBO J. 2001, 20, 2140–2151. [Google Scholar] [CrossRef] [PubMed]
- Esposito, D.; Koliopoulos, M.G.; Rittinger, K. Structural determinants of TRIM protein function. Biochem. Soc. Trans. 2017, 45, 183–191. [Google Scholar] [CrossRef]
- Koliopoulos, M.G.; Esposito, D.; Christodoulou, E.; Taylor, I.A.; Rittinger, K. Functional role of TRIM E3 ligase oligomerization and regulation of catalytic activity. EMBO J. 2016, 35, 1204–1218. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRISPR | PV TRIM40-2 F CRISPR | CACCGAGAACAGGGCTTCCGGCAGA |
| PV TRIM40-2 R CRISPR | AAACTCTGCCGGAAGCCCTGTTCTC | |
| Cloning | hTRIM40 F pCAGGS EcoRI | ATTATAGAATTCATGATCCCTTTGCAGAAGGACAACCA |
| hTRIM40 R pCAGGS SmaI | TATAATCCCGGGGAGCTTCTGAGGGGGCTGAAGAAGCA | |
| PV TRIM40 F EcoRI | ATAATAGAATTCATGTCTCCTTGAGGGAGAACAAC | |
| PV TRIM40 R SmaI | TATAATCCCGGGCTCAGGTATCAGGTCGTGGCTGTGGGGATGAGCAAAG | |
| PV TRIM40 C14.17A EcoRI F | ATAATAGAATTCATGGTCTCCTTGAGGGAGAACAACCGGGAAGAGGGCATCGCCCCCATCGCCCAGGAGCAC | |
| Pt-DDX58-FL-NotI-F | TGATTGCGGCCGCAATGACGGCCGAGGAGCGGCAGAATCTGTAC | |
| Pt-DDX58-FL-NheI-R | AATCAGCTAGCATTCATCATTTGGGCATTTCTGCAGCATCAAATGGGA | |
| Pt-IFIH1-FL-NotI-F | TGATTGCGGCCGCAATGTCGAATGAGTATTCTGCAGACAAGAGG | |
| Pt-IFIH1-FL-NheI-R | AATCAGCTAGCATTCATCAATCTTCATCACTAAACAAACAA |
| hEphrin A2 qPCR F | GACCAACGAGACCCTGTACG |
| hEphrin A2 qPCR R | CTGGGACTAGGAACCCAGGA |
| hEprhin A5 qPCR F | GGTGTTTCTGGTGCTCTGGA |
| hEphrin A5 qPCR R | AATCTGGGGTTGCTGCTGTT |
| hEphrin B1 qPCR F | CTCACACCATCCACCTCCAC |
| hEphrin B1 qPCR R | AAAAACCCTTCCGACTGCCA |
| hEphrin B2 qPCR F | TGCTGGGGTGTTTTGATGGT |
| hEphrin B2 qPCR R | ACCAGTCCTTGTCCAGGTAGA |
| hEphrin B3 qPCR F | CTCCCTTCCCTTGTGCTCTG |
| hEphrin B3 qPCR R | GCACATGGGTTCTTGGGGTA |
| Bat Ephrin A2 F qPCR | AAGTTCCAGCTCTTCACGCC |
| Bat Ephrin A2 R qPCR | CTGGTGAAGATGGGCTCAGG |
| Bat Ephrin A5 F qPCR | GTTGCACGTGGAGATGTTGA |
| Bat Ephrin A5 R qPCR | GTGGTAGTCACCCCTCTGGA |
| PV Ephrin B1 F qPCR | CCAGAGCAGGAGATTCGCTT |
| Bat Ephrin B1 R qPCR | TTGCCCGACCTTCATGACAA |
| PV Ephrin B2 F qPCR | CGCCGGACATTCTGGGAATA |
| Bat Ephrin B2 R qPCR | TGCTTCCTGTGTCTTCTCCG |
| Bat Ephrin B3 F qPCR | CTGCCCCAAACCTCCTTCTC |
| PV Ephrin B3 R qPCR | AGTAATCGTGGTGTGAGCGG |
| PV TRIM40 F | GTGTCCGGAATCTTTGACAT |
| PV TRIM40 R | GAGCTGAGTTGCTTTCTCTAA |
| PV TRIM6 F | TCACCACACGTTCCTCATGG |
| PV TRIM6 R | CATCTGATTCTTCCAGGATGTTTTC |
| PA TNFa F | TTCTGCCTGCTGCACTTCGGA |
| PA TNFa R | TCGGCTTGGGGGTTTGCTACA |
| PA IL6 F | TCACCTCTCCAAACCAAACC |
| PA IL6 R | TTTCTGCCATTTTTGGAAGG |
| EH Mx1 F | GTTACAAAAGTATGGCACAG |
| EH Mx1 R | AACAGCCGAATGTCGTTGTT |
| hTRIM40 F | CAACACACTGAAGAATGCTGG |
| hTRIM40 R | CTTCTGAGGGGGCTGAAGAAG |
| hTRIM56 F | CTCTGGCTAGTTCTCACAGGTCT |
| hTRIM56 R | CATGCTGAATGGCCAGGAACT |
| hTRIM6 F | GGGGTATGCAGCAATTCACT |
| hTRIM6 R | ACCCAATCACCCAGTATCCA |
| hTRIM25 F | CGGATGACTGCAAACAGAAA |
| hTRIM25 R | TCCTTGTCGAGGTGGTCTCT |
| hMx1 F | GGCTGTTTACCAGACTCCGACA |
| hMx1 R | CACAAAGCCTGGCAGCTCTCTA |
| hOAS1 F | GATCTCAGAAATACCCCAGCCA |
| hOAS1 R | AGCTACCTCGGAAGCACCTT |
| h18S F | GTAACCCGTTGAACCCCATT |
| h18S R | CCATCCAATCGGTAGTAGCG |
| PV TRIM40 F | GTGTCCGGAATCTTTGACAT |
| EH TRIM40 F | GTGTCCGGAATCCTTGACAT |
| PV TRIM40 R | GAGCTGAGTTGCTTTCTCTAA |
| EH TRIM40 R | GAACTGAGTTGCTTTCTCTGA |
| Pt/EH-TRIM56-RT-F | CCTGGACTGTGCTGATGACTTG |
| Pt/EH-TRIM56-RT-R | GTCATACCACCCTGCCCTGTA |
| Pt/EH-TRIM23-RT-F | GGGATGTTGGTGGAAAACACA |
| Pt/EH-TRIM23-RT-R | GGAGCAAAGCATCTCGGAGT |
| Pt/EH-TRIM25-RT-F | ACAAACCTTCACGCCCTGTA |
| Pt-TRIM25-RT-R | GAGTTTGGCTGCAAGGTAGC |
| EH-TRIM25-RT-R | GAGTTTGGCTGCAAGAGAGT |
| PV/EH GAPDH-F | ATACTTCTCATGGTTCACAC |
| PV/EH GAPDH-R | TCATTGACCTCAACTACATG |
| hGAPDH F | GCAAATTTCCATGGCACCGT |
| hGAPDH R | GCCCCACTTGATTTTGGAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Tol, S.; Hage, A.; Rajsbaum, R.; Freiberg, A.N. Pteropus vampyrus TRIM40 Is an Interferon-Stimulated Gene That Antagonizes RIG-I-like Receptors. Viruses 2023, 15, 2147. https://doi.org/10.3390/v15112147
van Tol S, Hage A, Rajsbaum R, Freiberg AN. Pteropus vampyrus TRIM40 Is an Interferon-Stimulated Gene That Antagonizes RIG-I-like Receptors. Viruses. 2023; 15(11):2147. https://doi.org/10.3390/v15112147
Chicago/Turabian Stylevan Tol, Sarah, Adam Hage, Ricardo Rajsbaum, and Alexander N. Freiberg. 2023. "Pteropus vampyrus TRIM40 Is an Interferon-Stimulated Gene That Antagonizes RIG-I-like Receptors" Viruses 15, no. 11: 2147. https://doi.org/10.3390/v15112147
APA Stylevan Tol, S., Hage, A., Rajsbaum, R., & Freiberg, A. N. (2023). Pteropus vampyrus TRIM40 Is an Interferon-Stimulated Gene That Antagonizes RIG-I-like Receptors. Viruses, 15(11), 2147. https://doi.org/10.3390/v15112147