
The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors



Abstract
1. Introduction
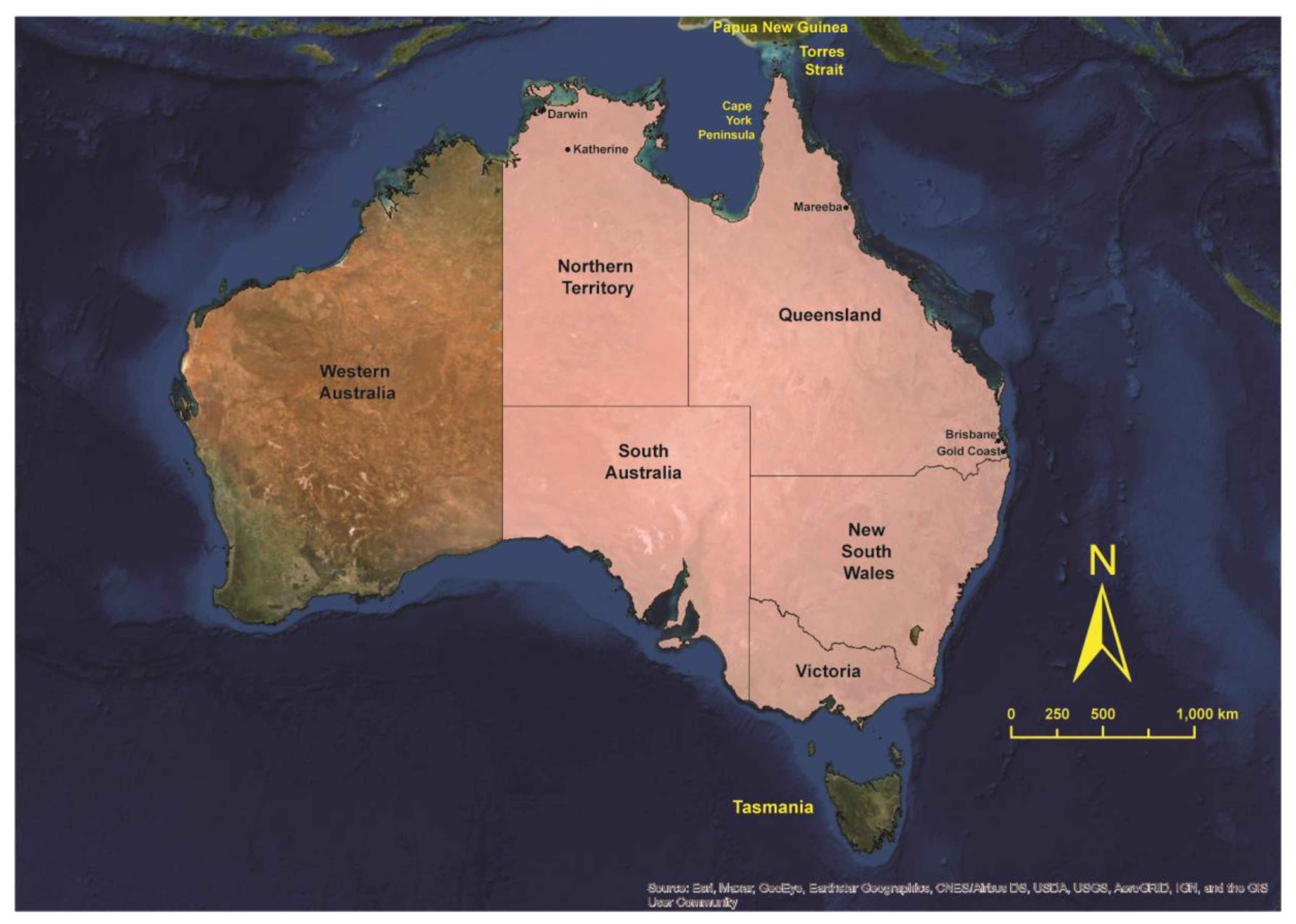
2. Incrimination of Australian Mosquitoes as Vectors of JEV
2.1. Detection of JEV in Field Populations of Mosquitoes
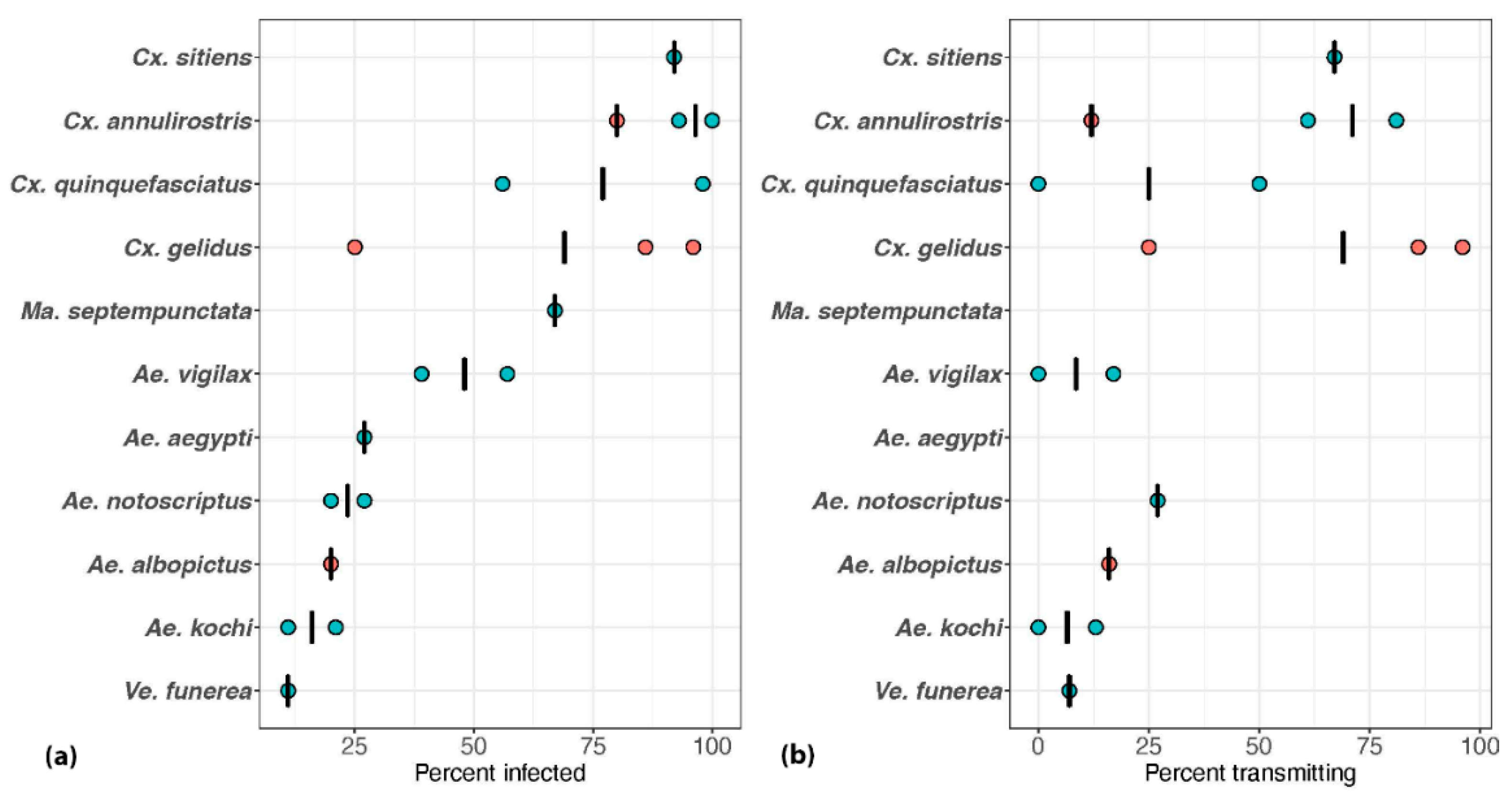
2.2. Intrinsic Ability of Australian Mosquitoes to Become Infected with and Transmit JEV
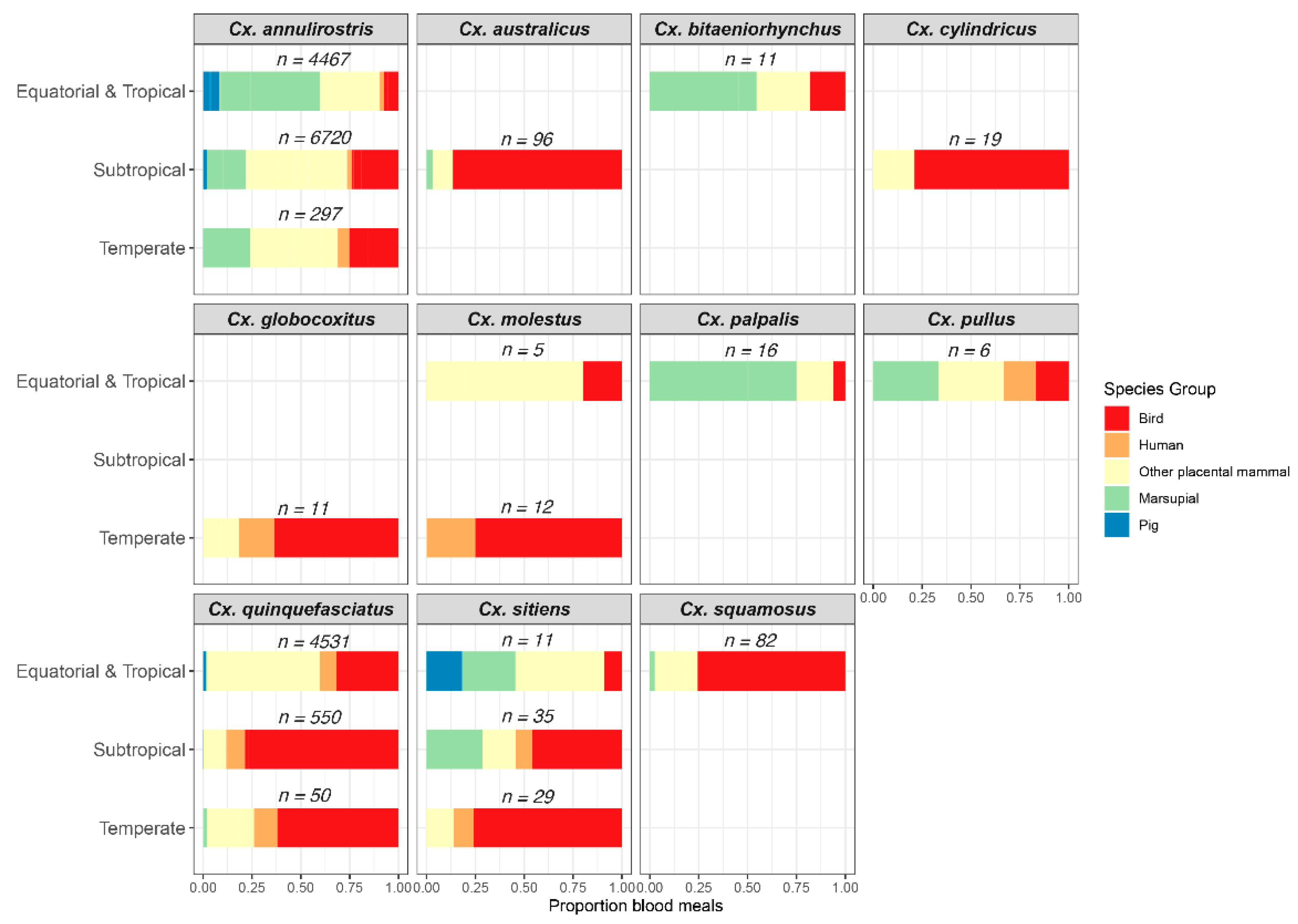
2.3. Host-Feeding Patterns of Mosquitoes Implicated as JEV Vectors
3. Australian Vectors of JEV: Reviewing the Evidence
3.1. Mosquitoes Implicated from Field and/or Laboratory Studies
3.2. Potential Australian Vectors
4. Other Factors That Impact the Role of Mosquitoes in JEV Transmission Cycles
4.1. Survival
4.2. Population Dynamics
4.3. Dispersal
4.4. Genetic Variation between Populations
4.5. Establishment of Exotic Vectors of JEV
5. Directions for Further Investigation
5.1. Detection of the Virus in Mosquito Field Populations during Periods of Recognized Virus Activity
5.2. Vector Competence of Australian Mosquito Species for the Newly Emergent JEV Genotype
5.3. The Genetic Diversity of Implicated Vectors
5.4. The Potential for JEV to Overwinter in Australian Mosquitoes
5.5. Bionomics of Invasive Australian JEV Vectors
5.6. Host-Feeding Patterns of Incriminated Vectors
5.7. Australian Vertebrate Fauna as Amplifying Hosts of JEV
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef]
- Quan, T.M.; Thao, T.T.N.; Duy, N.M.; Nhat, T.M.; Clapham, H. Estimates of the global burden of Japanese encephalitis and the impact of vaccination from 2000–2015. eLife 2020, 9, e51027. [Google Scholar] [CrossRef]
- Gao, X.; Liu, H.; Li, X.; Fu, S.; Cao, L.; Shao, N.; Zhang, W.; Wang, Q.; Lu, Z.; Lei, W.; et al. Changing Geographic Distribution of Japanese Encephalitis Virus Genotypes, 1935–2017. Vector-Borne Zoonotic Dis. 2019, 19, 35–44. [Google Scholar] [CrossRef]
- Schuh, A.J.; Ward, M.J.; Brown, A.L.; Barrett, A.D.T. Phylogeography of Japanese Encephalitis Virus: Genotype is Associated with Climate. PLOS Negl. Trop. Dis. 2013, 7, e2411. [Google Scholar] [CrossRef]
- Solomon, T.; Ni, H.; Beasley, D.W.C.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D.T. Origin and Evolution of Japanese Encephalitis Virus in Southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef]
- Buescher, E.L.; Scherer, W.F.; McClure, H.E.; Moyer, J.T.; Rosenberg, M.Z.; Yoshii, M.; Okada, Y. Ecologic Studies of Japanese Encephalitis Virus in Japan. Am. J. Trop. Med. Hyg. 1959, 8, 678–688. [Google Scholar] [CrossRef]
- Scherer, W.F.; Buescher, E.L.; McClure, H.E. Ecologic Studies of Japanese Encephalitis Virus in Japan. Am. J. Trop. Med. Hyg. 1959, 8, 689–697. [Google Scholar] [CrossRef]
- Buescher, E.L.; Scherer, W.F. Ecologic Studies of Japanese Encephalitis Virus in Japan. IX. Epidemiologic correlations and conclusions. Am. J. Trop. Med. Hyg. 1959, 8, 719–722. [Google Scholar] [CrossRef]
- Grossman, R.A.; Edelman, R.; Gould, D.J. Study of Japanese encephalitis virus in Chiangmai Valley, Thailand. VI. Summary and conclusions. Am. J. Epidemiol. 1974, 100, 69–76. [Google Scholar] [CrossRef]
- Endy, T.P.; Nisalak, A. Japanese Encephalitis Virus: Ecology and Epidemiology. Curr. Top. Microbiol. Immunol. 2002, 267, 11–48. [Google Scholar] [CrossRef]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging flaviviruses: The spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Shield, J.; Bailey, M.C.; Mackenzie, J.S.; Poidinger, M.; McCall, B.J.; Mills, P.J. An outbreak of Japanese encephalitis in the Torres Strait, Australia, 1995. Med. J. Aust. 1996, 165, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Hanna, J.N.; Ritchie, S.A.; Phillips, D.A.; Lee, J.M.; Hills, S.; van den Hurk, A.F.; Pyke, A.; Johansen, C.A.; Mackenzie, J.S. Japanese encephalitis in north Queensland, Australia, 1998. Med. J. Aust. 1999, 170, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.A.; van den Hurk, A.F.; Ritchie, S.A.; Zborowski, P.; Paru, R.; Bockarie, M.J.; Drew, A.C.; Khromykh, T.I.; Mackenzie, J.S. Isolation of Japanese encephalitis virus from mosquitoes (Diptera: Culicidae) collected in the Western Province of Papua New Guinea, 1997–1998. Am. J. Trop. Med. Hyg. 2000, 62, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Johansen, C.A.; Ritchie, S.A.; van den Hurk, A.F.; Hall, R.A. The Emergence and Spread of Japanese Encephalitis Virus in Australasia. Curr. Top. Microbiol. Immunol. 2002, 267, 49–73. [Google Scholar] [CrossRef]
- Ritchie, S.A.; Rochester, W. Wind-Blown Mosquitoes and Introduction of Japanese Encephalitis into Australia. Emerg. Infect. Dis. 2001, 7, 900–908. [Google Scholar] [CrossRef]
- Shield, J.; Hanna, J.; Phillips, D. Reappearance of the Japanese encephalitis virus in the Torres Strait, 1996. Commun. Dis. Intell. 1996, 20, 191. [Google Scholar]
- van den Hurk, A.F.; Pyke, A.T.; Mackenzie, J.S.; Hall-Mendelin, S.; Ritchie, S.A. Japanese Encephalitis Virus in Australia: From Known Known to Known Unknown. Trop. Med. Infect. Dis. 2019, 4, 38. [Google Scholar] [CrossRef]
- Pyke, A.T.; Williams, D.T.; Nisbet, D.J.; van den Hurk, A.F.; Taylor, C.T.; Johansen, C.A.; Macdonald, J.; Hall, R.A.; Simmons, R.J.; Mason, R.J.V.; et al. The appearance of a second genotype of Japanese encephalitis virus in the Australasian region. Am. J. Trop. Med. Hyg. 2001, 65, 747–753. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Montgomery, B.L.; Northill, J.A.; Smith, I.L.; Zborowski, P.; Ritchie, S.A.; Mackenzie, J.S.; Smith, G.A. The First Isolation of Japanese Encephalitis Virus from Mosquitoes Collected from Mainland Australia. Am. J. Trop. Med. Hyg. 2006, 75, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S. Japanese encephalitis: An emerging disease in the Australasian region, and its potential risk to Australia. Arbovirus Res. Aust. 1997, 7, 166–170. [Google Scholar]
- Northern Territory Government. Japanese Encephalitis. Available online: https://nt.gov.au/wellbeing/health-conditions-treatments/viral/japanese-encephalitis (accessed on 4 May 2022).
- Ritchie, S.A.; van den Hurk, A.F.; Zborowski, P.; Kerlin, T.J.; Banks, D.; Walker, J.A.; Lee, J.M.; Montgomery, B.L.; Smith, G.A.; Pyke, A.T.; et al. Operational Trials of Remote Mosquito Trap Systems for Japanese Encephalitis Virus Surveillance in the Torres Strait, Australia. Vector-Borne Zoonotic Dis. 2007, 7, 497–506. [Google Scholar] [CrossRef] [PubMed]
- World Organisation for Animal Health. OIE Immediate Notification—Japanese Encephalitis, Australia; World Organization for Animal Health: Paris, France, 2022. [Google Scholar]
- Australian Government Department of Agriculture, Water and the Environment. National Pest and Disease Outbreaks—Japanese Encephalitis. Available online: https://www.outbreak.gov.au/current-responses-to-outbreaks/japanese-encephalitis (accessed on 4 May 2022).
- Australian Government Department of Health. Japanese Encephalitis Virus (JEV). Available online: https://www.health.gov.au/health-alerts/japanese-encephalitis-virus-jev/about (accessed on 4 May 2022).
- Reeves, W.C. Arthropods as vectors and reservoirs of animal pathogenic viruses. In Handbuch der Virus Forschung; Hallauer, C., Meyer, K.F., Eds.; Springer: Vienna, Austria, 1957; Volume 4, pp. 177–202. [Google Scholar]
- Turell, M.J.; Dohm, D.J.; Sardelis, M.R.; O’Guinn, M.L.; Andreadis, T.G.; Blow, J.A. An Update on the Potential of North American Mosquitoes (Diptera: Culicidae) to Transmit West Nile Virus. J. Med. Entomol. 2005, 42, 57–62. [Google Scholar] [CrossRef] [PubMed]
- van den Hurk, A.F.; Ritchie, S.A.; Smith, G.A.; Montgomery, B.L.; Mackenzie, J.S. A Japanese encephalitis odyssey: Entomological studies in northern Australia, 2002–2004. Arbovirus Res. Aust. 2005, 9, 370–377. [Google Scholar]
- Beebe, N.W.; Russell, T.; Burkot, T.R.; Cooper, R.D. Anopheles punctulatus Group: Evolution, Distribution, and Control. Annu. Rev. Entomol. 2015, 60, 335–350. [Google Scholar] [CrossRef]
- Jansen, C.C.; Hemmerter, S.; van den Hurk, A.F.; Whelan, P.I.; Beebe, N.W. Morphological versus molecular identification of Culex annulirostris and Culex palpalis—Key members of the Culex sitiens (Diptera: Culicidae) subgroup in Australasia. Aust. J. Entomol. 2013, 52, 356–362. [Google Scholar] [CrossRef]
- Beebe, N.W.; van den Hurk, A.F.; Chapman, H.F.; Frances, S.P.; Williams, C.R.; Cooper, R.D. Development and evaluation of a species diagnostic polymerase chain reaction-restriction fragment-length polymorphism procedure for cryptic members of the Culex sitiens (Diptera: Culicidae) subgroup in Australia and the southwest Pacific. J. Med. Entomol. 2002, 39, 362–369. [Google Scholar] [CrossRef]
- Chapman, H.F.; Kay, B.H.; Ritchie, S.A.; van den Hurk, A.F.; Hughes, J.M. Definition of species in the Culex sitiens subgroup (Diptera: Culicidae) from Papua New Guinea and Australia. J. Med. Entomol. 2000, 37, 736–742. [Google Scholar] [CrossRef]
- Broom, A.K.; Hall, R.A.; Johansen, C.A.; Oliveira, N.; Howard, M.A.; Lindsay, M.D.; Kay, B.H.; Mackenzie, J.S. Identification of Australian arboviruses in inoculated cell cultures using monoclonal antibodies in ELISA. Pathology 1998, 30, 286–288. [Google Scholar] [CrossRef]
- Pyke, A.T.; Smith, I.L.; van den Hurk, A.F.; Northill, J.A.; Chuan, T.F.; Westacott, A.J.; Smith, G.A. Detection of Australasian Flavivirus encephalitic viruses using rapid fluorogenic TaqMan RT-PCR assays. J. Virol. Methods 2004, 117, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Phillips, D.; Broom, A.; Mackenzie, J.; Poidinger, M.; Hurk, A.V.D. Isolation of Japanese Encephalitis Virus from Culex annulirostris in Australia. Am. J. Trop. Med. Hyg. 1997, 56, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.A.; van den Hurk, A.F.; Pyke, A.T.; Zborowski, P.; Phillips, D.A.; Mackenzie, J.S.; Ritchie, S.A. Entomological Investigations of an Outbreak of Japanese Encephalitis Virus in the Torres Strait, Australia, in 1998. J. Med. Entomol. 2001, 38, 581–588. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Nisbet, D.J.; Johansen, C.A.; Foley, P.N.; Ritchie, S.A.; Mackenzie, J.S. Japanese encephalitis on Badu Island, Australia: The first isolation of Japanese encephalitis virus from Culex gelidus in the Australasian region and the role of mosquito host-feeding patterns in virus transmission cycles. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 595–600. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Ritchie, S.A.; Johansen, C.A.; Mackenzie, J.S.; Smith, G.A. Domestic Pigs and Japanese Encephalitis Virus Infection, Australia. Emerg. Infect. Dis. 2008, 14, 1736–1738. [Google Scholar] [CrossRef]
- Johansen, C.A.; Nisbet, D.J.; Foley, P.N.; van den Hurk, A.F.; Hall, R.A.; Mackenzie, J.S.; Ritchie, S.A. Flavivirus isolations from mosquitoes collected from Saibai Island in the Torres Strait, Australia, during an incursion of Japanese encephalitis virus. Med. Veter. Entomol. 2004, 18, 281–287. [Google Scholar] [CrossRef]
- Vythilingam, I.; Oda, K.; Mahadevan, S.; Abdullah, G.; Thim, C.S.; Hong, C.C.; Vijayamalar, B.; Sinniah, M.; Igarashi, A. Abundance, Parity, and Japanese Encephalitis Virus Infection of Mosquitoes (Diptera: Culicidae) in Sepang District, Malaysia. J. Med. Entomol. 1997, 34, 257–262. [Google Scholar] [CrossRef]
- Vythilingam, I.; Oda, K.; Tsuchie, H.; Mahadevan, S.; Vijayamalar, B. Isolation of Japanese encephalitis virus from Culex sitiens mosquitoes in Selangor, Malaysia. J. Am. Mosq. Control Assoc. 1994, 10, 228–229. [Google Scholar]
- Weng, M.H.; Lien, J.C.; Wang, Y.M.; Lin, C.C.; Lin, H.C.; Chin, C. Isolation of Japanese encephalitis virus from mosquitoes collected in Northern Taiwan between 1995 and 1996. J. Microbiol. Immunol. Infect. 1999, 32, 9–13. [Google Scholar]
- Johnson, P.H.; Hall-Mendelin, S.; Whelan, P.I.; Frances, S.P.; Jansen, C.C.; Mackenzie, D.O.; Northill, J.A.; van den Hurk, A.F. Vector competence of Australian Culex gelidus Theobald (Diptera: Culicidae) for endemic and exotic arboviruses. Aust. J. Entomol. 2009, 48, 234–240. [Google Scholar] [CrossRef]
- Nicholson, J.; Ritchie, S.A.; van den Hurk, A.F. Aedes albopictus (Diptera: Culicidae) as a Potential Vector of Endemic and Exotic Arboviruses in Australia. J. Med. Entomol. 2014, 51, 661–669. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Johnson, P.H.; Hall-Mendelin, S.; Northill, J.A.; Simmons, R.J.; Jansen, C.C.; Frances, S.P.; Smith, G.A.; Ritchie, S.A. Expectoration of Flaviviruses during sugar feeding by mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 845–850. [Google Scholar] [CrossRef]
- van den Hurk, A.F.; Nisbet, D.J.; Hall, R.A.; Kay, B.H.; Mackenzie, J.S.; Ritchie, S.A. Vector Competence of Australian Mosquitoes (Diptera: Culicidae) for Japanese Encephalitis Virus. J. Med. Entomol. 2003, 40, 82–90. [Google Scholar] [CrossRef]
- Takahashi, M. Varaiation in susceptibility among colony strains of Culex tritaeniorhynchus to Japanese encephalitis virus infection. Jap. J. Med. Sci. Biol. 1980, 33, 321–329. [Google Scholar] [CrossRef]
- Takahashi, M. Differential transmission efficiency for Japanese encephalitis virus among colonised strains of Culex tritaeniorhynchus. Jap. J. San. Zool. 1982, 33, 325–333. [Google Scholar] [CrossRef]
- Kay, B.; Fanning, I.; Carley, J. The vector competence of Australian Culex annulirostris with Murray Valley encephalitis and Kunjin viruses. Aust. J. Exp. Biol. Med. Sci. 1984, 62, 641–650. [Google Scholar] [CrossRef]
- Azar, S.R.; Weaver, S.C. Vector Competence: What Has Zika Virus Taught Us? Viruses 2019, 11, 867. [Google Scholar] [CrossRef]
- Takahashi, M. The Effects of Environmental and Physiological Conditions of Culex tritaeniorhynchus on the Pattern of Transmission of Japanese Encephalitis Virus. J. Med. Entomol. 1976, 13, 275–284. [Google Scholar] [CrossRef]
- Blackwell, A.; Mordue, A.J.M.; Mordue, W. Identification of bloodmeals of the Scottish biting midge, Culicoides impunctatus, by indirect enzyme-linked immunosorbent assay (ELISA). Med. Vet. Entomol. 1994, 8, 20–24. [Google Scholar] [CrossRef]
- Ngo, K.A.; Kramer, L.D. Identification of Mosquito Bloodmeals Using Polymerase Chain Reaction (PCR) with Order-Specific Primers. J. Med. Entomol. 2003, 40, 215–222. [Google Scholar] [CrossRef]
- Flies, E.J.; Flies, A.S.; Fricker, S.R.; Weinstein, P.; Williams, C.R. Regional Comparison of Mosquito Bloodmeals in South Australia: Implications for Ross River Virus Ecology. J. Med. Entomol. 2016, 53, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Frances, S.P.; Cooper, R.D.; Rowcliffe, K.L.; Chen, N.; Cheng, Q. Occurrence of Ross River Virus and Barmah Forest Virus in Mosquitoes at Shoalwater Bay Military Training Area, Queensland, Australia. J. Med. Entomol. 2004, 41, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Hall-Mendelin, S.; Jansen, C.C.; Cheah, W.Y.; Montgomery, B.L.; Hall, R.A.; Ritchie, S.A.; van den Hurk, A.F. Culex annulirostris (Diptera: Culicidae) host feeding patterns and Japanese encephalitis virus ecology in northern Australia. J. Med. Entomol. 2012, 49, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.C.; Webb, C.E.; Graham, G.C.; Craig, S.B.; Zborowski, P.; Ritchie, S.A.; Russell, R.C.; van den Hurk, A.F. Blood Sources of Mosquitoes Collected from Urban and Peri-Urban Environments in Eastern Australia with Species-Specific Molecular Analysis of Avian Blood Meals. Am. J. Trop. Med. Hyg. 2009, 81, 849–857. [Google Scholar] [CrossRef]
- Johansen, C.A.; Power, S.L.; Broom, A.K. Determination of Mosquito (Diptera: Culicidae) Bloodmeal Sources in Western Australia: Implications for Arbovirus Transmission. J. Med. Entomol. 2009, 46, 1167–1175. [Google Scholar] [CrossRef][Green Version]
- Kay, B.H.; Boreham, P.F.L.; Fanning, I.D. Host-Feeding Patterns of Culex annulirostris and Other Mosquitoes (Diptera: Culicidae) at Charleville, Southwestern Queensland, Australia. J. Med. Entomol. 1985, 22, 529–535. [Google Scholar] [CrossRef]
- Kay, B.H.; Boreham, P.F.L.; Williams, G.M. Host preferences and feeding patterns of mosquitoes (Diptera: Culicidae) at Kowanyama, Cape York Peninsula, northern Queensland. Bull. Entomol. Res. 1979, 69, 441–457. [Google Scholar] [CrossRef]
- Kay, B.H.; Ryan, P.A.; Hall, R.A.; Boyd, A.M. Mosquito feeding patterns and natural infection of vertebrates with Ross River and Barmah Forest viruses in Brisbane, Australia. Am. J. Trop. Med. Hyg. 2007, 76, 417–423. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Johansen, C.A.; Zborowski, P.; Paru, R.; Foley, P.N.; Beebe, N.W.; Mackenzie, J.S.; Ritchie, S.A. Mosquito host-feeding patterns and implications for Japanese encephalitis virus transmission in northern Australia and Papua New Guinea. Med. Vet. Entomol. 2003, 17, 403–411. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Smith, I.L.; Smith, G.A. Development and evaluation of real-time polymerase chain reaction assays to identify mosquito (Diptera: Culicidae) blood meals originating from native Australian mammals. J. Med. Entomol. 2007, 44, 85–92. [Google Scholar] [CrossRef]
- Daniels, P.; Middleton, D.; Lunt, R.A. Assessment of the Potential of Australian Fauna as Maintenance or Amplifying Hosts of Japanese Encephalitis (JE) Virus; CSIRO Australian Animal Health Laboratory: Geelong, Australia, 2000; pp. 1–3. [Google Scholar]
- van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and Geographical Expansion of Japanese Encephalitis Virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C. Dispersal of the arbovirus vector Culex annulirostris Skuse (Diptera: Culicidae) in the Murray VaIIey of Victoria, Australia. Gen. Appl. Entomol. 1986, 18, 5–9. [Google Scholar]
- Banerjee, K.; Deshmukh, P.K. Transmission of Japanese encephalitis virus to chicks by individual Culex bitaeniorhynchus mosquitoes. Indian J. Med. Res. 1987, 86, 726–727. [Google Scholar] [PubMed]
- Banerjee, K.; Deshmukh, P.K.; Ilkal, M.A.; Dhanda, V. Experimental transmission of Japanese encephalitis virus through Anopheles tessellatus and Culex fatigans mosquitoes. Indian J. Med. Res. 1977, 65, 746–752. [Google Scholar] [PubMed]
- Gould, D.J.; Barnett, H.C.; Suyemoto, W. Transmission of Japanese encephalitis virus by Culex gelidus Theobald. Trans. R. Soc. Trop. Med. Hyg. 1962, 56, 429–435. [Google Scholar] [CrossRef]
- Vythilingam, I.; Oda, K.; Chew, T.K.; Mahadevan, S.; Vijayamalar, B.; Morita, K.; Tsuchie, H.; Igarashi, A. Isolation of Japanese encephalitis virus from mosquitoes collected in Sabak Bernam, Selangor, Malaysia in 1992. J. Am. Mosq. Control Assoc. 1995, 11, 94–98. [Google Scholar]
- Dhanda, V.; Thenmozhi, V.; Kumar, N.P.; Hiriyan, J.; Arunachalam, N.; Balasubramanian, A.; Ilango, A.; Gajanana, A. Virus isolation from wild-caught mosquitoes during a Japanese encephalitis outbreak in Kerala in 1996. Indian J. Med. Res. 1997, 106, 4–6. [Google Scholar]
- Frances, S.P.; Mackenzie, D.O.; Rowcliffe, K.L.; Corcoran, S.K. Comparative Field Evaluation of Repellent Formulations Containing Deet and IR3535 Against Mosquitoes in Queensland, Australia. J. Am. Mosq. Control Assoc. 2009, 25, 511–513. [Google Scholar] [CrossRef]
- Kay, B.H.; Watson, T.M.; Ryan, P.A. Definition of productive Aedes notoscriptus (Diptera: Culicidae) habitats in western Brisbane, and a strategy for their control. Aust. J. Entomol. 2008, 47, 142–148. [Google Scholar] [CrossRef]
- Rosen, L.; Tesh, R.B.; Lien, J.C.; Cross, J.H. Transovarial Transmission of Japanese Encephalitis Virus by Mosquitoes. Science 1978, 199, 909–911. [Google Scholar] [CrossRef]
- Marshall, I.D. Epidemiology of Murray Valley encephalitis in eastern Australia—Patterns of arbovirus activity and strategies of arbovirus survival. Arbovirus Res. Aust. 1979, 2, 47–53. [Google Scholar]
- Marshall, I.D.; Woodroofe, G.M.; Hirsch, S. Viruses recovered from mosquitoes and wildlife serum collected in the Murray Valley of south-eastern Australia, February 1974, during an epidemic of encephalitis. Aust. J. Exp. Biol. Med. Sci. 1982, 60, 457–470. [Google Scholar] [CrossRef] [PubMed]
- McLean, D.M. Transmission of Murray Valley encephalitis virus by mosquitoes. Aust. J. Exp. Biol. Med. Sci. 1953, 31, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Van den Hurk, A.F.; Hall-Mendelin, S.; Webb, C.E.; Tan, C.S.; Frentiu, F.D.; Prow, N.A.; Hall, R.A. Role of enhanced vector transmission of a new West Nile virus strain in an outbreak of equine disease in Australia in 2011. Parasites Vectors 2014, 7, 586. [Google Scholar] [CrossRef]
- Russell, R.C. A Review of the Status and Significance of the Species Within the Culex pipiens Group in Australia. J. Am. Mosq. Control Assoc. 2012, 28, 24–27. [Google Scholar] [CrossRef]
- Weng, M.-H.; Lien, J.-C.; Lin, C.-C.; Yao, C.-W. Vector Competence of Culex pipiens molestus (Diptera: Culicidae) from Taiwan for a Sympatric Strain of Japanese Encephalitis Virus. J. Med. Entomol. 2000, 37, 780–783. [Google Scholar] [CrossRef]
- Dhileepan, K. Mosquito seasonality and arboviral disease incidence in Murray Valley, southeast Australia. Med. Vet. Entomol. 1996, 10, 375–384. [Google Scholar] [CrossRef]
- Kay, B.H.; Fanning, I.D.; Mottram, P. The vector competence of Culex annulirostris, Aedes sagax and Aedes alboannulatus for Murray Valley encephalitis virus at different temperatures. Med. Veter. Entomol. 1989, 3, 107–112. [Google Scholar] [CrossRef]
- Wada, Y. Theoretical considerations on the epidemic of Japanese encephalitis. Trop. Med. 1975, 16, 171–199. [Google Scholar]
- Kay, B.H.; Saul, A.J.; McCullagh, A. A mathematical model for the rural amplification of Murray Valley encephalitis virus in southern Australia. Am. J. Epidemiol. 1987, 125, 690–705. [Google Scholar] [CrossRef]
- Kay, B.H. Age Structure of Populations of Culex annulirostris (Diptera: Culicidae) at Kowanyama and Charleville, Queensland. J. Med. Entomol. 1979, 16, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C. Population age composition and female longevity of the arbovirus vector Culex annulirostris Skuse near Echuca, Victoria, in the Murray valley of southeastern Australia 1979–1985. Aust. J. Exp. Biol. Med. Sci. 1986, 64, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Kay, B. Seasonal abundance of Culex annulirostris and other mosquitoes at Kowanyama, north Queensland, and Charleville, south west Queensland. Aust. J. Exp. Biol. Med. Sci. 1979, 57, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.C. Seasonal Abundance and Age Composition of Two Populations of Culex annulirostris (Diptera: Culicidae) at Darwin, Northern Territory, Australia. J. Med. Entomol. 1986, 23, 279–285. [Google Scholar] [CrossRef]
- Russell, R.C. Seasonal activity and abundance of the arbovirus vector Culex annulirostris Skuse near Echuca, Victoria, in the Murray valley of southeastern Australia 1979-1985. Aust. J. Exp. Biol. Med. Sci. 1986, 64, 97–103. [Google Scholar] [CrossRef]
- Russell, R.C. Culex annuliroslris Skuse (Diptera: Culicidae) at Appin, N.S.W.- bionomics and behaviour. J. Aust. Enlomol. Soc. 1986, 25, 103–109. [Google Scholar] [CrossRef]
- Doggett, S.; Clancy, J.; Haniotis, J.; Webb, C.; Russell, R.C.; Hueston, L.; Dwyer, D.E. The New South Wales Arbovirus Surveillance and Mosquito Monitoring Program 2010–2011 Annual Report; Department of Medical Entomology, ICPMR, Westmead Hospital: Westmead, Australia, 2011; p. 37. [Google Scholar]
- Marshall, I.D.; Thibos, E.; Clarke, K. Species composition of mosquitoes collected in the Murray Valley of south-eastern Australia during an epidemic of arboviral encephalitis. Aust. J. Exp. Biol. Med. Sci. 1982, 60, 447–456. [Google Scholar] [CrossRef]
- McDonald, G.; McLaren, I.W.; Shelden, G.P.; Smith, I.R. The effect of temperature on the population growth potential of Culex annulirostris Skuse (Diptera: Culicidae). Austral. Ecol. 1980, 5, 379–384. [Google Scholar] [CrossRef]
- Russell, R.C. Age composition and overwintering of Culex annulirostris Skuse (Diptera: Culicidae) near Deniliquin, in the Murray Valley of New South Wales. Aust. J. Entomol. 1987, 26, 93–96. [Google Scholar] [CrossRef]
- Bryan, J.H.; O’Donnell, M.S.; Berry, G.; Carvan, T. Dispersal of adult female Culex annulirostris in Griffith, New South Wales, Australia: A further study. J. Am. Mosq. Control Assoc. 1992, 8, 398–403. [Google Scholar]
- O’Donnell, M.S.; Berry, G.; Carvan, T.; Bryan, J.H. Dispersal of adult females of Culex annulirostris in Griffith, New South Wales, Australia. J. Am. Mosq. Control Assoc. 1992, 8, 159–165. [Google Scholar] [PubMed]
- Mackenzie, J.S.; Lindsay, M.D.; Daniels, P.W. The effect of climate on the incidence of vector-borne viral diseases: The potential value of seasonal forecasting. In Applications of Seasonal Climate Forecasting in Agriculture and Natural Ecosystems—The Australian Experience; Hammer, G., Nicholls, N., Mitchell, C., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 429–452. [Google Scholar]
- Kay, B.H.; Farrow, R.A. Mosquito (Diptera: Culicidae) Dispersal: Implications for the Epidemiology of Japanese and Murray Valley Encephalitis Viruses in Australia. J. Med. Entomol. 2000, 37, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.-G.; Jin, H.; Riley, J.R.; Reynolds, D.R.; Smith, A.D.; Wang, R.-L.; Cheng, J.-Y.; Cheng, X.-N. Autumn southward “return” migration of the mosquito Culex tritaeniorhynchus in China. Med. Vet. Entomol. 1993, 7, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.; Smith, A.D.; Mukhopadhyay, S.; Chowdhury, A.K.; De, B.K.; Nath, P.S.; Mondal, S.K.; Das, B.K.; Mukhopadhyay, S. Atmospheric transport of mosquitoes in northeast India. Med. Vet. Entomol. 1996, 10, 185–186. [Google Scholar] [CrossRef] [PubMed]
- Hemmerter, S.; Šlapeta, J.; van den Hurk, A.F.; Cooper, R.D.; Whelan, P.I.; Russell, R.C.; Johansen, C.A.; Beebe, N.W. A curious coincidence: Mosquito biodiversity and the limits of the Japanese encephalitis virus in Australasia. BMC Evol. Biol. 2007, 7, 100. [Google Scholar] [CrossRef] [PubMed]
- Aardema, M.L.; Vonholdt, B.M.; Fritz, M.L.; Davis, S.R. Global evaluation of taxonomic relationships and admixture within the Culex pipiens complex of mosquitoes. Parasites Vectors 2020, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.J.; Montgomery, B.L.; Ingram, A.; Ritchie, S.A. First records of Culex gelidus from Australia. J. Am. Mosq. Control Assoc. 2001, 17, 79–80. [Google Scholar]
- Whelan, P.; Hayes, G.; Carter, J.; Wilson, A.; Haigh, B. Detection of the exotic mosquito Culex gelidus in the Northern Territory. Commun. Dis. Intell. 2000, 24, 74–75. [Google Scholar]
- Ritchie, S.; Haseler, B.; Foley, P.; Montgomery, B. Exotic mosquitoes in north Queensland: The true millenium bug. Arbovirus Res. Aust. 2001, 8, 288–293. [Google Scholar]
- Williams, C.R.; Ritchie, S.A.; Whelan, P.I. Potential distribution of the Asian disease vector Culex gelidus Theobald (Diptera: Culicidae) in Australia and New Zealand: A prediction based on climate suitability. Aust. J. Entomol. 2005, 44, 425–430. [Google Scholar] [CrossRef]
- Lessard, B.D.; Kurucz, N.; Rodriguez, J.; Carter, J.; Hardy, C.M. Detection of the Japanese encephalitis vector mosquito Culex tritaeniorhynchus in Australia using molecular diagnostics and morphology. Parasites Vectors 2021, 14, 411. [Google Scholar] [CrossRef] [PubMed]
- Fansiri, T.; Fontaine, A.; Diancourt, L.; Caro, V.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Ponlawat, A.; Lambrechts, L. Genetic Mapping of Specific Interactions between Aedes aegypti Mosquitoes and Dengue Viruses. PLoS Genet. 2013, 9, e1003621. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Mifune, K.; Matsuo, S.; Shichijo, A.; Suzuki, H.; Ura, M.; Makino, Y.; Wada, Y.; Oda, T.; Mogi, M.; et al. Ecology of Japanese encephalitis virus in Japan, particularly the results of surveys in every interepidemic season from 1964 to 1976. Trop. Med. 1978, 20, 81–96. [Google Scholar]
- Lee, H.W. Study on overwintering mechanisms of Japanese encephalitis virus in Korea. J. Korean Med. Assoc. 1971, 14, 871–878. [Google Scholar]
- Stephenson, E.B.; Murphy, A.K.; Jansen, C.C.; Peel, A.J.; McCallum, H. Interpreting mosquito feeding patterns in Australia through an ecological lens: An analysis of blood meal studies. Parasites Vectors 2019, 12, 156. [Google Scholar] [CrossRef] [PubMed]
- Boyle, D.B.; Dickerman, R.W.; Marshall, I.D. Primary viraemia responses of herons to experimental infection with Murray Valley encephalitis, Kunjin and Japanese encephalitis viruses. Aust. J. Exp. Biol. Med. Sci. 1983, 61, 655–664. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Location | Year | Number of Culex Processed | Number of Mosquitoes of Other Genera Processed | Trapping Method a | Number of JEV Positive Pools | Composition of Positive Pools | Reference |
|---|---|---|---|---|---|---|---|
| Badu (Torres Strait) | 1995 | 2987 | 10,313 | CDC-LT | 8 | Culex sitiens subgroup b | [37] |
| 1998 | 26,158 | 5730 | CDC-LT | 42 | Cx. sitiens subgroup | [38] | |
| CDC-LT | 1 | Aedes vigilax | |||||
| 2000 | 7779 | 83,461 | CDC-LT | 1 | Culex gelidus | [39] | |
| 2002 | Not separated by taxonomic group c | MM | 3 | Unknown | [24] | ||
| 2003 | 22,157 | Not processed d | CDC-LT, MM | 18 | Cx. sitiens subgroup | [24,40] | |
| CDC-LT, MM | 2 | Cx. gelidus | [24], this paper | ||||
| CDC-GT | 1 | Culex bitaeniorhynchus | This paper | ||||
| 2004 | 1795 | Not processed | NMT | 1 | Cx. gelidus | [24] | |
| 2005 | 4563 | Not processed | NMT | 1 | Cx. sitiens subgroup | [24] | |
| Moa (Torres Strait) | 2003 | 1400 | Not processed | MM | 2 | Cx. sitiens subgroup | [24] |
| MM | 1 | Cx. gelidus | [24] | ||||
| Cape York Peninsula | 2004 | 23,144 | Not processed | CDC-LT | 1 | Cx. sitiens subgroup | [21] |
| Saibai (Torres Strait) | 2000 | 45,581 | 38,473 | CDC-LT | 1 | Cx. sitiens subgroup | [41] |
| Papua New Guinea | 1997 | 35,038 | 25,445 | CDC-LT | 1 | Cx. sitiens subgroup | [15] |
| 1998 | 210,200 | 49,717 | CDC-LT | 2 | Cx. sitiens subgroup | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van den Hurk, A.F.; Skinner, E.; Ritchie, S.A.; Mackenzie, J.S. The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors. Viruses 2022, 14, 1208. https://doi.org/10.3390/v14061208
van den Hurk AF, Skinner E, Ritchie SA, Mackenzie JS. The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors. Viruses. 2022; 14(6):1208. https://doi.org/10.3390/v14061208
Chicago/Turabian Stylevan den Hurk, Andrew F., Eloise Skinner, Scott A. Ritchie, and John S. Mackenzie. 2022. "The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors" Viruses 14, no. 6: 1208. https://doi.org/10.3390/v14061208
APA Stylevan den Hurk, A. F., Skinner, E., Ritchie, S. A., & Mackenzie, J. S. (2022). The Emergence of Japanese Encephalitis Virus in Australia in 2022: Existing Knowledge of Mosquito Vectors. Viruses, 14(6), 1208. https://doi.org/10.3390/v14061208