
Torque Teno Virus Primary Infection Kinetics in Early Childhood

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects and Samples
2.2. Ethics
2.3. DNA Extraction
2.4. TTV Quantification
2.5. TTV Genotyping
2.5.1. Sample Selection for TTV Genotyping
2.5.2. Rolling Circle Amplification
2.5.3. Multiplex PCR for TTV Typing
2.5.4. Cloning and Sequencing
2.5.5. TTV Control Plasmids for TTV Genotyping
3. Results
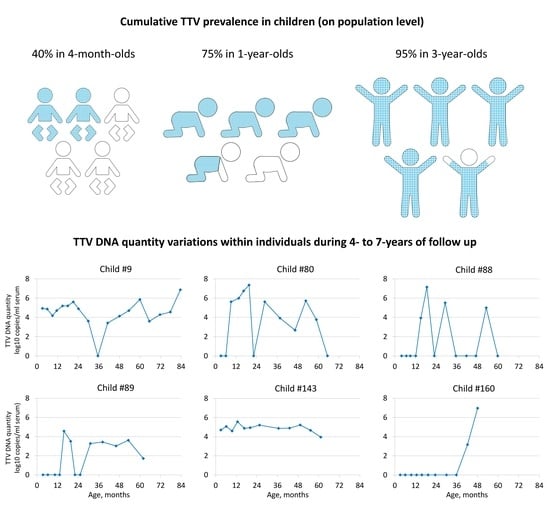
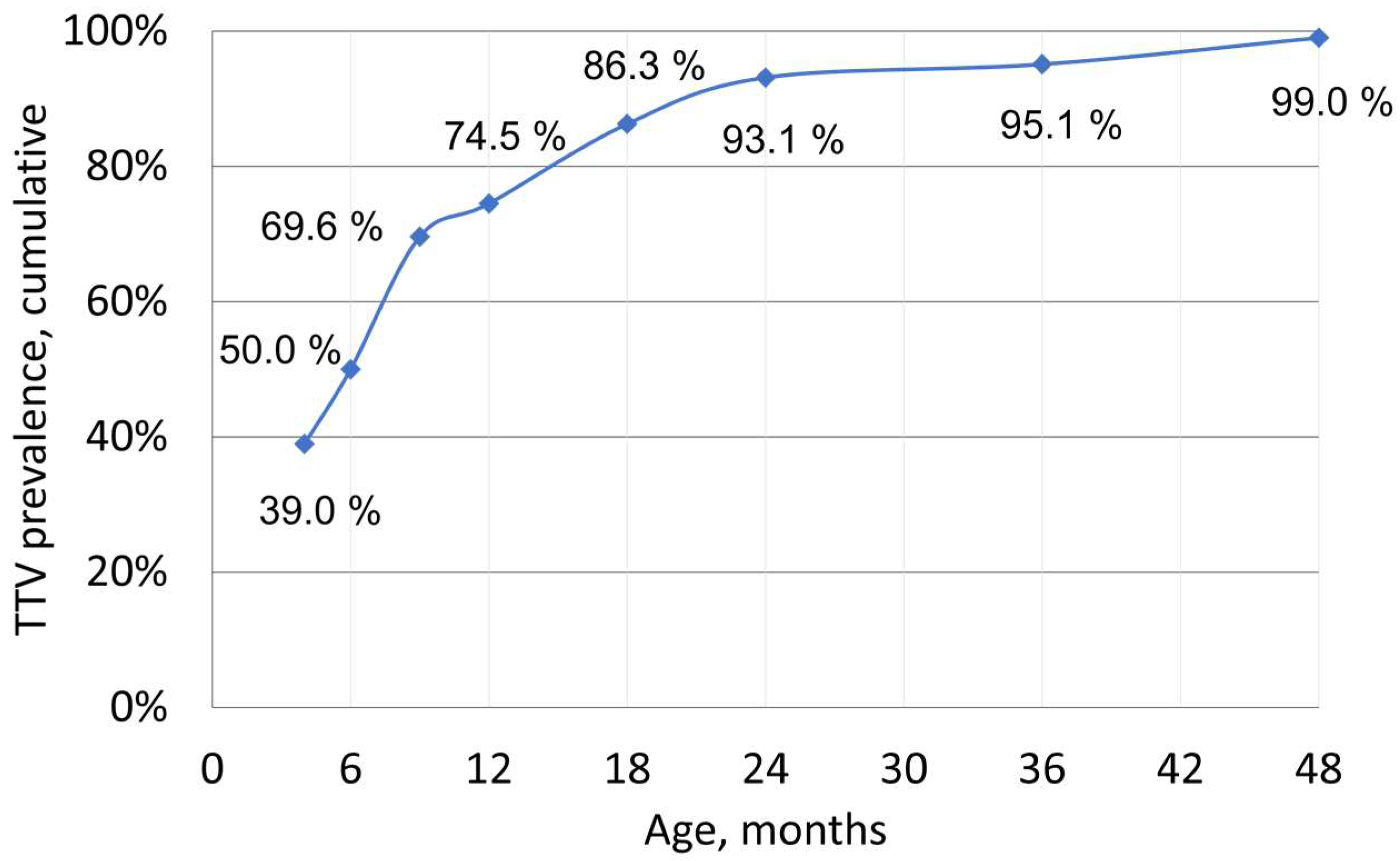
3.1. TTV Prevalence and Quantity in Children
3.2. TTV Quantity Follow-Up
3.3. TTV Strains in Follow-Up
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varsani, A.; Opriessnig, T.; Celer, V.; Maggi, F.; Okamoto, H.; Blomström, A.-L.; Cadar, D.; Harrach, B.; Biagini, P.; Kraberger, S. Taxonomic update for mammalian anelloviruses (family Anelloviridae). Arch. Virol. 2021, 166, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Kaczorowska, J.; Van Der Hoek, L. Human anelloviruses: Diverse, omnipresent and commensal members of the virome. FEMS Microbiol. Rev. 2020, 44, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Arze, C.A.; Springer, S.; Dudas, G.; Patel, S.; Bhattacharyya, A.; Swaminathan, H.; Brugnara, C.; Delagrave, S.; Ong, T.; Kahvejian, A.; et al. Global genome analysis reveals a vast and dynamic anellovirus landscape within the human virome. Cell Host Microbe 2021, 29, 1305–1315.e6. [Google Scholar] [CrossRef]
- Vasilyev, E.V.; Trofimov, D.Y.; Tonevitsky, A.G.; Ilinsky, V.V.; Korostin, D.O.; Rebrikov, D.V. Torque Teno Virus (TTV) distribution in healthy Russian population. Virol. J. 2009, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Pistello, M.; Morrica, A.; Maggi, F.; Vatteroni, M.L.; Freer, G.; Fornai, C.; Casula, F.; Marchi, S.; Ciccorossi, P.; Rovero, P.; et al. TT virus levels in the plasma of infected individuals with different hepatic and extrahepatic pathology. J. Med. Virol. 2001, 63, 189–195. [Google Scholar] [CrossRef]
- Jaksch, P.; Kundi, M.; Görzer, I.; Muraközy, G.; Lambers, C.; Benazzo, A.; Hoetzenecker, K.; Klepetko, W.; Puchhammer-Stöckl, E. Torque Teno Virus as a Novel Biomarker Targeting the Efficacy of Immunosuppression after Lung Transplantation. J. Infect. Dis. 2018, 218, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- Strassl, R.; Schiemann, M.; Doberer, K.; Görzer, I.; Puchhammer-Stöckl, E.; Eskandary, F.; Kikić, Ž.; Gualdoni, G.; Vossen, M.G.; Rasoul-Rockenschaub, S.; et al. Quantification of Torque Teno Virus Viremia as a Prospective Biomarker for Infectious Disease in Kidney Allograft Recipients. J. Infect. Dis. 2018, 218, 1191–1199. [Google Scholar] [CrossRef]
- Niel, C.; Saback, F.L.; Lampe, E. Coinfection with multiple TT virus strains belonging to different genotypes is a common event in healthy Brazilian adults. J. Clin. Microbiol. 2000, 38, 1926–1930. [Google Scholar] [CrossRef]
- Li, L.; Deng, X.; Da Costa, A.C.; Bruhn, R.; Deeks, S.G.; Delwart, E. Virome analysis of antiretroviral-treated HIV patients shows no correlation between T-cell activation and anelloviruses levels. J. Clin. Virol. 2015, 72, 106–113. [Google Scholar] [CrossRef]
- Ball, J.K.; Curran, R.; Berridge, S.; Grabowska, A.M.; Jameson, C.L.; Thomson, B.J.; Irving, W.L.; Sharp, P.M. TT virus sequence heterogeneity in vivo: Evidence for co-infection with multiple genetic types. J. Gen. Virol. 1999, 80, 1759–1768. [Google Scholar] [CrossRef][Green Version]
- Koonin, E.V.; Dolja, V.V.; Krupovic, M. The healthy human virome: From virus–host symbiosis to disease. Curr. Opin. Virol. 2021, 47, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Tyschik, E.A.; Shcherbakova, S.M.; Ibragimov, R.R.; Rebrikov, D.V. Transplacental transmission of torque teno virus. Virol. J. 2017, 14, 92. [Google Scholar] [CrossRef] [PubMed]
- Tyschik, E.A.; Rasskazova, A.S.; Degtyareva, A.V.; Rebrikov, D.V.; Sukhikh, G.T. Torque teno virus dynamics during the first year of life. Virol. J. 2018, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.S.; Zhou, Y.; Zhao, G.; Bauer, I.K.; Droit, L.; Ndao, I.M.; Warner, B.B.; Tarr, P.I.; Wang, D.; Holtz, L.R. Early life dynamics of the human gut virome and bacterial microbiome in infants. Nat. Med. 2015, 21, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- McElvania TeKippe, E.; Wylie, K.M.; Deych, E.; Sodergren, E.; Weinstock, G.; Storch, G.A. Increased Prevalence of Anellovirus in Pediatric Patients with Fever. PLoS ONE 2012, 7, e50937. [Google Scholar] [CrossRef]
- Meriluoto, M.; Hedman, L.; Tanner, L.; Simell, V.; Mäkinen, M.; Simell, S.; Mykkänen, J.; Korpelainen, J.; Ruuskanen, O.; Ilonen, J.; et al. Association of human bocavirus 1 infection with respiratory disease in childhood follow-up study Finland. Emerg. Infect. Dis. 2012, 18, 264–271. [Google Scholar] [CrossRef]
- Chen, T.; Tanner, L.; Simell, V.; Hedman, L.; Mäkinen, M.; Sadeghi, M.; Veijola, R.; Hyoty, H.; Ilonen, J.; Knip, M.; et al. Diagnostic methods for and clinical pictures of polyomavirus primary infections in children, Finland. Emerg. Infect. Dis. 2014, 20, 689–692. [Google Scholar] [CrossRef]
- Kantola, K.; Hedman, L.; Tanner, L.; Simnell, V.; Mäkinen, M.; Partanen, J.; Sadeghi, M.; Veijola, R.; Knip, M.; Ilonen, J.; et al. B-cell responses to human bocaviruses 1-4: New insights from a childhood follow-up study. PLoS ONE 2015, 10, e0139096. [Google Scholar]
- Toppinen, M.; Pratas, D.; Väisänen, E.; Söderlund-Venermo, M.; Hedman, K.; Perdomo, M.F.; Sajantila, A. The landscape of persistent human DNA viruses in femoral bone. Forensic Sci. Int. Genet. 2020, 48, 102353. [Google Scholar] [CrossRef]
- Maggi, F.; Pifferi, M.; Fornai, C.; Andreoli, E.; Tempestini, E.; Vatteroni, M.; Presciuttini, S.; Marchi, S.; Pietrobelli, A.; Boner, A.; et al. TT Virus in the Nasal Secretions of Children with Acute Respiratory Diseases: Relations to Viremia and Disease Severity. J. Virol. 2003, 77, 2418–2425. [Google Scholar] [CrossRef]
- Bagaglio, S.; Sitia, G.; Prati, D.; Cella, D.; Hasson, H.; Novati, R.; Lazzarin, A.; Morsica, G. Mother-to-child transmission of TT virus: Sequence analysis of non-coding region of TT virus in infected mother-infant pairs. Arch. Virol. 2002, 147, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Gerner, P.; Oettinger, R.; Gerner, W.; Falbrede, J.; Wirth, S. Mother-to-infant transmission of TT virus: Prevalence, extent and mechanism of vertical transmission. Pediatr. Infect. Dis. J. 2000, 19, 1074–1077. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Sugiyama, K.; Ando, T.; Mizutani, F.; Terabe, K.; Tanaka, K.; Nishiyama, M.; Wada, Y. Detection rates of TT virus DNA in serum of umbilical cord blood, breast milk and saliva. Tohoku J. Exp. Med. 2000, 191, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Antonelli, G.; Pistello, M.; Maggi, F. Torquetenovirus: The human virome from bench to bedside. Clin. Microbiol. Infect. 2016, 22, 589–593. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Oligo Name | Sequence 5′ ⟶ 3′ |
|---|---|---|
| TTV qPCR | AMTS fwd | GTGCCGNAGGTGAGTTTA |
| AMTAS rev | AGCCCGGCCAGTCC | |
| AMTAS gr4 rev | AGCCCGGCCAGACC | |
| AMTPTU probe | FAM-GGGGCAATTCGGGCT-BHQ1 | |
| TTV multiplex PCR for ~500 nt amplicon | UTR5 | GGGTGCCGRAGGTGAGTTTAC |
| 9rU | CGSCGYCTCCTTACTSTTC | |
| 9r1 | CGGCGTCTCCTTACGTTTC | |
| 9r11 | CCGYCTACTCACATATCGTC | |
| 12r | AACAGCTCGTCGAGKTCTTC | |
| 9r20 | CTSCGCCTCCTTACTCGTC | |
| 9r21 | CCTCCGYCTCCTTACTSTG | |
| 9r23 | CCTGCKCCTCCTTACTSTTG | |
| 24r | CGTCTATAGCGTCGAGSAGGTC | |
| 25r | GTCTCCTTACTCTGGGKCGTCTA | |
| 27r | AGCTCTTCGTCTGCGAGKTCT | |
| 9r46 | CATCTCCTACTTACAWATCGTC | |
| Rev primers for amplifying TTV control plasmids | p1 | GTTWGTGGTGRGCHGAAYGG |
| p1gr4 | CCACTTGTTTAGCATKAGTTTDGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Väisänen, E.; Kuisma, I.; Mäkinen, M.; Ilonen, J.; Veijola, R.; Toppari, J.; Hedman, K.; Söderlund-Venermo, M. Torque Teno Virus Primary Infection Kinetics in Early Childhood. Viruses 2022, 14, 1277. https://doi.org/10.3390/v14061277
Väisänen E, Kuisma I, Mäkinen M, Ilonen J, Veijola R, Toppari J, Hedman K, Söderlund-Venermo M. Torque Teno Virus Primary Infection Kinetics in Early Childhood. Viruses. 2022; 14(6):1277. https://doi.org/10.3390/v14061277
Chicago/Turabian StyleVäisänen, Elina, Inka Kuisma, Marjaana Mäkinen, Jorma Ilonen, Riitta Veijola, Jorma Toppari, Klaus Hedman, and Maria Söderlund-Venermo. 2022. "Torque Teno Virus Primary Infection Kinetics in Early Childhood" Viruses 14, no. 6: 1277. https://doi.org/10.3390/v14061277
APA StyleVäisänen, E., Kuisma, I., Mäkinen, M., Ilonen, J., Veijola, R., Toppari, J., Hedman, K., & Söderlund-Venermo, M. (2022). Torque Teno Virus Primary Infection Kinetics in Early Childhood. Viruses, 14(6), 1277. https://doi.org/10.3390/v14061277