
HPLC-Based Purification and Isolation of Potent Anti-HIV and Latency Reversing Daphnane Diterpenes from the Medicinal Plant Gnidia sericocephala (Thymelaeaceae)

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sequential Extraction of Gnidia sericocephala Roots
2.2. Microfractionation of the DCM Extract
2.3. UPLC-QTOF-MS/MS Characterization of Microfractions
2.4. Isolation and Purification of Compounds from the DCM Extract
2.5. Anti-HIV and Cytotoxicity In Vitro Screening of Compounds and Extracts of Gnidia sericocephala
2.6. Anti-HIV Screening of Compound 1 in PBMC
2.7. In Vitro Latency Reversal, Cell Viability, and T Cell Activation Assays
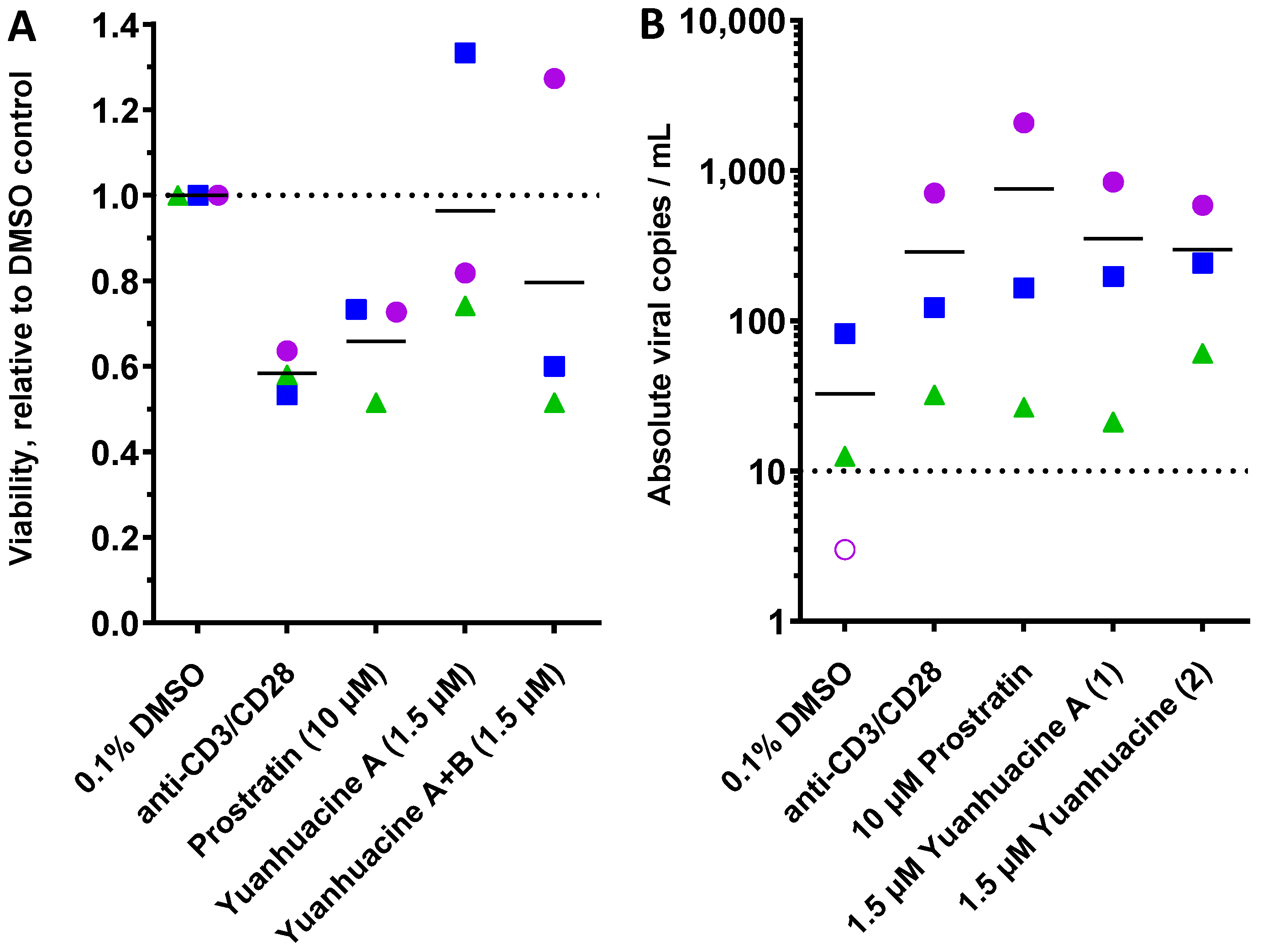
2.8. Ex Vivo Latency Reversal Assays
3. Results and Discussion
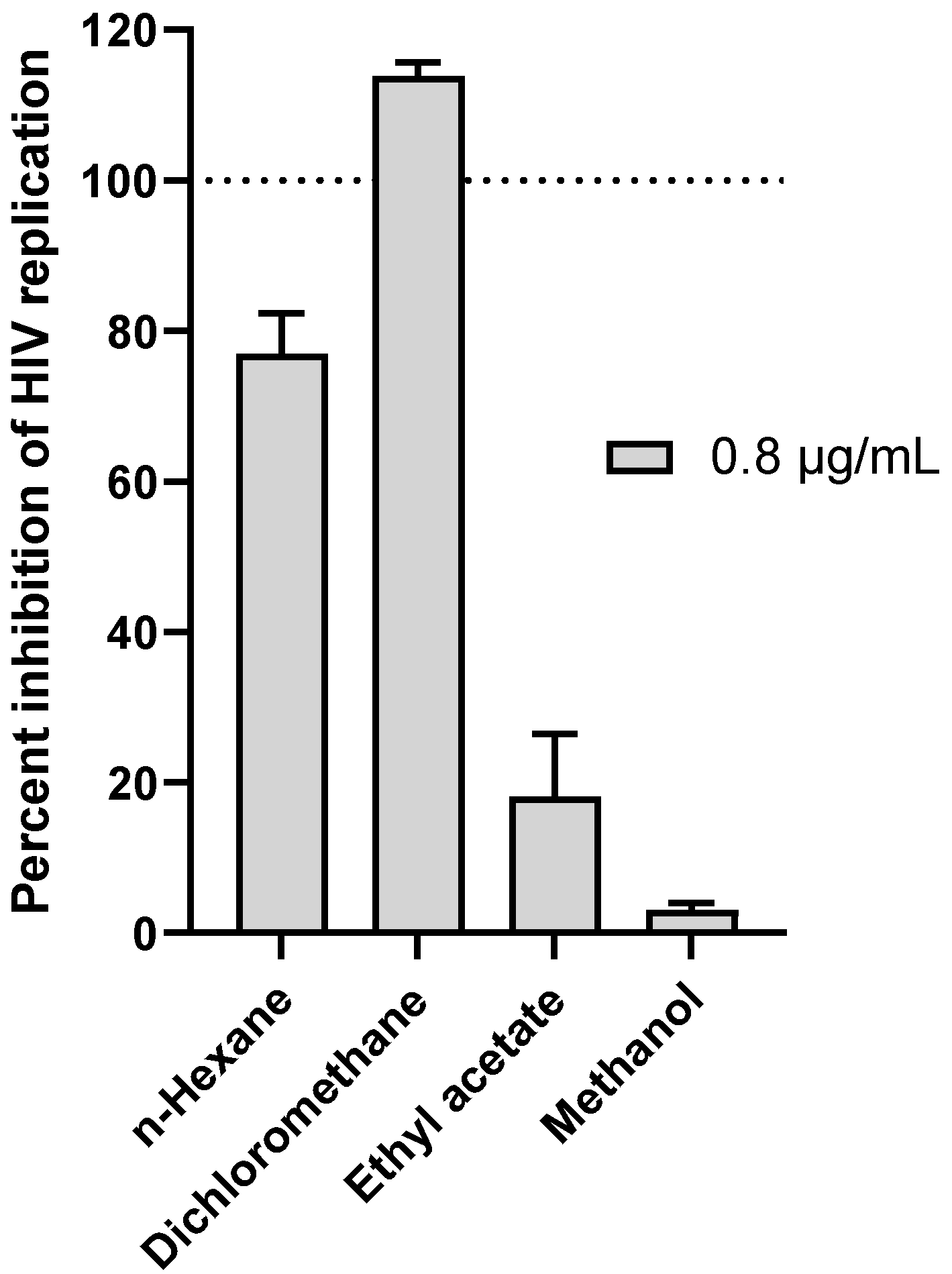
3.1. Anti-HIV Activity of G. Sericocephala Extracts
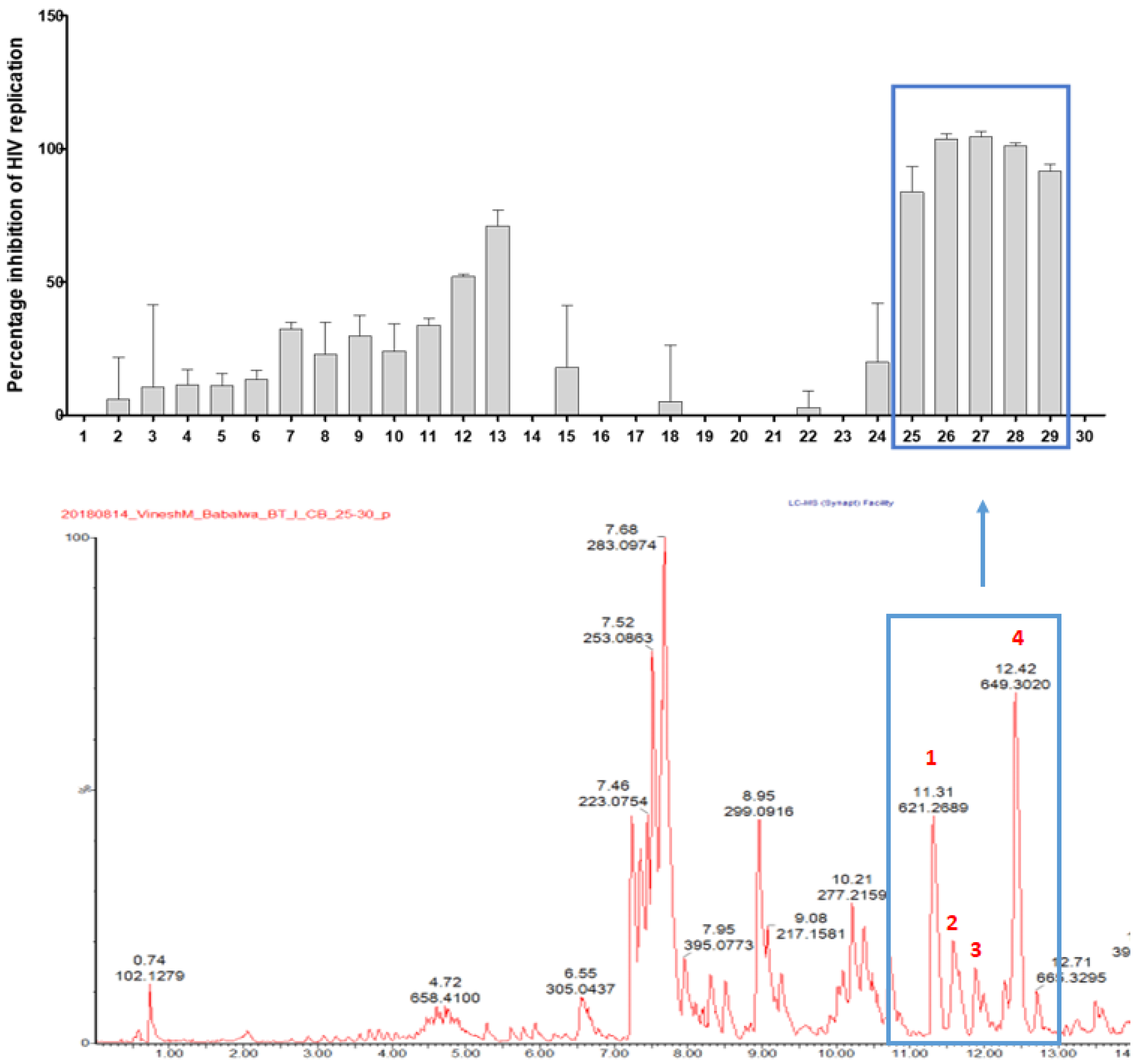
3.2. Anti-HIV Activity of HPLC Microfractions of the DCM Extract
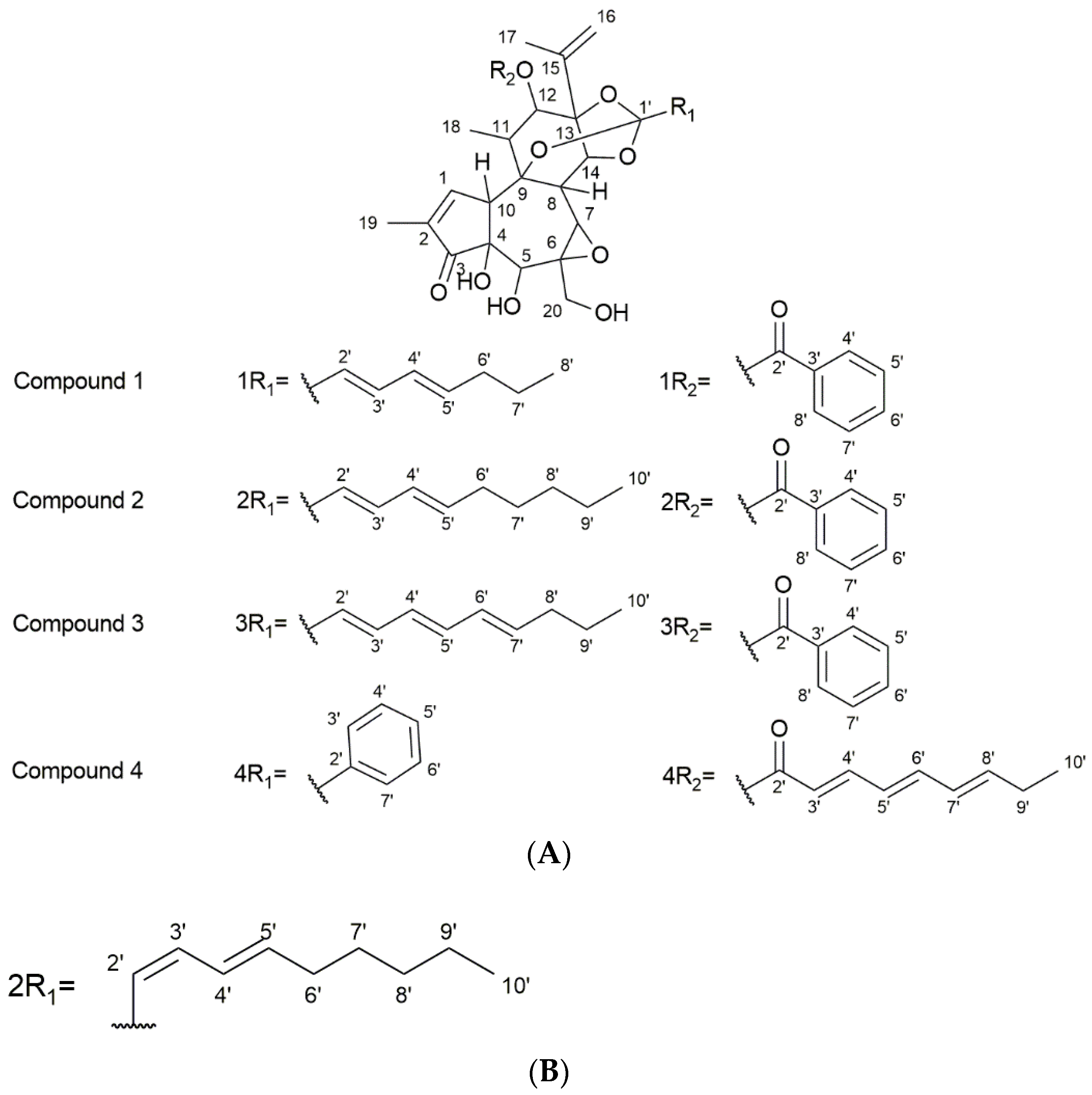
3.3. NMR and MS Analysis of Purified Compounds
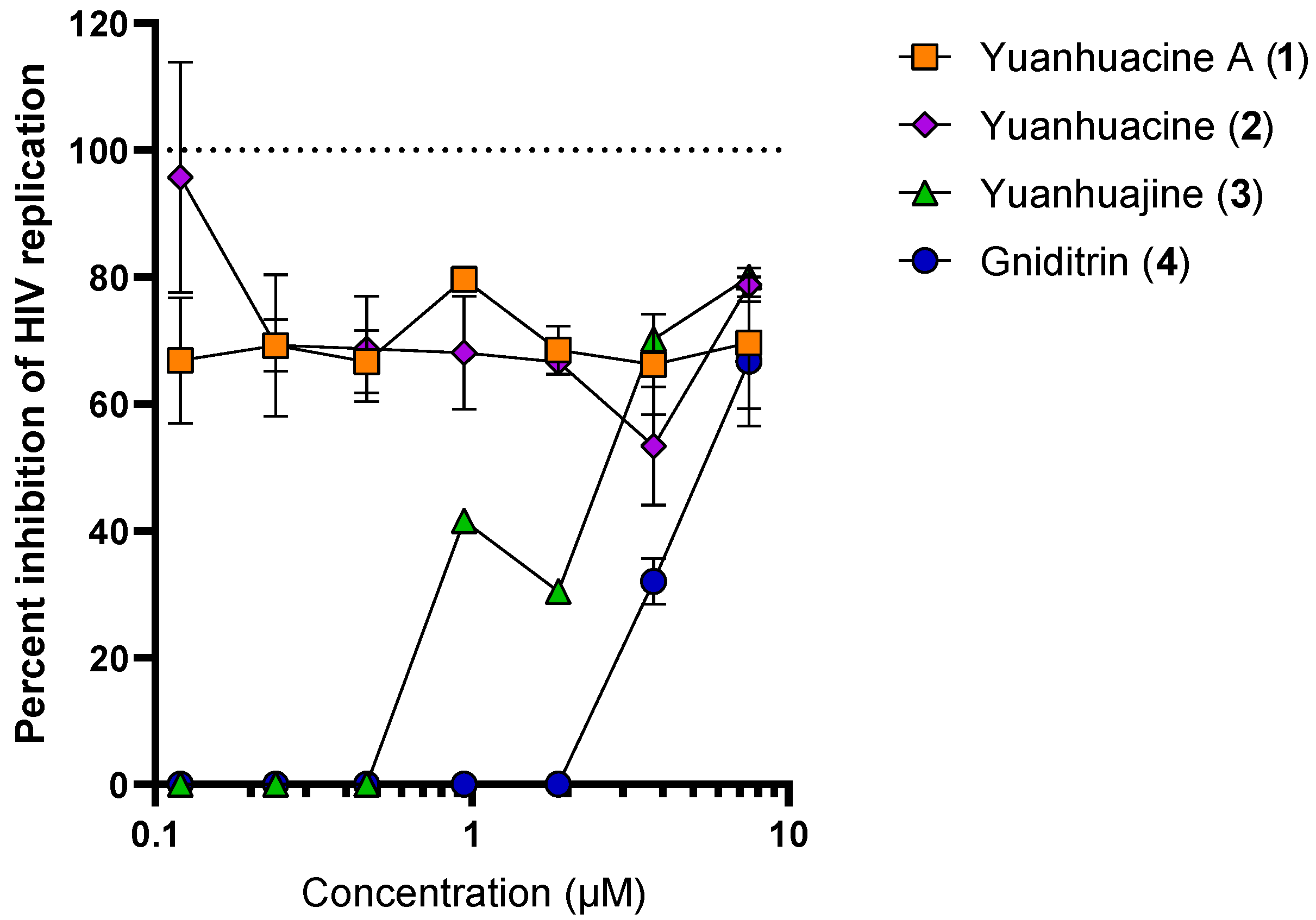
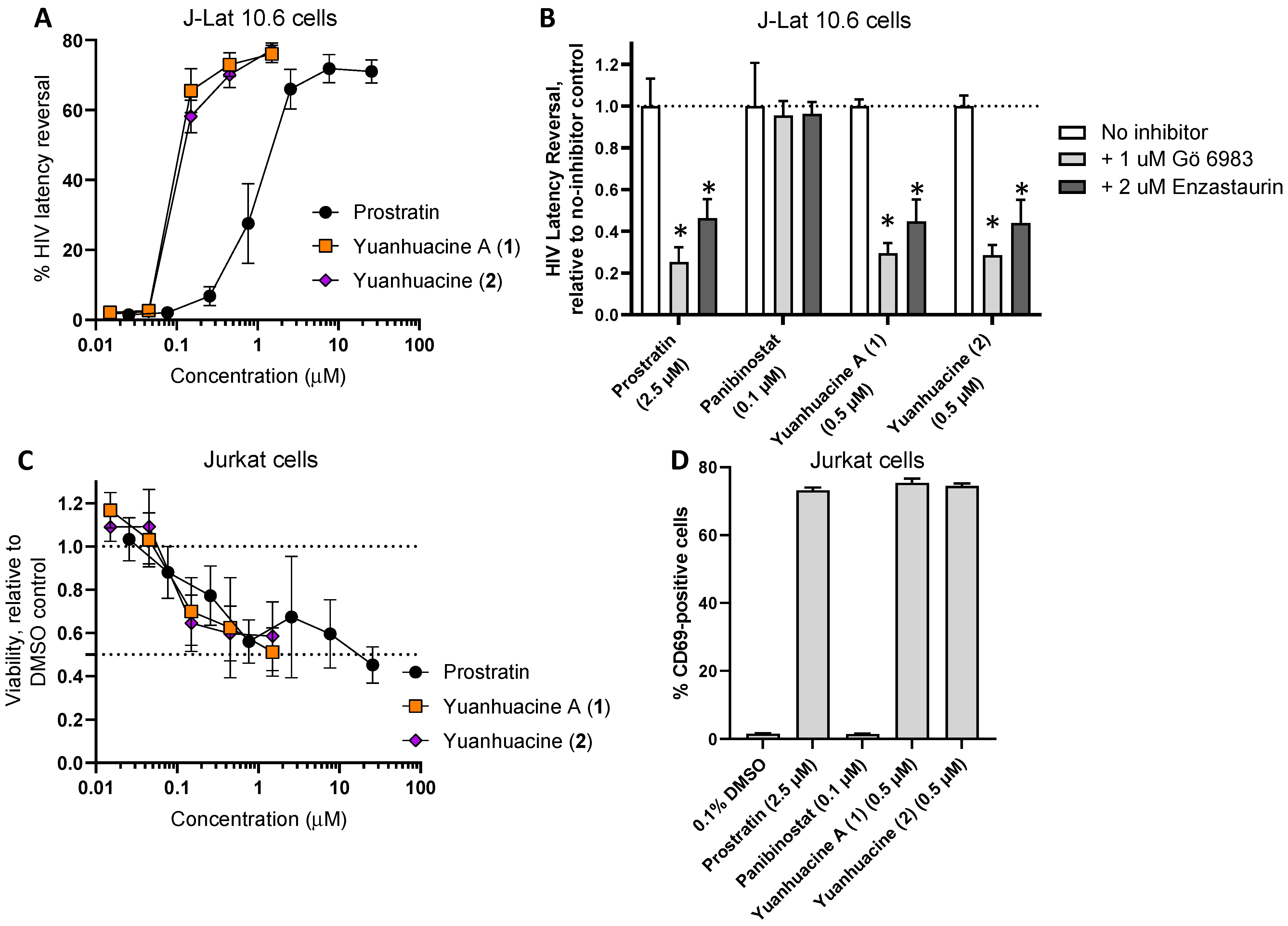
3.4. Anti-HIV Activity of Isolated Compounds
3.5. Anti-HIV Activity of Yuanhuacine A (1) in PBMC Isolates
3.6. Effects of Yuanhuacine A (1) and the Yuanhuacine-Containing Isomeric Mixture (2) on HIV Latency Reversal, Cell Viability, and T Cell Activation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNAIDS. Global AIDS update: Seizing the Moment: Tackling Entrenched Inequalities to End Epidemics. Geneva 2020. Available online: https://healtheducationresources.unesco.org/library/documents/global-aids-update-2020-seizing-moment-tackling-entrenched-inequalities-end#:~:text=to%20end%20epidemics-Global%20AIDS%20update%202020%3A%20seizing%20the%20moment%3A%20tackling,entrenched%20inequalities%20to%20end%20epidemics&text=380%20p.&text=UNAIDS%20report%20on%20the%20global,HIV%20progress%20way%20off%20course (accessed on 27 June 2022).
- Omolo, J.J.; Maharaj, V.; Naidoo, D.; Klimkait, T.; Malebo, H.M.; Mtullu, S.; Lyaruu, H.V.M.; de Koning, C.B. Bioassay-Guided Investigation of the Tanzanian Plant Pyrenacantha kaurabassana for Potential Anti-HIV-Active Compounds. J. Nat. Prod. 2012, 75, 1712–1716. [Google Scholar] [CrossRef] [PubMed]
- Ngo-Giang-Huong, N.; Huynh, T.H.K.; Dagnra, A.Y.; Toni, T.-D.; I Maiga, A.; Kania, D.; Eymard-Duvernay, S.; Peeters, M.; Soulie, C.; Peytavin, G.; et al. Prevalence of pretreatment HIV drug resistance in West African and Southeast Asian countries. J. Antimicrob. Chemother. 2018, 74, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Chimukangara, B.; Kharsany, A.B.; Lessells, R.J.; Naidoo, K.; Rhee, S.-Y.; Manasa, J.; Gräf, T.; Lewis, L.; Cawood, C.; Khanyile, D.; et al. Moderate-to-High Levels of Pretreatment HIV Drug Resistance in KwaZulu-Natal Province, South Africa. AIDS Res. Hum. Retrovir. 2019, 35, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Moyo, S.; Hunt, G.; Zuma, K.; Zungu, M.; Marinda, E.; Mabaso, M.; Kana, V.; Kalimashe, M.; Ledwaba, J.; Naidoo, I.; et al. HIV drug resistance profile in South Africa: Findings and implications from the 2017 national HIV household survey. PLoS ONE 2020, 15, e0241071. [Google Scholar] [CrossRef]
- UNAIDS. UNAIDS Data 2021. Geneva. Available online: https://www.unaids.org/en/resources/presscentre/pressreleaseandstatementarchive/2021/july/20210714_global-aids-update_en.pdf/ (accessed on 12 May 2022).
- Cillo, A.R.; Sobolewski, M.D.; Bosch, R.J.; Fyne, E.; Piatak, M.; Coffin, J.M.; Mellors, J.W. Quantification of HIV-1 latency reversal in resting CD4+ T cells from patients on suppressive antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2014, 111, 7078–7083. [Google Scholar] [CrossRef]
- Ventura, J.D. Human immunodeficiency virus 1 (HIV-1): Viral latency, the reservoir, and the cure. Yale J. Biol. Med. 2020, 93, 549–560. [Google Scholar]
- Finzi, D.; Hermankova, M.; Pierson, T.; Carruth, L.M.; Buck, C.; Chaisson, R.E.; Quinn, T.C.; Chadwick, K.; Margolick, J.; Brookmeyer, R.; et al. Identification of a Reservoir for HIV-1 in Patients on Highly Active Antiretroviral Therapy. Science 1997, 278, 1295–1300. [Google Scholar] [CrossRef]
- Hashemi, P.; Sadowski, I. Diversity of small molecule HIV-1 latency reversing agents identified in low- and high-throughput small molecule screens. Med. Res. Rev. 2019, 40, 881–908. [Google Scholar] [CrossRef]
- Archin, N.M.; Liberty, A.L.; Kashuba, A.D.; Choudhary, S.K.; Kuruc, J.D.; Crooks, A.M.; Parker, D.C.; Anderson, E.M.; Kearney, M.; Strain, M.C.; et al. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy. Nature 2012, 487, 482–485. [Google Scholar] [CrossRef]
- Abner, E.; Jordan, A. HIV “shock and kill” therapy: In need of revision. Antivir. Res. 2019, 166, 19–34. [Google Scholar] [CrossRef]
- Boucau, J.; Das, J.; Joshi, N.; Le Gall, S. Latency reversal agents modulate HIV antigen processing and presentation to CD8 T cells. PLOS Pathog. 2020, 16, e1008442. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.J.; Ntie-Kang, F.; Tietjen, I. Natural product-derived compounds in HIV suppression, remission, and eradication strategies. Antivir. Res. 2018, 158, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.N.; McMonagle, E.L.; Hosie, M.J.; Willett, B.J. Prostratin exhibits both replication enhancing and inhibiting effects on FIV infection of feline CD4+ T-cells. Virus Res. 2013, 171, 121–128. [Google Scholar] [CrossRef][Green Version]
- Mehla, R.; Bivalkar-Mehla, S.; Zhang, R.; Handy, I.; Albrecht, H.; Giri, S.; Nagarkatti, P.; Nagarkatti, M.; Chauhan, A. Bryostatin Modulates Latent HIV-1 Infection via PKC and AMPK Signaling but Inhibits Acute Infection in a Receptor Independent Manner. PLoS ONE 2010, 5, e11160. [Google Scholar] [CrossRef]
- Vidal, V.; Potterat, O.; Louvel, S.; Hamy, F.; Mojarrab, M.; Sanglier, J.-J.; Klimkait, T.; Hamburger, M. Library-Based Discovery and Characterization of Daphnane Diterpenes as Potent and Selective HIV Inhibitors in Daphne gnidium. J. Nat. Prod. 2011, 75, 414–419. [Google Scholar] [CrossRef]
- Pierens, G.K. 1H and 13C NMR scaling factors for the calculation of chemical shifts in commonly used solvents using density functional theory. J. Comput. Chem. 2014, 35, 1388–1394. [Google Scholar] [CrossRef]
- Lanier, E.R.; Ptak, R.G.; Lampert, B.M.; Keilholz, L.; Hartman, T.; Buckheit, R.W.; Mankowski, M.K.; Osterling, M.C.; Almond, M.R.; Painter, G.R. Development of Hexadecyloxypropyl Tenofovir (CMX157) for Treatment of Infection Caused by Wild-Type and Nucleoside/Nucleotide-Resistant HIV. Antimicrob. Agents Chemother. 2010, 54, 2901–2909. [Google Scholar] [CrossRef]
- Ptak, R.G.; Gallay, P.A.; Jochmans, D.; Halestrap, A.P.; Ruegg, U.T.; Pallansch, L.A.; Bobardt, M.D.; de Béthune, M.-P.; Neyts, J.; De Clercq, E.; et al. Inhibition of Human Immunodeficiency Virus Type 1 Replication in Human Cells by Debio-025, a Novel Cyclophilin Binding Agent. Antimicrob. Agents Chemother. 2008, 52, 1302–1317. [Google Scholar] [CrossRef]
- Jordan, A.; Bisgrove, D.; Verdin, E. J Lat 10-6-HIV-repsoducibly establishes a latent infection after acute infection of T cells in vitro-cdg188. EMBO J. 2003, 22, 1868–1877. [Google Scholar] [CrossRef]
- Schonhofer, C.; Yi, J.; Sciorillo, A.; Andrae-Marobela, K.; Cochrane, A.; Harris, M.; Brumme, Z.L.; Brockman, M.A.; Mounzer, K.; Hart, C.; et al. Flavonoid-based inhibition of cyclin-dependent kinase 9 without concomitant inhibition of histone deacetylases durably reinforces HIV latency. Biochem. Pharmacol. 2021, 186, 114462. [Google Scholar] [CrossRef]
- Yuan, Z.; Wang, N.; Kang, G.; Niu, W.; Li, Q.; Guo, J. Controlling Multicycle Replication of Live-Attenuated HIV-1 Using an Unnatural Genetic Switch. ACS Synth. Biol. 2017, 6, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Adachi, A.; E Gendelman, H.; Koenig, S.; Folks, T.; Willey, R.; Rabson, A.; A Martin, M. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol. 1986, 59, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.-L.; Yao, G.-D.; Song, S.-J. Daphnane-type diterpenes from genus Daphne and their anti-tumor activity. Chin. Herb. Med. 2021, 13, 145–156. [Google Scholar] [CrossRef]
- De Mieri, M.; Du, K.; Neuburger, M.; Saxena, P.; Zietsman, P.C.; Hering, S.; van der Westhuizen, J.H.; Hamburger, M. hERG Channel Inhibitory Daphnane Diterpenoid Orthoesters and Polycephalones A and B with Unprecedented Skeletons from Gnidia polycephala. J. Nat. Prod. 2015, 78, 1697–1707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, X.; Zhang, F.; Yang, P.; Gao, X.; Song, Q. Preparation of yuanhuacine and relative daphne diterpene esters from Daphne genkwa and structure—Activity relationship of potent inhibitory activity against DNA topoisomerase I. Bioorganic Med. Chem. 2006, 14, 3888–3895. [Google Scholar] [CrossRef] [PubMed]
- Görick, C.; Melzig, M.F. Gniditrin is the main diterpenoid constituent in the bark of Daphne mezereum L. Die Pharm. 2013, 68, 640–642. [Google Scholar] [CrossRef]
- Emert, J.; Breslow, R. Gnididin, gniditrin, and gnidicin, novel potent antileukemic diterpenoid esters from Gnidia lamprantha. J. Am. Chem. Soc. 1975, 97, 672–673. [Google Scholar]
- Jin, Y.-X.; Shi, L.-L.; Zhang, D.-P.; Wei, H.-Y.; Si, Y.; Ma, G.-X.; Zhang, J. A Review on Daphnane-Type Diterpenoids and Their Bioactive Studies. Molecules 2019, 24, 1842. [Google Scholar] [CrossRef]
- Liao, S.-G.; Chen, H.-D.; Yue, J.-M. Plant Orthoesters. Chem. Rev. 2009, 109, 1092–1140. [Google Scholar] [CrossRef]
- Wang, M.; Tietjen, I.; Chen, M.; Williams, D.E.; Daoust, J.; Brockman, M.A.; Andersen, R.J. Sesterterpenoids Isolated from the Sponge Phorbas sp. Activate Latent HIV-1 Provirus Expression. J. Org. Chem. 2016, 81, 11324–11334. [Google Scholar] [CrossRef]
- Shawon, J.; Akter, Z.; Hossen, M.M.; Akter, Y.; Sayeed, A.; Junaid, M.; Afrose, S.S.; Khan, M.A. Current Landscape of Natural Products against Coronaviruses: Perspectives in COVID-19 Treatment and Anti-viral Mechanism. Curr. Pharm. Des. 2020, 26, 5241–5260. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.; Huang, L.; Zhu, L.; Ferrari, G.; Chan, C.; Li, W.; Lee, K.-H.; Chen, C.-H. Gnidimacrin, a Potent Anti-HIV Diterpene, Can Eliminate Latent HIV-1 Ex Vivo by Activation of Protein Kinase C β. J. Med. Chem. 2015, 58, 8638–8646. [Google Scholar] [CrossRef] [PubMed]
- Otsuki, K.; Li, W.; Miura, K.; Asada, Y.; Huang, L.; Chen, C.-H.; Lee, K.-H.; Koike, K. Isolation, Structural Elucidation, and Anti-HIV Activity of Daphnane Diterpenoids from Daphne odora. J. Nat. Prod. 2020, 83, 3270–3277. [Google Scholar] [CrossRef]
- Huang, L.; Ho, P.; Yu, J.; Zhu, L.; Lee, K.-H.; Chen, C.-H. Picomolar Dichotomous Activity of Gnidimacrin Against HIV-1. PLoS ONE 2011, 6, e26677. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Compound | ||||||
|---|---|---|---|---|---|---|---|
| Yuanhuacine A (1) | Yuanhuacine * (2) | Yuanhuajine (3) | Gniditrin (4) | ||||
| δC | δH (J) | δC | δH (J) | δC | δH (J) | δH (J) | |
| 1 | 160.47, CH | 7.48, t (8.06) | 160.50, CH | 7.62, s | 160.05, CH | 7.39, d (4.5) | 7.59, s |
| 2 | 136.96, C | 137.04, C | - | 136.98, C | |||
| 3 | 209.50, C | 209.54, C | - | 209.50, C | |||
| 4 | 71.98, C | 60.58, C | - | 78.37, C | |||
| 5 | 72.17, CH | 4.25, s | 64.07, CH | 3.66, s | 72.13, CH | 4.27, s | 4.27, s |
| 6 | 60.57, C | - | 72.15, C | - | 60.54, C | ||
| 7 | 64.73, CH | 3.84, d (13.0) | 72.00, CH | 4.27, s | 64.41, CH | 3.98, br d (7.6, 4.7) | 3.48, s |
| 8 | 34.80, CH | 3.65, d (2.6) | 35.7, CH | 3.66, s | 35.56, CH | 3.81, dd (12.4, 12.7) | 3.80, dt (5.7, 12.5) |
| 9 | 80.50, C | 78.40, C | - | 72.22, C | |||
| 10 | 47.50, CH | 3.87, quint (5.2) | 47.3, CH | 3.88, d (19.5) | 47.51, CH | 3.93, m | 3.97, m |
| 11 | 44.15, CH | 2.58, q (7.3) | 44.2, CH | 2.57, m | 44.21, CH | 2.56, m | 2.51, m |
| 12 | 78.24, CH | 5.24, s | 78.9, CH | 5.24, s | 78.64, CH | 5.11, t (7.9) | 5.10, q (4.4, 8.5) |
| 13 | 83.92, C | 83.92, C | - | 84.27, C | |||
| 14 | 78.96, CH | 4.92, d (1.2) | 81.2, CH | 4.92, s | 80.86, CH | 4.93, d (2.5) | 4.92, s |
| 15 | 143.00, C | 143.06, C | - | 142.9, C | |||
| 16 | 113.67, CH2 | 5.02, m 5.03, s | 114.62, CH2 | 5.04, t (8.4) | 113.83, CH2 | 5.02, d (4.9) | 5.02, d (4.12) |
| 17 | 18.80, CH3 | 1.89, br s | 18.79, CH3 | 1.90, s | 18.37, CH3 | 1.88, s | 1.88, s |
| 18 | 18.37, CH3 | 1.41, d (7.69) | 18.37, CH3 | 1.42, m | 18.97, CH3 | 1.41, t (3.9) | 1.41, t (3.9) |
| 19 | 9.90, CH3 | 1.79, dd (1.27) | 9.90, CH3 | 1.80, s | 9.96, CH3 | 1.75, s | 1.75, s |
| 20 | 64.07, CH2 | 3.97, d (12.7), 3.67 s | 64.72, CH2 | 3.97, d (19.5) | 63.8, CH2 | 3.64, dd (7.6, 4.7) | 3.63, dt (2.7, 3.1) |
| 1′ | 117.10, C | 117.1, C | - | 119.3, C | |||
| 2′ | 122.29, CH | 5.70, d (15.4) | 122.41, CH | 5.70, d (15.44) | 135.3, CH | 5.81, d (15.9) | |
| 3′ | 135.24, CH | 6.72, dd (10.87, 15.33) | 135.4, CH | 6.73, dd (11.07, 15.31) | 128.2, CH | 7.39, m | 7.72, dd (4.9, 1.96) |
| 4′ | 128.64, CH | 6.09, dd (10.81, 15.3) | 128.7, CH | 6.08 (11.09, 15.03) | 127.2, CH | 6.20, m | 7.39, d (6.2) |
| 5′ | 139.20, CH | 5.89, dt (15.3, 14.5) | 139.6, CH | 5.89, m (6.9,15.03) | 129.9, CH | 6.6, m | 7.39, d (6.22) |
| 6′ | 35.81, CH2 | 2.11, q (7.3) | 32.8, CH2 | 2.12, d (7.35) | 127.2, CH | 6.16, m | 7.39, d (6.22) |
| 7′ | 22.24, CH2 | 1.44, sext (7.3) | 28.9, CH2 | 1.41, dd (4.1, 6.7) | 141.5, CH | 5.97, m | 7.59, s |
| 8′ | 13.65, CH3 | 0.93, t (7.3) | 31.4, CH2 | 1.27, s | 36.1, CH2 | 2.1, q (8.7, 15.3) | |
| 9′ | 22.68, CH2 | 1.31, br s | 22.7, CH2 | 1.44, m | |||
| 10′ | 14.2, CH3 | 0.91, t (6.7) | 13.8, CH3 | 0.92, t (7.4) | |||
| 1″ | 165.54, C | 165.8, C | - | 165.9, C | |||
| 2″ | 129.48, C | 129.8, C | - | 135.6, C | 5.75, m | ||
| 3″ | 7.39, d (6.22) | ||||||
| 3″-7″ | 129.71, CH | 7.92, dd (8.2, 8.4) | 129.5, CH | 7.92, d (7.6) | 126.3, CH | 7.72, m | |
| 4″-6″ | 128.76, CH | 7.48, t (8.1, 15.2) | 128.8, CH | 7.49, t (7.9) | 128.2, CH | 7.39, m | |
| 4″ | 6.17, m | ||||||
| 5″ | 133.45, CH | 7.61, m | 133.5, CH | 7.62, s | 129.9, CH | 7.59, m | 6.56, m |
| 6″ | 5.95, m | ||||||
| 7″ | 5.10, q | ||||||
| 8″ | 2.1, q (8.7, 15.3) | ||||||
| 9″ | 1.44, m | ||||||
| 10″ | 0.92, t (7.4) | ||||||
| Compound | Antiviral Activity (µM) | Cytotoxicity (µM) | Selectivity Index | |
|---|---|---|---|---|
| EC50 | EC90 | CC50 | CC50/IC50 | |
| Yuanhuacine (1) | 0.03 | 0.09 | >15 | >500 |
| AZT | 0.007 | 0.05 | >1 | >142 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tembeni, B.; Sciorillo, A.; Invernizzi, L.; Klimkait, T.; Urda, L.; Moyo, P.; Naidoo-Maharaj, D.; Levitties, N.; Gyampoh, K.; Zu, G.; et al. HPLC-Based Purification and Isolation of Potent Anti-HIV and Latency Reversing Daphnane Diterpenes from the Medicinal Plant Gnidia sericocephala (Thymelaeaceae). Viruses 2022, 14, 1437. https://doi.org/10.3390/v14071437
Tembeni B, Sciorillo A, Invernizzi L, Klimkait T, Urda L, Moyo P, Naidoo-Maharaj D, Levitties N, Gyampoh K, Zu G, et al. HPLC-Based Purification and Isolation of Potent Anti-HIV and Latency Reversing Daphnane Diterpenes from the Medicinal Plant Gnidia sericocephala (Thymelaeaceae). Viruses. 2022; 14(7):1437. https://doi.org/10.3390/v14071437
Chicago/Turabian StyleTembeni, Babalwa, Amanda Sciorillo, Luke Invernizzi, Thomas Klimkait, Lorena Urda, Phanankosi Moyo, Dashnie Naidoo-Maharaj, Nathan Levitties, Kwasi Gyampoh, Guorui Zu, and et al. 2022. "HPLC-Based Purification and Isolation of Potent Anti-HIV and Latency Reversing Daphnane Diterpenes from the Medicinal Plant Gnidia sericocephala (Thymelaeaceae)" Viruses 14, no. 7: 1437. https://doi.org/10.3390/v14071437
APA StyleTembeni, B., Sciorillo, A., Invernizzi, L., Klimkait, T., Urda, L., Moyo, P., Naidoo-Maharaj, D., Levitties, N., Gyampoh, K., Zu, G., Yuan, Z., Mounzer, K., Nkabinde, S., Nkabinde, M., Gqaleni, N., Tietjen, I., Montaner, L. J., & Maharaj, V. (2022). HPLC-Based Purification and Isolation of Potent Anti-HIV and Latency Reversing Daphnane Diterpenes from the Medicinal Plant Gnidia sericocephala (Thymelaeaceae). Viruses, 14(7), 1437. https://doi.org/10.3390/v14071437