
Metagenomic Analysis of Marigold: Mixed Infection Including Two New Viruses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
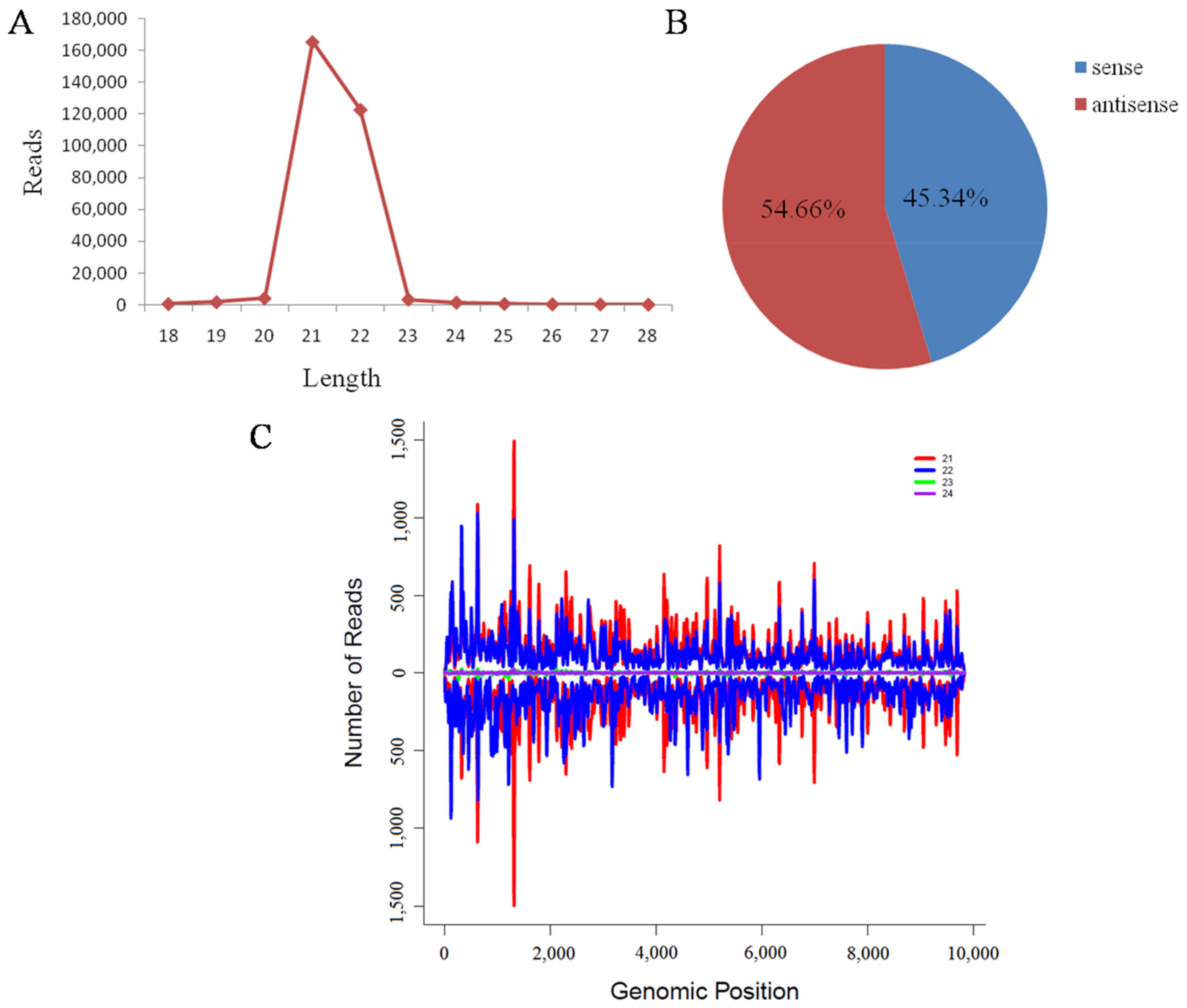
Abstract
Share and Cite
Yin, H.; Dong, Z.; Wang, X.; Lu, S.; Xia, F.; Abuduwaili, A.; Bi, Y.; Li, Y. Metagenomic Analysis of Marigold: Mixed Infection Including Two New Viruses. Viruses 2021, 13, 1254. https://doi.org/10.3390/v13071254
Yin H, Dong Z, Wang X, Lu S, Xia F, Abuduwaili A, Bi Y, Li Y. Metagenomic Analysis of Marigold: Mixed Infection Including Two New Viruses. Viruses. 2021; 13(7):1254. https://doi.org/10.3390/v13071254
Chicago/Turabian StyleYin, Hang, Zheng Dong, Xulong Wang, Shuhao Lu, Fei Xia, Annihaer Abuduwaili, Yang Bi, and Yongqiang Li. 2021. "Metagenomic Analysis of Marigold: Mixed Infection Including Two New Viruses" Viruses 13, no. 7: 1254. https://doi.org/10.3390/v13071254
APA StyleYin, H., Dong, Z., Wang, X., Lu, S., Xia, F., Abuduwaili, A., Bi, Y., & Li, Y. (2021). Metagenomic Analysis of Marigold: Mixed Infection Including Two New Viruses. Viruses, 13(7), 1254. https://doi.org/10.3390/v13071254