
Canid herpesvirus 1 Preferentially Infects Polarized Madin-Darby Canine Kidney Cells from the Basolateral Surface



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
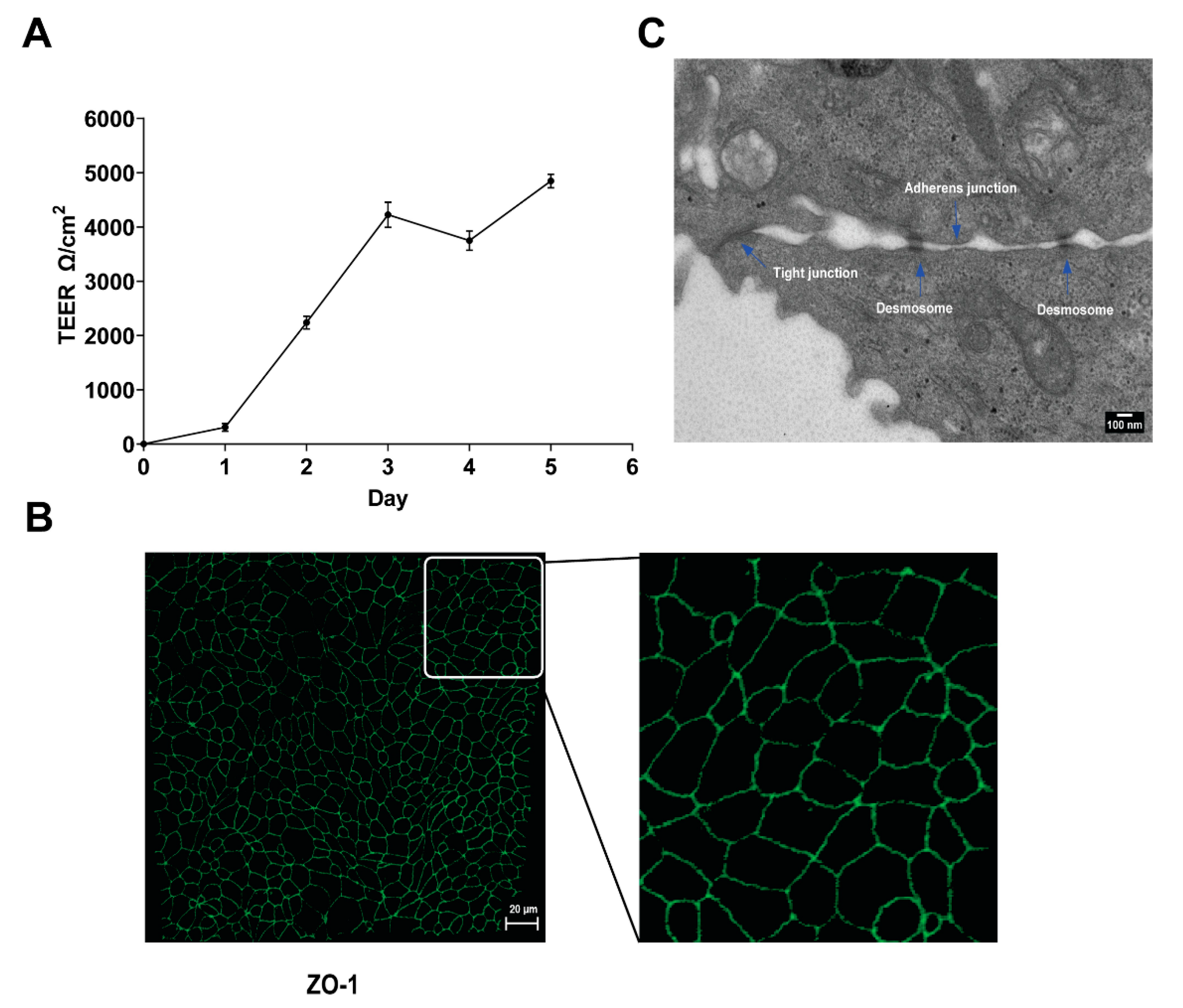
2.2. Polarized Cell Cultures
2.3. Confocal Microscopy
2.4. Scanning and Transmission Electron Microscopy
2.5. Viral Yield Assays
3. Results
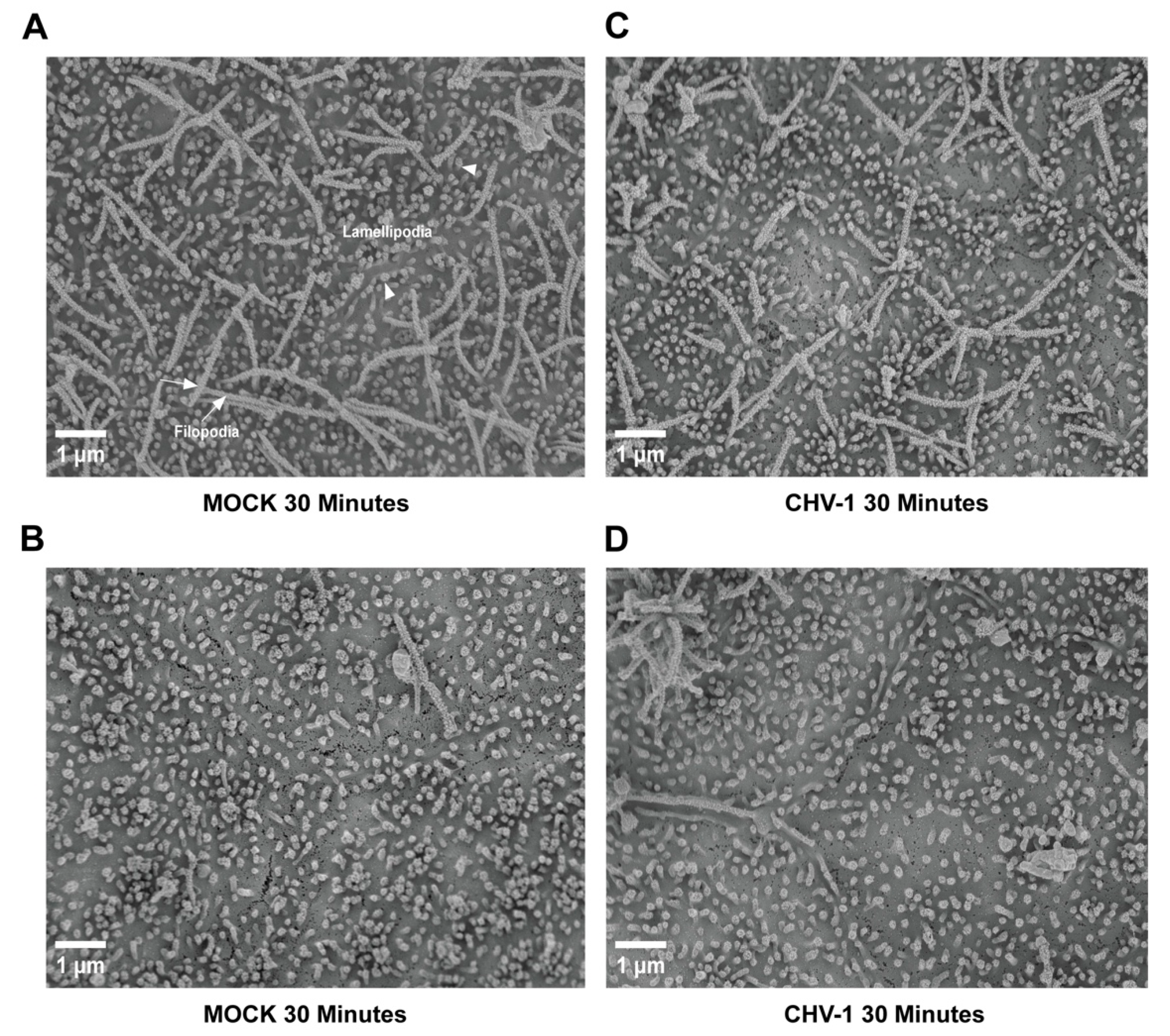
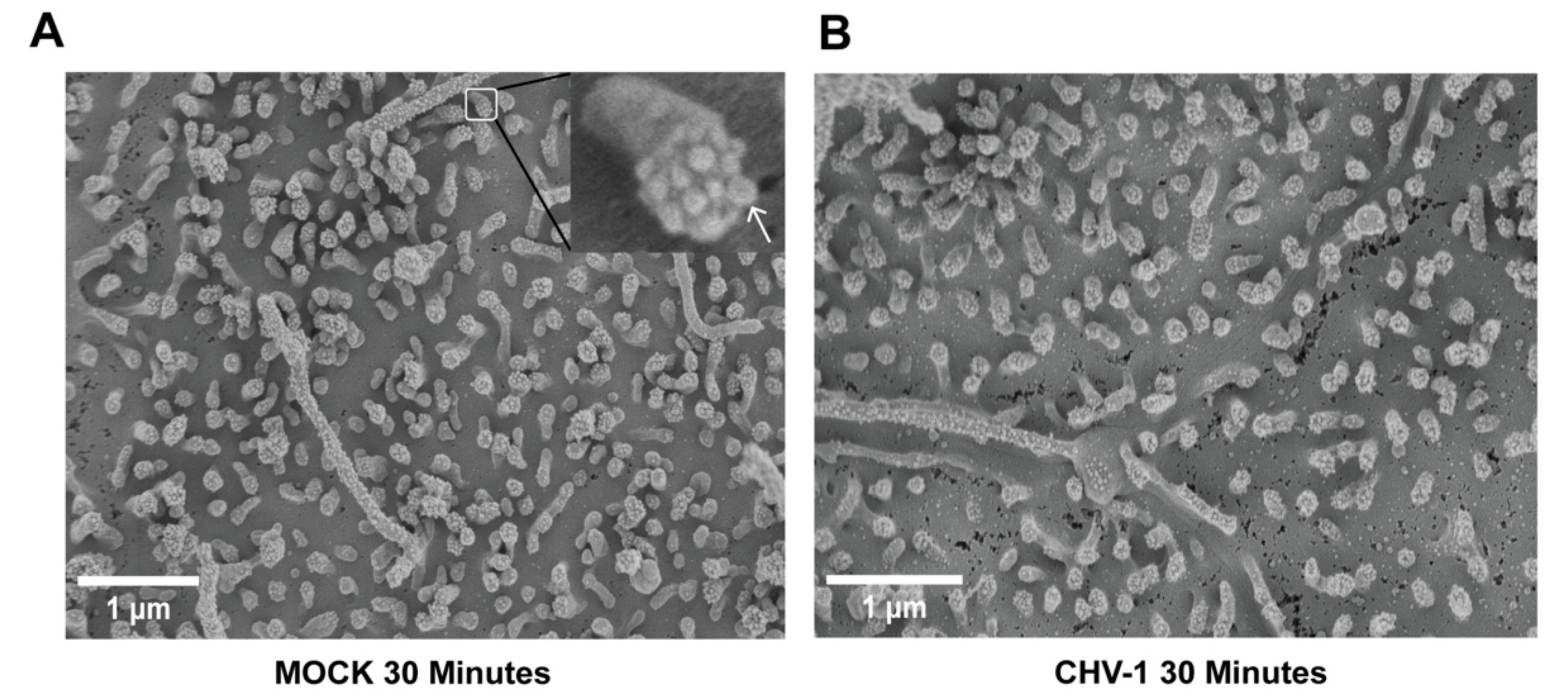
3.1. Impact of CHV-1 on Membrane Protrusions of Polarized MDCK Cells
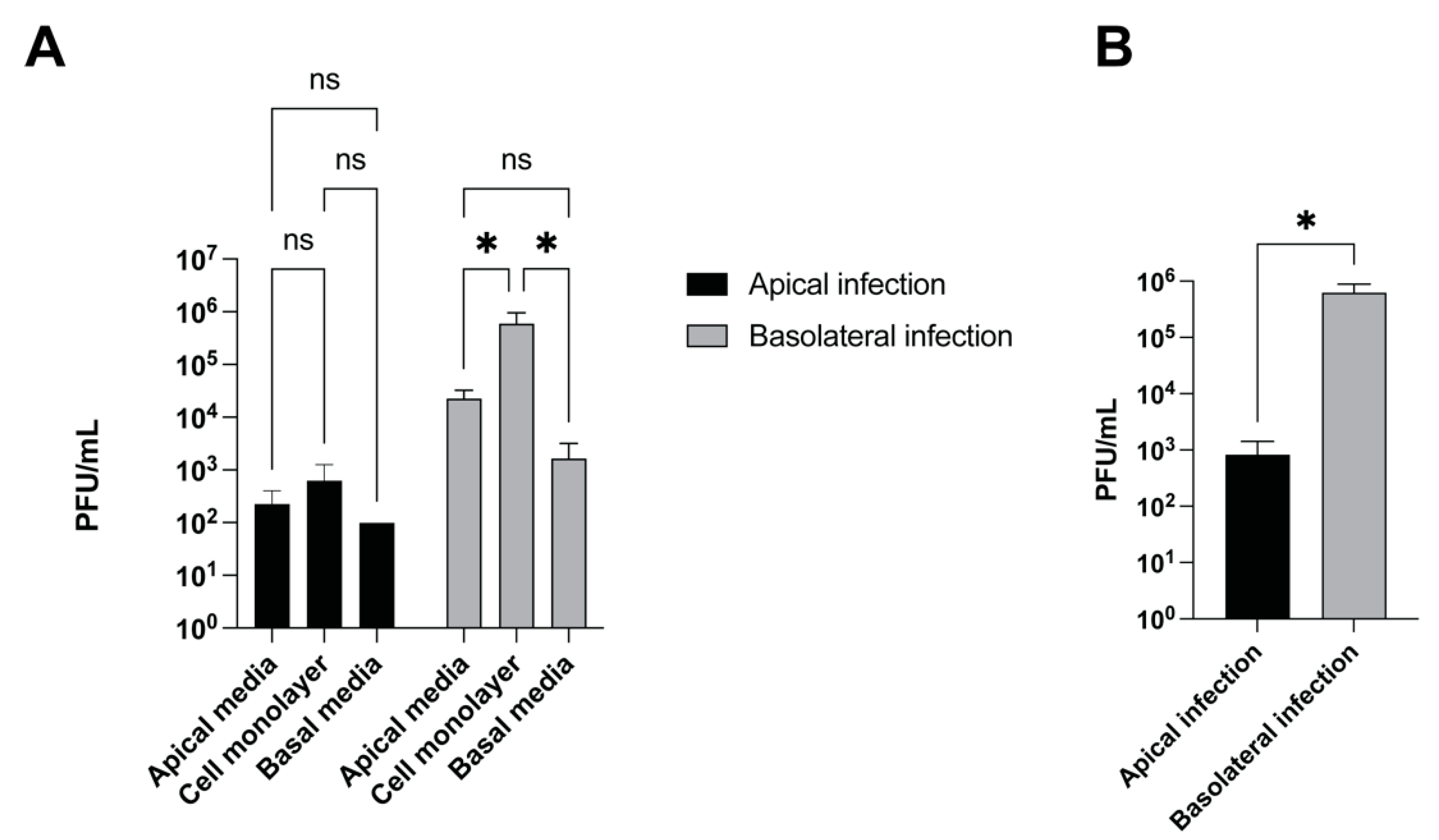
3.2. CHV-1 Infects the Basolateral Side of Polarized MDCK Cells More Efficiently than the Apical Side
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ledbetter, E.C.; Kim, S.G.; Dubovi, E.J.; Bicalho, R.C. Experimental reactivation of latent canine herpesvirus-1 and induction of recurrent ocular disease in adult dogs. Vet. Microbiol. 2009, 138, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, M.; Ishii, Y.; Takiguchi, M.; Takada, A.; Yasuda, J.; Hashimoto, A.; Okazaki, K.; Kida, H. Detection of canine herpesvirus DNA in the ganglionic neurons and the lymph node lymphocytes of latently infected dogs. J. Vet. Med. Sci. Jpn. Soc. Vet. Sci. 1999, 61, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, K.; Ohara, K.; Matsumoto, Y.; Otsuka, H. Attachment and penetration of canine herpesvirus 1 in non-permissive cells. J. Vet. Med. Sci. Jpn. Soc. Vet. Sci. 2000, 62, 965–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laquerre, S.; Argnani, R.; Anderson, D.B.; Zucchini, S.; Manservigi, R.; Glorioso, J.C. Heparan sulfate proteoglycan binding by herpes simplex virus type 1 glycoproteins B and C, which differ in their contributions to virus attachment, penetration, and cell-to-cell spread. J. Virol. 1998, 72, 6119–6130. [Google Scholar] [CrossRef] [Green Version]
- Tal-Singer, R.; Peng, C.; Ponce De Leon, M.; Abrams, W.R.; Banfield, B.W.; Tufaro, F.; Cohen, G.H.; Eisenberg, R.J. Interaction of herpes simplex virus glycoprotein gC with mammalian cell surface molecules. J. Virol. 1995, 69, 4471–4483. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Negussie, H.; Qiu, Y.; Reddy, V.R.; Mateusen, B.; Nauwynck, H.J. Early events of canine herpesvirus 1 infections in canine respiratory and genital mucosae by the use of ex vivo models. Res. Vet. Sci. 2016, 105, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Eisa, M.; Loucif, H.; van Grevenynghe, J.; Pearson, A. Entry of the Varicellovirus Canid herpesvirus 1 into Madin-Darby canine kidney epithelial cells is pH-independent and occurs via a macropinocytosis-like mechanism but without increase in fluid uptake. Cell. Microbiol. 2021, 23, e13398. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, K.V.; Suarez, N.M.; Wilkie, G.S.; McDonald, M.; Graham, E.M.; Davison, A.J. Genome Sequence of Canine Herpesvirus. PLoS ONE 2016, 11, e0156015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeil, E.; Capaldo, C.T.; Macara, I.G. Zonula occludens-1 function in the assembly of tight junctions in Madin-Darby canine kidney epithelial cells. Mol. Biol. Cell 2006, 17, 1922–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marozin, S.; Prank, U.; Sodeik, B. Herpes simplex virus type 1 infection of polarized epithelial cells requires microtubules and access to receptors present at cell-cell contact sites. J. Gen. Virol. 2004, 85, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Whittaker, G.R. Influenza entry pathways in polarized MDCK cells. Biochem. Biophys. Res. Commun. 2014, 450, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Sambuy, Y.; Rodriguez-Boulan, E. Isolation and characterization of the apical surface of polarized Madin-Darby canine kidney epithelial cells. Proc. Natl. Acad. Sci. USA 1988, 85, 1529–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galen, B.; Cheshenko, N.; Tuyama, A.; Ramratnam, B.; Herold, B.C. Access to nectin favors herpes simplex virus infection at the apical surface of polarized human epithelial cells. J. Virol. 2006, 80, 12209–12218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, M.; Wang, F.; Li, W.; Zhang, X.; Zhang, Z.; Zhang, X.E.; Cui, Z. Ebola Virus Uptake into Polarized Cells from the Apical Surface. Viruses 2019, 11, 1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, A.; Fu, Y.; Meens, J.; Yang, W.; Meng, F.; Herrler, G.; Becher, P. Infection of polarized bovine respiratory epithelial cells by bovine viral diarrhea virus (BVDV). Virulence 2021, 12, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Kommnick, C.; Lepper, A.; Hensel, M. Correlative light and scanning electron microscopy (CLSEM) for analysis of bacterial infection of polarized epithelial cells. Sci. Rep. 2019, 9, 17079. [Google Scholar] [CrossRef] [PubMed]
- Harman, R.M.; Bussche, L.; Ledbetter, E.C.; Van de Walle, G.R. Establishment and characterization of an air-liquid canine corneal organ culture model to study acute herpes keratitis. J. Virol. 2014, 88, 13669–13677. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eisa, M.; Micky, S.; Pearson, A. Canid herpesvirus 1 Preferentially Infects Polarized Madin-Darby Canine Kidney Cells from the Basolateral Surface. Viruses 2022, 14, 1291. https://doi.org/10.3390/v14061291
Eisa M, Micky S, Pearson A. Canid herpesvirus 1 Preferentially Infects Polarized Madin-Darby Canine Kidney Cells from the Basolateral Surface. Viruses. 2022; 14(6):1291. https://doi.org/10.3390/v14061291
Chicago/Turabian StyleEisa, Mohamed, Samar Micky, and Angela Pearson. 2022. "Canid herpesvirus 1 Preferentially Infects Polarized Madin-Darby Canine Kidney Cells from the Basolateral Surface" Viruses 14, no. 6: 1291. https://doi.org/10.3390/v14061291
APA StyleEisa, M., Micky, S., & Pearson, A. (2022). Canid herpesvirus 1 Preferentially Infects Polarized Madin-Darby Canine Kidney Cells from the Basolateral Surface. Viruses, 14(6), 1291. https://doi.org/10.3390/v14061291