
A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Strains and Clinical Samples
2.2. Primer and Probe Design
2.3. Cloning of Recombinant Plasmids and Preparation of RNA Reference Transcripts
2.4. Conventional RT-PCR Assays
2.5. Real Time RT-PCR Assays
2.6. Determination of Analytical Sensitivity and Specificity
2.7. Statistical Analysis
3. Results
3.1. Conventional RT-PCR and Comparison with Published Primers Detecting a Broad Range of Bornaviruses
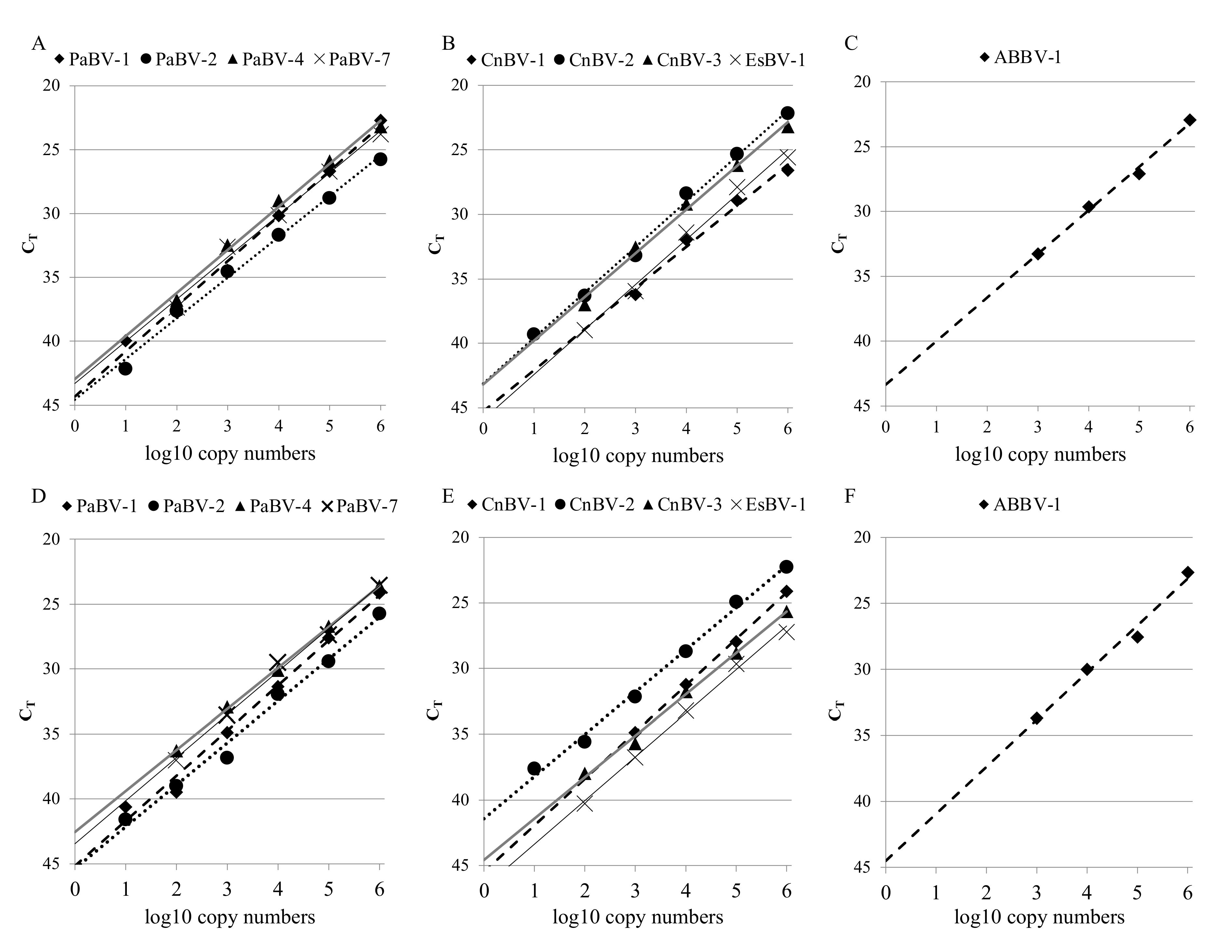
3.2. Analytical Sensitivity and Specificity of the Avian Bornavirus Multiplex rRT-PCR Assay
3.3. Detection of Avian Bornavirus RNA in Field Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kohno, T.; Goto, T.; Takasaki, T.; Morita, C.; Nakaya, T.; Ikuta, K.; Kurane, I.; Sano, K.; Nakai, M. Fine Structure and Morphogenesis of Borna Disease Virus. J. Virol. 1999, 73, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Briese, T.; de la Torre, J.C.; Lewis, A.; Ludwig, H.; Lipkin, W.I. Borna disease virus, a negative-strand RNA virus, transcribes in the nucleus of infected cells. Proc. Natl. Acad. Sci. USA 1992, 89, 11486–11489. [Google Scholar] [CrossRef]
- Honkavuori, K.S.; Shivaprasad, H.; Williams, B.L.; Quan, P.-L.; Hornig, M.; Street, C.; Palacios, G.; Hutchison, S.K.; Franca, M.; Egholm, M.; et al. Novel Borna Virus in Psittacine Birds with Proventricular Dilatation Disease. Emerg. Infect. Dis. 2008, 14, 1883–1886. [Google Scholar] [CrossRef] [PubMed]
- Kistler, A.L.; Gancz, A.; Clubb, S.; Skewes-Cox, P.; Fischer, K.; Sorber, K.; Chiu, C.Y.; Lublin, A.; Mechani, S.; Farnoushi, Y.; et al. Recovery of divergent avian bornaviruses from cases of proventricular dilatation disease: Identification of a candidate etiologic agent. Virol. J. 2008, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gancz, A.Y.; Clubb, S.; Shivaprasad, H. Advanced Diagnostic Approaches and Current Management of Proventricular Dilatation Disease. Veter. Clin. N. Am. Exot. Anim. Pract. 2010, 13, 471–494. [Google Scholar] [CrossRef]
- Gray, P.; Hoppes, S.; Suchodolski, P.; Mirhosseini, N.; Payne, S.; Villanueva, I.; Shivaprasad, H.; Honkavuori, K.S.; Briese, T.; Reddy, S.M.; et al. Use of Avian Bornavirus Isolates to Induce Proventricular Dilatation Disease in Conures. Emerg. Infect. Dis. 2010, 16, 473–479. [Google Scholar] [CrossRef]
- Hoppes, S.M.; Tizard, I.; Shivaprasad, H. Avian Bornavirus and Proventricular Dilatation Disease. Veter. Clin. N. Am. Exot. Anim. Pract. 2013, 16, 339–355. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Dürrwald, R.; Bào, Y.; Briese, T.; Carbone, K.; Clawson, A.N.; DeRisi, J.L.; Garten, W.; Jahrling, P.B.; Kolodziejek, J.; et al. Taxonomic reorganization of the family Bornaviridae. Arch. Virol. 2015, 160, 621–632. [Google Scholar] [CrossRef]
- Amarasinghe, G.K.; Ayllón, M.A.; Bào, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef]
- Delnatte, P.; Ojkic, D.; DeLay, J.; Campbell, D.; Crawshaw, G.; Smith, D.A. Pathology and diagnosis of avian bornavirus infection in wild Canada geese (Branta canadensis), trumpeter swans (Cygnus buccinator) and mute swans (Cygnus olor) in Canada: A retrospective study. Avian Pathol. 2013, 42, 114–128. [Google Scholar] [CrossRef]
- Guo, J.; Shivaprasad, H.L.; Rech, R.R.; Heatley, J.J.; Tizard, I.; Payne, S. Characterization of a new genotype of avian bornavirus from wild ducks. Virol. J. 2014, 11, 197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Thomsen, A.F.; Nielsen, J.B.; Hjulsager, C.K.; Chriél, M.; Smith, D.A.; Bertelsen, M.F. Aquatic Bird Bornavirus 1 in Wild Geese, Denmark. Emerg. Infect. Dis. 2015, 21, 2201–2203. [Google Scholar] [CrossRef]
- Payne, S.; Covaleda, L.; Jianhua, G.; Swafford, S.; Baroch, J.; Ferro, P.J.; Lupiani, B.; Heatley, J.; Tizard, I. Detection and Characterization of a Distinct Bornavirus Lineage from Healthy Canada Geese (Branta canadensis). J. Virol. 2011, 85, 12053–12056. [Google Scholar] [CrossRef][Green Version]
- Murray, M.; Guo, J.; Tizard, I.; Jennings, S.; Shivaprasad, H.L.; Payne, S.; Ellis, J.C.; Van Wettere, A.; O’Brien, K.M. Aquatic Bird Bornavirus-Associated Disease in Free-Living Canada Geese (Branta canadensis) in the Northeastern USA. J. Wildl. Dis. 2017, 53, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Schmidt, V.; Rinder, M.; Legler, M.; Twietmeyer, S.; Schwemmer, P.; Corman, V.M. Phylogenetic Analysis Supports Horizontal Transmission as a Driving Force of the Spread of Avian Bornaviruses. PLoS ONE 2016, 11, e0160936. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Tizard, I.; Baroch, J.; Shivaprasad, H.L.; Payne, S.L. Avian Bornaviruses in North American Gulls. J. Wildl. Dis. 2015, 51, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Johansson, M.; Montell, H.; Berg, A.L. Wild birds as a possible natural reservoir of Borna disease virus. Epidemiol. Infect. 2001, 127, 173–178. [Google Scholar] [CrossRef]
- Rubbenstroth, D.; Rinder, M.; Stein, M.; Höper, D.; Kaspers, B.; Brosinski, K.; Horie, M.; Schmidt, V.; Legler, M.; Korbel, R.; et al. Avian bornaviruses are widely distributed in canary birds (Serinus canaria f. domestica). Veter. Microbiol. 2013, 165, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Schmidt, V.; Rinder, M.; Legler, M.; Corman, V.M.; Staeheli, P. Discovery of a new avian bornavirus genotype in estrildid finches (Estrildidae) in Germany. Veter. Microbiol. 2014, 168, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Kistler, A.L.; Smith, J.M.; Greninger, A.; DeRisi, J.L.; Ganem, D. Analysis of Naturally Occurring Avian Bornavirus Infection and Transmission during an Outbreak of Proventricular Dilatation Disease among Captive Psittacine Birds. J. Virol. 2009, 84, 2176–2179. [Google Scholar] [CrossRef]
- Herzog, S.; Enderlein, D.; Heffels-Redmann, U.; Piepenbring, A.; Neumann, D.; Kaleta, E.F.; Müller, H.; Lierz, M.; Herden, C. Indirect Immunofluorescence Assay for Intra Vitam Diagnosis of Avian Bornavirus Infection in Psittacine Birds. J. Clin. Microbiol. 2010, 48, 2282–2284. [Google Scholar] [CrossRef] [PubMed]
- Kerski, A.; De Kloet, A.H.; De Kloet, S.R. Vertical Transmission of Avian Bornavirus in Psittaciformes: Avian Bornavirus RNA and Anti-Avian Bornavirus Antibodies in Eggs, Embryos, and Hatchlings Obtained from Infected Sun Conures (Aratinga solstitialis). Avian Dis. 2012, 56, 471–478. [Google Scholar] [CrossRef]
- Olbert, M.; Römer-Oberdörfer, A.; Herden, C.; Malberg, S.; Runge, S.; Staeheli, P.; Rubbenstroth, D. Viral vector vaccines expressing nucleoprotein and phosphoprotein genes of avian bornaviruses ameliorate homologous challenge infections in cockatiels and common canaries. Sci. Rep. 2016, 6, 36840. [Google Scholar] [CrossRef]
- Runge, S.; Olbert, M.; Herden, C.; Malberg, S.; Römer-Oberdörfer, A.; Staeheli, P.; Rubbenstroth, D. Viral vector vaccines protect cockatiels from inflammatory lesions after heterologous parrot bornavirus 2 challenge infection. Vaccine 2017, 35, 557–563. [Google Scholar] [CrossRef]
- Piepenbring, A.K.; Enderlein, D.; Herzog, S.; Al-Ibadi, B.; Heffels-Redmann, U.; Heckmann, J.; Lange-Herbst, H.; Herden, C.; Lierz, M. Parrot Bornavirus (PaBV)-2 isolate causes different disease patterns in cockatiels than PaBV-4. Avian Pathol. 2016, 45, 156–168. [Google Scholar] [CrossRef]
- Guo, J.; Payne, S.; Zhang, S.; Turner, D.; Tizard, I.; Suchodolski, P. Avian Bornaviruses: Diagnosis, Isolation, and Genotyping. Curr. Protoc. Microbiol. 2014, 34, 15I.1.1–15I.1.33. [Google Scholar] [CrossRef] [PubMed]
- Delnatte, P.; Nagy, E.; Ojkic, D.; Leishman, D.; Crawshaw, G.; Elias, K.; Smith, D.A. Avian bornavirus in free-ranging waterfowl: Prevalence of antibodies and cloacal shedding of viral rna. J. Wildl. Dis. 2014, 50, 512–523. [Google Scholar] [CrossRef]
- Schlottau, K.; Forth, L.; Angstwurm, K.; Höper, D.; Zecher, D.; Liesche, F.; Hoffmann, B.; Kegel, V.; Seehofer, D.; Platen, S.; et al. Fatal Encephalitic Borna Disease Virus 1 in Solid-Organ Transplant Recipients. New Engl. J. Med. 2018, 379, 1377–1379. [Google Scholar] [CrossRef]
- Weissenböck, H.; Bakonyi, T.; Sekulin, K.; Ehrensperger, F.; Doneley, R.J.; Dürrwald, R.; Hoop, R.; Erdélyi, K.; Gál, J.; Kolodziejek, J.; et al. Avian Bornaviruses in Psittacine Birds from Europe and Australia with Proventricular Dilatation Disease. Emerg. Infect. Dis. 2009, 15, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Rinder, M.; Kaspers, B.; Staeheli, P. Efficient isolation of avian bornaviruses (ABV) from naturally infected psittacine birds and identification of a new ABV genotype from a salmon-crested cockatoo (Cacatua moluccensis). Veter. Microbiol. 2012, 161, 36–42. [Google Scholar] [CrossRef]
- Kessler, S.; Heenemann, K.; Krause, T.; Twietmeyer, S.; Fuchs, J.; Lierz, M.; Corman, V.M.; Vahlenkamp, T.M.; Rubbenstroth, D. Monitoring of free-ranging and captive Psittacula populations in Western Europe for avian bornaviruses, circoviruses and polyomaviruses. Avian Pathol. 2019, 49, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega, Accurate Alignment of Very Large Numbers of Sequences. In Multiple Sequence Alignment Methods; Springer: Berlin/Heidelberg, Germany, 2013; Volume 1079, pp. 105–116. [Google Scholar]
- Raymaekers, M.; Smets, R.; Maes, B.; Cartuyvels, R. Checklist for optimization and validation of real-time PCR assays. J. Clin. Lab. Anal. 2009, 23, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.; Wakem, M.; Dijkman, G.; Alsarraj, M.; Nguyen, M. A practical approach to RT-qPCR—Publishing data that conform to the MIQE guidelines. Methods 2010, 50, S1–S5. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Virus | Detection Limit of Xcon | Comparative Analysis of RT-PCR Assays (Highest log10 Dilution Factor of Reference cDNA with pos. Result) b | |||
|---|---|---|---|---|---|
| (Copy Numbers per Reaction) a | Xcon | Ncon [4] | Mcon [4] | Mcon-W [24] | |
| PaBV-1 | 102 | 3.0 | 2.0 | 3.0 | 2.0 |
| PaBV-2 | 103 | 4.0 | 4.0 | 3.5 | 2.0 |
| PaBV-4 | 102 | 3.0 | 3.0 | 2.5 | 2.0 |
| PaBV-7 | 102 | 3.5 | 3.0 | 2.5 | 2.0 |
| CnBV-1 | 103 | 3.0 | 2.5 | 0.5 | 0.5 |
| CnBV-2 | 102 | 2.0 | 0.5 | 2.5 | 1.5 |
| CnBV-3 | 103 | 2.5 | 0.5 | 1.0 | 0.5 |
| EsBV-1 | 103 | 0.5 | 1.0 | 0.5 | nd |
| ABBV-1 | 104 | 0.5 | nd | nd | nd |
| BoDV-1 | 108 | nd | 2.0 | 0.5 | nd |
| BoDV-2 | nd | nd | 1.5 | 0.5 | nd |
| Parameter | Probe Target | BornaP_FAM | BornaC_Aby | BornaA_A674N | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PaBV-1 | PaBV-2 | PaBV-4 | PaBV-7 | CnBV-1 | CnBV-2 | CnBV-3 | EsBV-1 | ABBV-1 | ||
| r2 | singleplex | 0.993 | 0.993 | 0.992 | 0.990 | 0.980 | 0.995 | 0.992 | 0.990 | 0.993 |
| multiplex | 0.987 | 0.989 | 0.999 | 0.993 | 0.999 | 0.994 | 0.995 | 0.996 | 0.983 | |
| E | singleplex | 91.8% | 106.1% | 98.0% | 100.9% | 107.3% | 92.1% | 97.3% | 93.5% | 95.2% |
| multiplex | 94.1% | 104.1% | 107.4% | 100.3% | 91.3% | 104.8% | 107.6% | 99.9% | 90.9% | |
| Detection limit a | singleplex | 101 | 101 | 102 | 102 | 103 | 101 | 102 | 102 | 103 |
| multiplex | 101 | 101 | 102 | 102 | 103 | 101 | 102 | 102 | 103 | |
| Probe Target | PaBV-1 | PaBV-2 | PaBV-4 | PaBV-7 | CnBV-1 | CnBV-2 | CnBV-3 | EsBV-1 | ABBV-1 | |
|---|---|---|---|---|---|---|---|---|---|---|
| mean CT | st | 24.15 ±0.19 | 25.72 ±0.36 | 23.61 ±0.55 | 23.55 ±0.49 | 24.11 ±0.97 | 22.24 ±0.89 | 25.64 ±0.80 | 27.20 ±0.67 | 22.64 ±0.33 |
| mt | 24.04 ±0.15 | 29.36 ±0.34 | 25.26 ±0.18 | 21.99 ±0.20 | 28.19 ±0.68 | 23.52 ±0.84 | 25.49 ±0.91 | 24.45 ±0.87 | 23.72 ±1.03 | |
| p-value | 0.5912 | 0.0002 | 0.0155 | 0.0170 | 0.0409 | 0.2299 | 0.8026 | 0.0130 | 0.1035 | |
| Sample Origin Virus | Sample Number | multiBornaX | panBorna 7.2 | PaBVcon MD | PaBVcon PG | PaBV-2 P | PaBV-4 P | CnBV-2 P | ABBV-1 M | ABBVcon P | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Probe P | Probe C | Probe A | ||||||||||
| Psittaciformes | ||||||||||||
| PaBV-1 | 2 | 2 | 0 | 0 | 2 | 2 | 2 | 2 | 2 | - | - | - |
| PaBV-2 | 5 | 5 | 0 | 0 | 5 | 5 | 5 | 5 | 0 | - | - | - |
| PaBV-4 | 21 | 18 a | 0 | 0 | 16 | 20 | 19 | 19 | 21 | - | - | - |
| PaBV-4 & -7 b | 2 | 2 | 0 | 0 | 2 | 2 | 2 | 2 | 2 | - | - | - |
| PaBV-7 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | - | - | - |
| undetermined | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | - | - | - |
| negative | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | - | - |
| Passeriformes | ||||||||||||
| CnBV-1 | 14 | 0 | 14 | 0 | 14 | - | - | - | - | 11 | - | - |
| CnBV-2 | 14 | 0 | 14 | 0 | 14 | - | - | - | - | 14 | - | - |
| CnBV-3 | 2 | 0 | 2 | 0 | 2 | - | - | - | - | 2 | - | - |
| EsBV-1 | 2 | 0 | 2 | 0 | 2 | - | - | - | - | 2 | - | - |
| negative | 4 | 0 | 0 | 0 | 0 | - | - | - | - | 0 | - | - |
| aquatic birds | ||||||||||||
| ABBV-1 | 5 | 0 | 0 | 4 | 3 | - | - | - | - | - | 5 | 5 |
| negative | 4 | 0 | 0 | 0 | 0 | - | - | - | - | - | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sigrist, B.; Geers, J.; Albini, S.; Rubbenstroth, D.; Wolfrum, N. A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses. Viruses 2021, 13, 1358. https://doi.org/10.3390/v13071358
Sigrist B, Geers J, Albini S, Rubbenstroth D, Wolfrum N. A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses. Viruses. 2021; 13(7):1358. https://doi.org/10.3390/v13071358
Chicago/Turabian StyleSigrist, Brigitte, Jessica Geers, Sarah Albini, Dennis Rubbenstroth, and Nina Wolfrum. 2021. "A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses" Viruses 13, no. 7: 1358. https://doi.org/10.3390/v13071358
APA StyleSigrist, B., Geers, J., Albini, S., Rubbenstroth, D., & Wolfrum, N. (2021). A New Multiplex Real-Time RT-PCR for Simultaneous Detection and Differentiation of Avian Bornaviruses. Viruses, 13(7), 1358. https://doi.org/10.3390/v13071358