
Lipofection with Synthetic mRNA as a Simple Method for T-Cell Immunomonitoring

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Messenger RNA Preparation
2.2. Cell Culture and Luciferase Experiments
2.3. Measuring the Stimulation of the Innate Immune Response
2.4. Measuring the Stimulation of Adaptive Immunity in Murine Cells
2.5. Measuring the Stimulation of Adaptive Immunity in Human Cells
3. Results
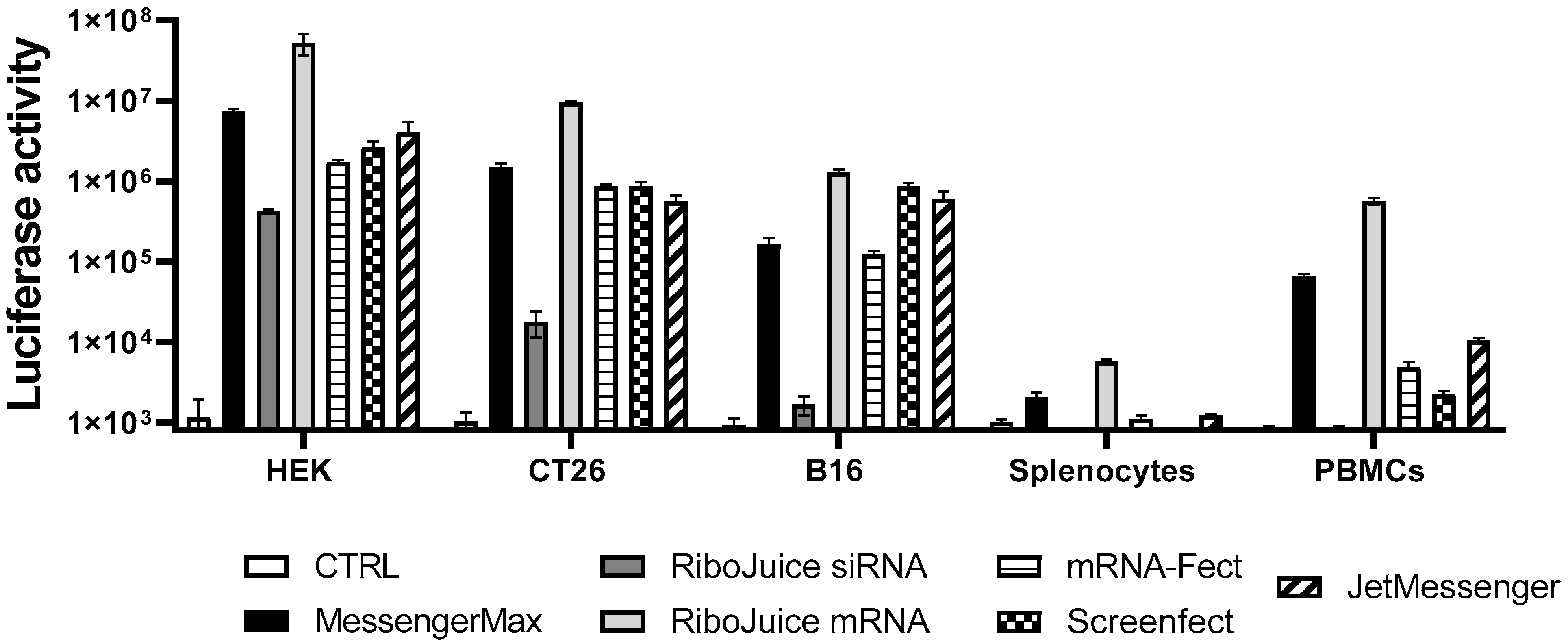
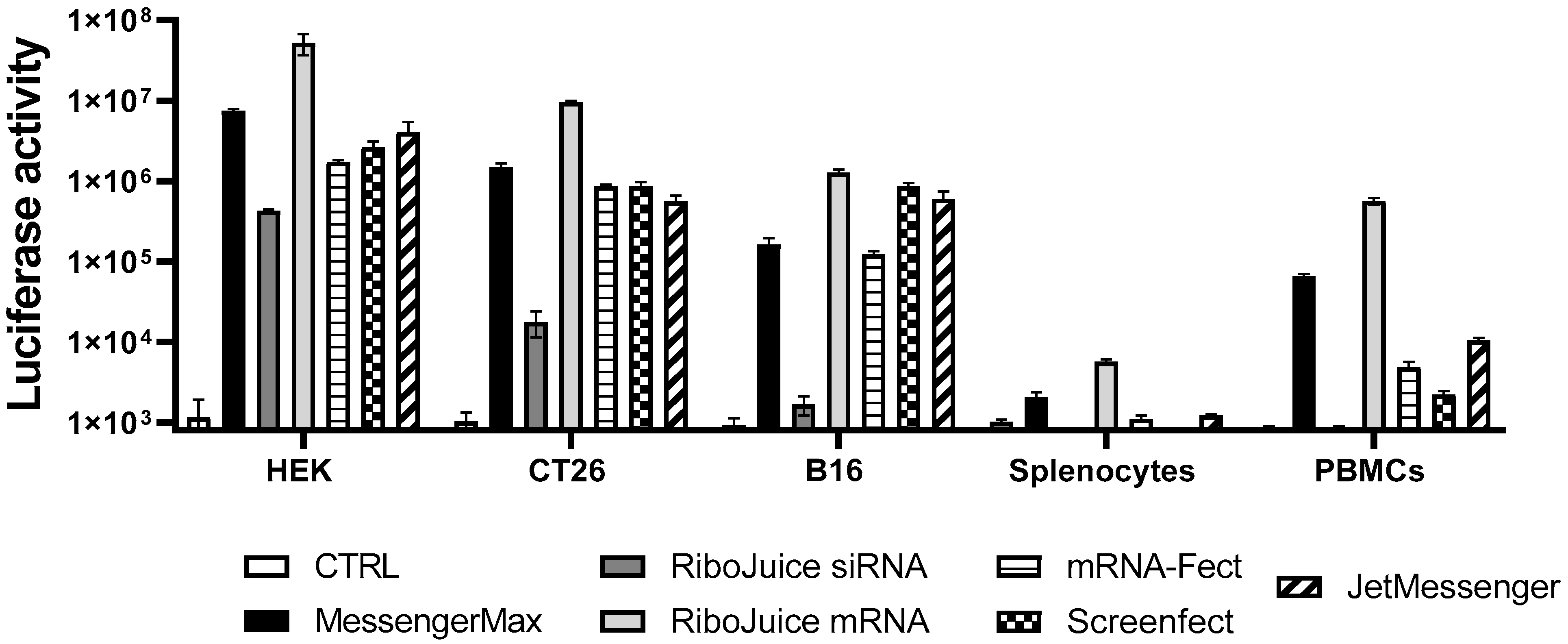
3.1. Screening of an Optimal Transfection Agent for Primary Immune Cells
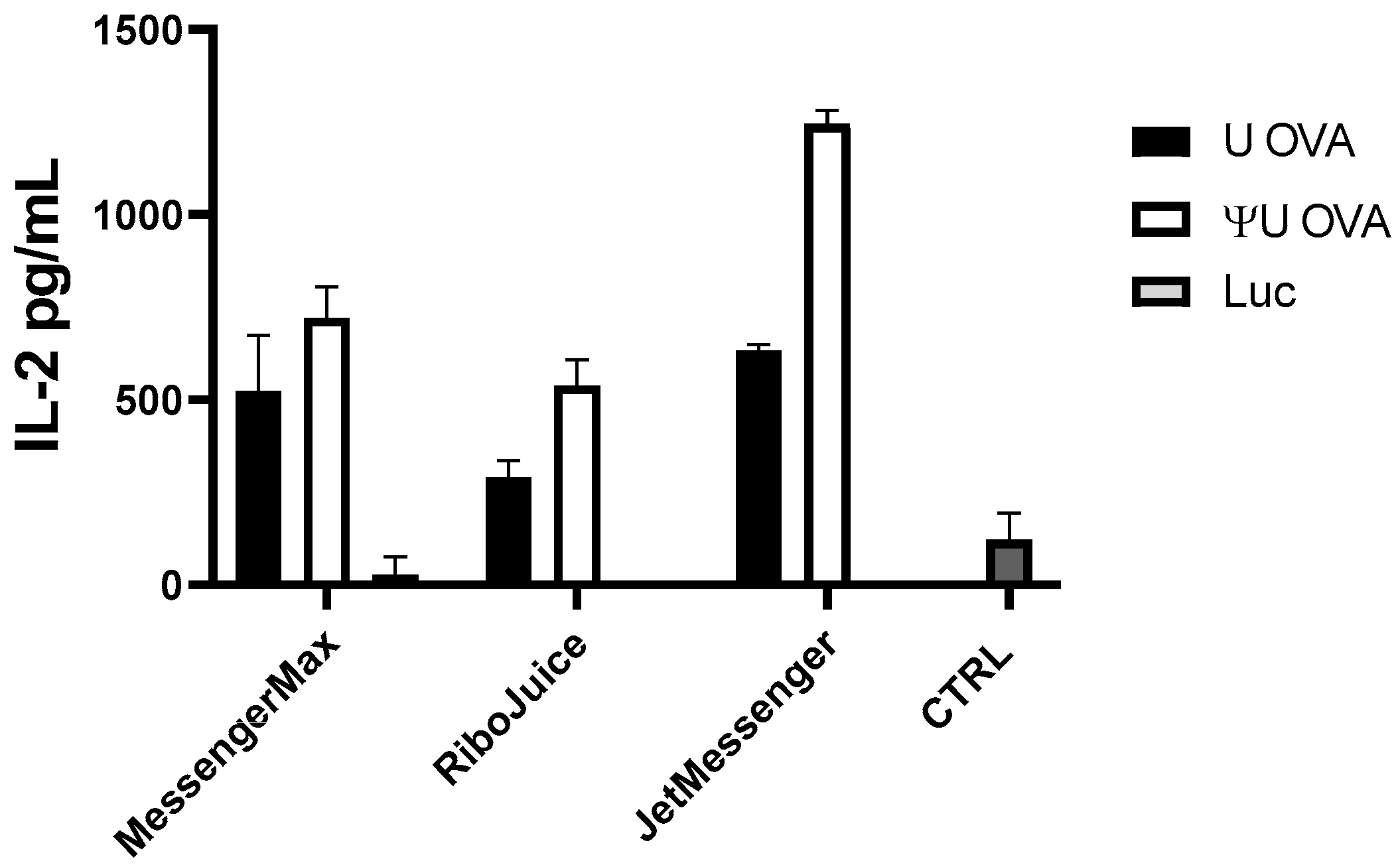
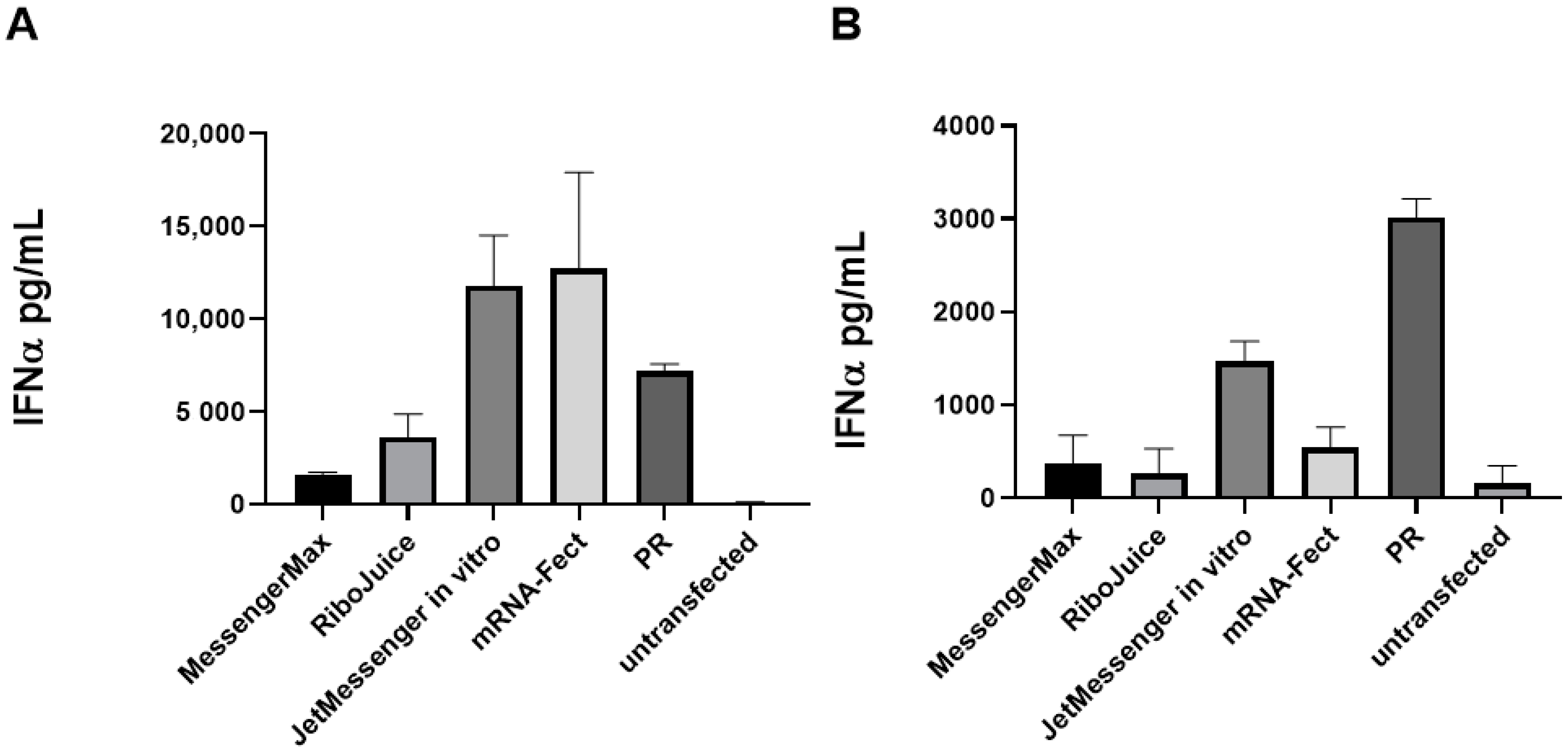
3.2. Effect of the Carrier on the Capacity of mRNA to Stimulate an Innate Immune Response
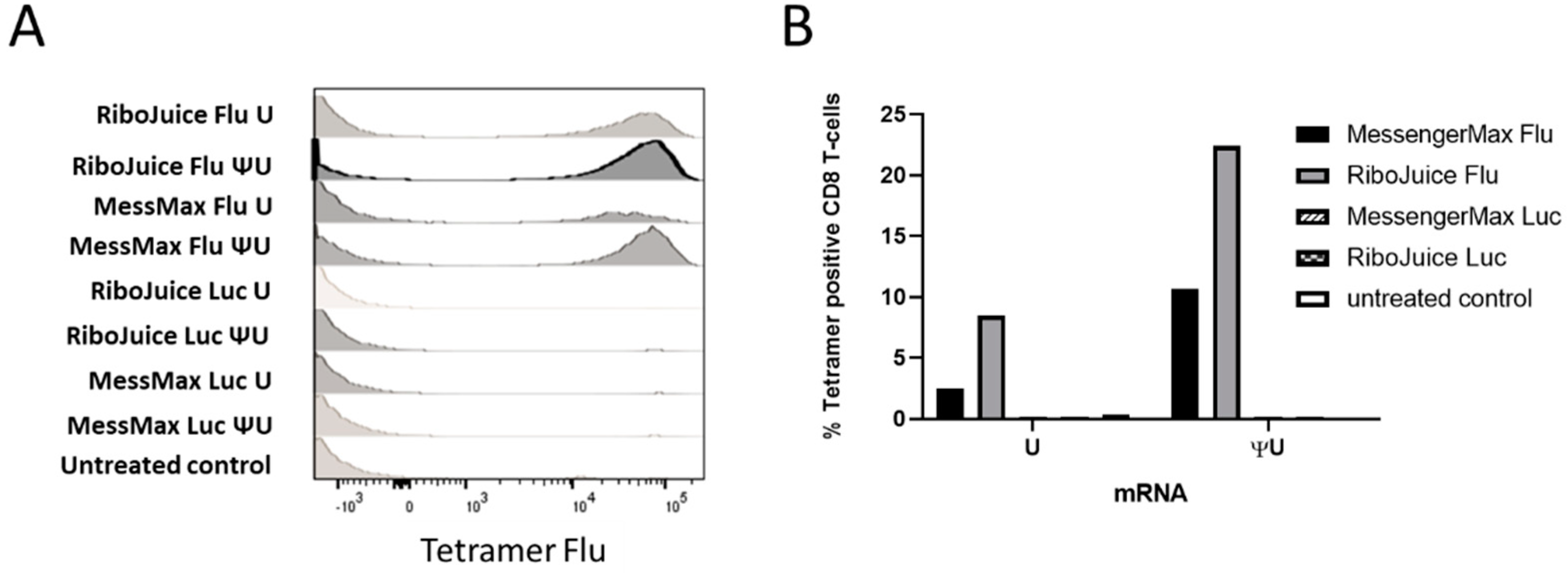
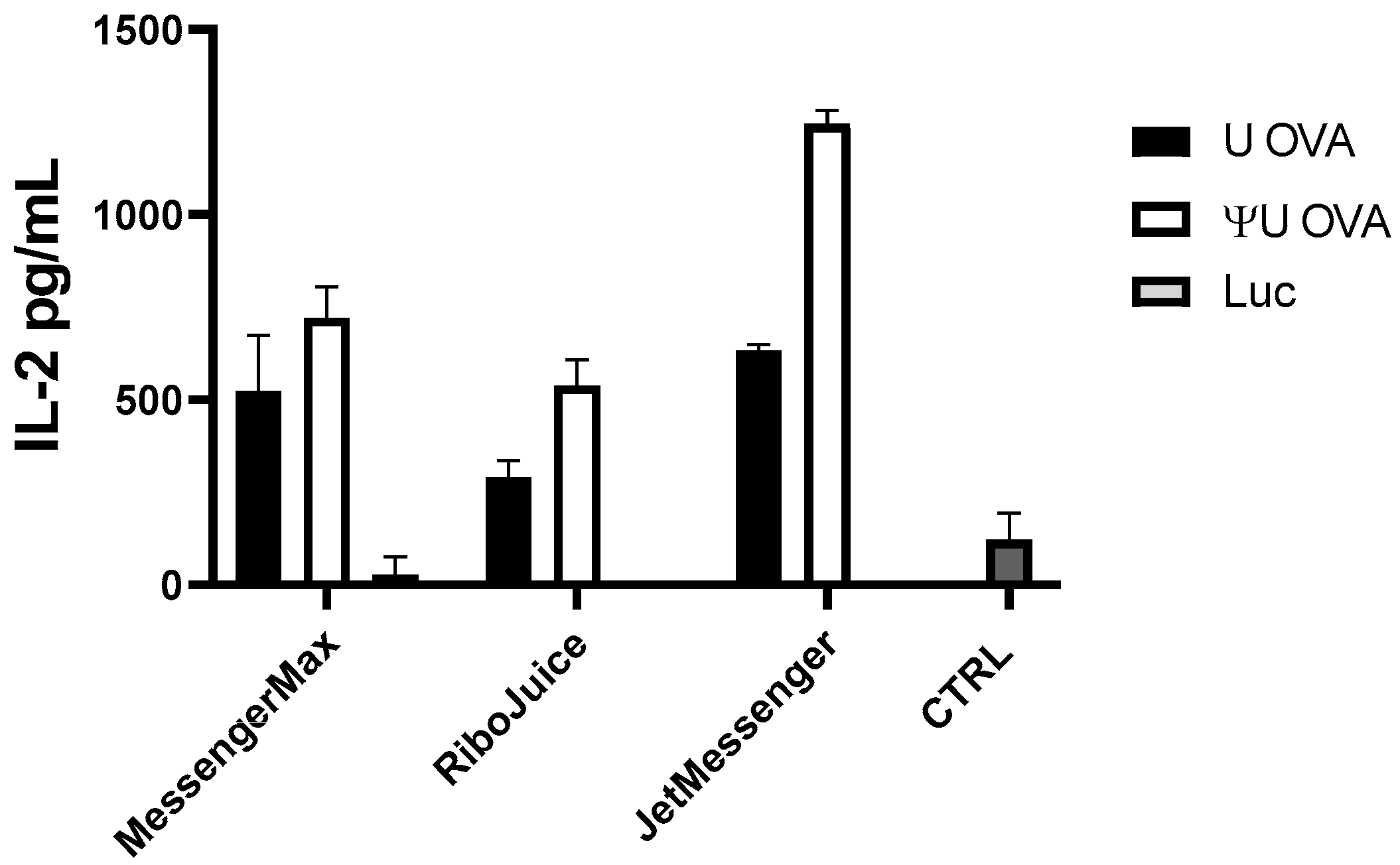
3.3. Optimising mRNA-Based T-Cell Immunomonitoring In Vitro in Murine Cells
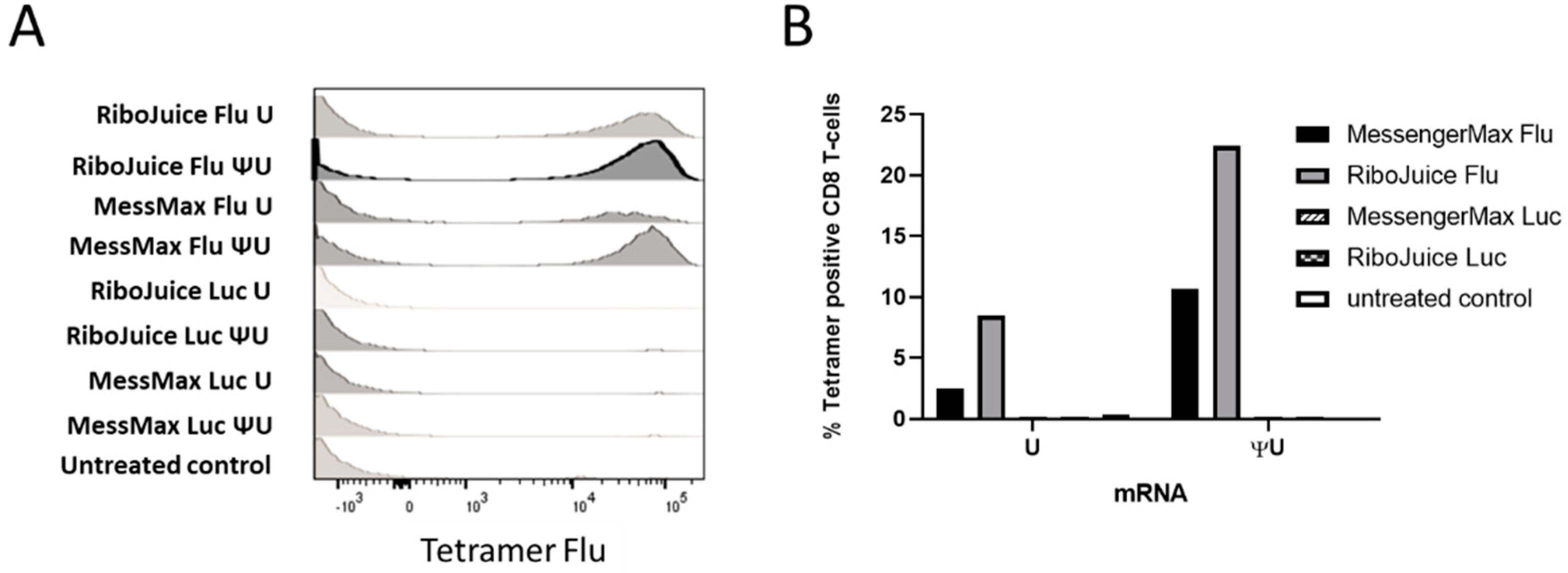
3.4. mRNA-Based In Vitro Immunomonitoring of Human T-Cells in Response to an Influenza Viral Protein
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duffy, D. Standardized immunomonitoring: Separating the signals from the noise. Trends Biotechnol. 2018, 36, 1107–1115. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, F.J.; Babdor, J.; Gherardini, P.F.; Amir, E.-A.D.; Jones, K.; Sahaf, B.; Marquez, D.M.; Krutzik, P.; O’Donnell, E.; Sigal, N.; et al. Comprehensive immune monitoring of clinical trials to advance human immunotherapy. Cell Rep. 2019, 28, 819–831.e4. [Google Scholar] [CrossRef] [Green Version]
- Teufel, R.; Carralot, J.P.; Scheel, B.; Probst, J.; Walter, S.; Jung, G.; Hoerr, I.; Rammensee, H.G.; Pascolo, S. Human peripheral blood mononuclear cells transfected with messenger rna stimulate antigen-specific cytotoxic t-lymphocytes in vitro. Cell. Mol. Life Sci. CMLS 2005, 62, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Uenaka, A.; Wada, H.; Isobe, M.; Saika, T.; Tsuji, K.; Sato, E.; Sato, S.; Noguchi, Y.; Kawabata, R.; Yasuda, T.; et al. T cell immunomonitoring and tumor responses in patients immunized with a complex of cholesterol-bearing hydrophobized pullulan (chp) and ny-eso-1 protein. Cancer Immun. Arch. 2007, 7, 9. [Google Scholar]
- Medaglini, D.; Siegrist, C.-A. Immunomonitoring of human responses to the rvsv-zebov ebola vaccine. Curr. Opin. Virol. 2017, 23, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Knights, A.J.; Nuber, N.; Thomson, C.W.; de la Rosa, O.; Jäger, E.; Tiercy, J.M.; van den Broek, M.; Pascolo, S.; Knuth, A.; Zippelius, A. Modified tumour antigen-encoding mrna facilitates the analysis of naturally occurring and vaccine-induced cd4 and cd8 t cells in cancer patients. Cancer Immunol. Immunother. 2009, 58, 325–338. [Google Scholar] [CrossRef]
- Bins, A.D.; Jorritsma, A.; Wolkers, M.C.; Hung, C.-F.; Wu, T.C.; Schumacher, T.N.M.; Haanen, J.B.A.G. A rapid and potent DNA vaccination strategy defined by in vivo monitoring of antigen expression. Nat. Med. 2005, 11, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase i/ii study of covid-19 rna vaccine bnt162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef]
- Kreiter, S.; Selmi, A.; Diken, M.; Sebastian, M.; Osterloh, P.; Schild, H.; Huber, C.; Türeci, O.; Sahin, U. Increased antigen presentation efficiency by coupling antigens to mhc class i trafficking signals. J. Immunol. 2008, 180, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Camp, K.; Cools, N.; Stein, B.; Van de Velde, A.; Goossens, H.; Berneman, Z.N.; Van Tendeloo, V. Efficient mrna electroporation of peripheral blood mononuclear cells to detect memory t cell responses for immunomonitoring purposes. J. Immunol. Methods 2010, 354, 1–10. [Google Scholar] [CrossRef]
- Hiura, M.; Ueno, K.; Suehiro, Y.; Hazama, S.; Oka, M.; Imai, K.; Hinoda, Y. A simple immunomonitoring procedure for mrna-loaded dendritic cell therapy. Tumor Biol. 2007, 28, 350–357. [Google Scholar] [CrossRef]
- Tusup, M.; Kundig, T.; Pascolo, S. An eif4g-recruiting aptamer increases the functionality of in vitro transcribed mrna. EPH Int. J. Med. Health Sci. 2018, 4, 29–35. [Google Scholar]
- Danckwardt, S.; Hentze, M.W.; Kulozik, A.E. 3′ end mrna processing: Molecular mechanisms and implications for health and disease. EMBO J. 2008, 27, 482–498. [Google Scholar] [CrossRef] [Green Version]
- Tusup, M.; Pascolo, S. Generation of immunostimulating 130 nm protamine–rna nanoparticles. In Rna Vaccines: Methods and Protocols; Kramps, T., Elbers, K., Eds.; Springer: New York, NY, USA, 2017; pp. 155–163. [Google Scholar]
- Jarzebska, N.T.; Lauchli, S.; Iselin, C.; French, L.E.; Johansen, P.; Guenova, E.; Kündig, T.M.; Pascolo, S. Functional differences between protamine preparations for the transfection of mrna. Drug Deliv. 2020, 27, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Gotch, F.; Rothbard, J.; Howland, K.; Townsend, A.; McMichael, A. Cytotoxic t lymphocytes recognize a fragment of influenza virus matrix protein in association with hla-a2. Nature 1987, 326, 881–882. [Google Scholar] [CrossRef]
- Moradian, H.; Roch, T.; Lendlein, A.; Gossen, M. Mrna transfection-induced activation of primary human monocytes and macrophages: Dependence on carrier system and nucleotide modification. Sci. Rep. 2020, 10, 4181. [Google Scholar] [CrossRef] [Green Version]
- Scheel, B.; Braedel, S.; Probst, J.; Carralot, J.-P.; Wagner, H.; Schild, H.; Jung, G.; Rammensee, H.-G.; Pascolo, S. Immunostimulating capacities of stabilized rna molecules. Eur. J. Immunol. 2004, 34, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Scheel, B.; Aulwurm, S.; Probst, J.; Stitz, L.; Hoerr, I.; Rammensee, H.-G.; Weller, M.; Pascolo, S. Therapeutic anti-tumor immunity triggered by injections of immunostimulating single-stranded rna. Eur. J. Immunol. 2006, 36, 2807–2816. [Google Scholar] [CrossRef]
- Jarrossay, D.; Napolitani, G.; Colonna, M.; Sallusto, F.; Lanzavecchia, A. Specialization and complementarity in microbial molecule recognition by human myeloid and plasmacytoid dendritic cells. Eur. J. Immunol 2001, 31, 3388–3393. [Google Scholar] [CrossRef]
- Erickson, S.; Sangfelt, O.; Castro, J.; Heyman, M.; Einhorn, S.; Grandér, D. Interferon-alpha inhibits proliferation in human t lymphocytes by abrogation of interleukin 2-induced changes in cell cycle-regulatory proteins. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1999, 10, 575–582. [Google Scholar]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of rna recognition by toll-like receptors: The impact of nucleoside modification and the evolutionary origin of rna. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Clarke, S.R.; Barnden, M.; Kurts, C.; Carbone, F.R.; Miller, J.F.; Heath, W.R. Characterization of the ovalbumin-specific tcr transgenic line ot-i: Mhc elements for positive and negative selection. Immunol. Cell Biol. 2000, 78, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Marrack, P.; Kappler, J.; Mitchell, T. Type i interferons keep activated t cells alive. J. Exp. Med. 1999, 189, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Dondi, E.; Rogge, L.; Lutfalla, G.; Uzé, G.; Pellegrini, S. Down-modulation of responses to type i ifn upon t cell activation. J. Immunol. 2003, 170, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of pseudouridine into mrna yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded rna via toll-like receptor 7 and 8. Science 2004, 303, 1526. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.R.; Muramatsu, H.; Nallagatla, S.R.; Bevilacqua, P.C.; Sansing, L.H.; Weissman, D.; Karikó, K. Incorporation of pseudouridine into mrna enhances translation by diminishing pkr activation. Nucleic Acids Res. 2010, 38, 5884–5892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, R.; Ho, N.W.; Gilham, P.T. Selective chemical modifications of uridine and pseudouridine in polynucleotides and their effect on the specificities of ribonuclease and phosphodiesterases. J. Am. Chem. Soc. 1965, 87, 4209–4210. [Google Scholar] [CrossRef]
- Davis, D.R. Stabilization of rna stacking by pseudouridine. Nucleic Acids Res 1995, 23, 5020–5026. [Google Scholar] [CrossRef] [PubMed]
- Macchia, I.; La Sorsa, V.; Ruspantini, I.; Sanchez, M.; Tirelli, V.; Carollo, M.; Fedele, G.; Leone, P.; Schiavoni, G.; Buccione, C.; et al. Multicentre harmonisation of a six-colour flow cytometry panel for naïve/memory t cell immunomonitoring. J. Immunol. Res. 2020, 2020, 1938704. [Google Scholar] [CrossRef] [Green Version]
- Brodin, P.; Davis, M.M. Human immune system variation. Nat. Rev. Immunol. 2017, 17, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, L.; Pekkarinen, P.T.; Lakshmikanth, T.; Tan, Z.; Consiglio, C.R.; Pou, C.; Chen, Y.; Mugabo, C.H.; Nguyen, N.A.; Nowlan, K.; et al. Systems-level immunomonitoring from acute to recovery phase of severe covid-19. Cell Rep. Med. 2020, 1, 100078. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Table | Compound | Transfection Efficiency | ||||
|---|---|---|---|---|---|---|
| HEK | CT26 | B16F10 | Murine Splenocytes | hPBMCs | ||
| Messenger Max | Liposome | ++ | ++ | + | ++ | ++ |
| RiboJuice mRNA | cationic polymer/lipid mixture | +++ | +++ | +++ | +++ | +++ |
| RiboJuice siRNA | cationic polymer/lipid mixture | + | +/− | − | − | − |
| mRNA-Fect | amphiphilic polymer | + | ++ | + | + | + |
| Screenfect | Liposome | ++ | ++ | ++ | − | +/− |
| JetMessenger | cationic polymer | ++ | ++ | ++ | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarzebska, N.T.; Frei, J.; Lauchli, S.; French, L.E.; Guenova, E.; Gouttefangeas, C.; Kündig, T.M.; Mellett, M.; Pascolo, S. Lipofection with Synthetic mRNA as a Simple Method for T-Cell Immunomonitoring. Viruses 2021, 13, 1232. https://doi.org/10.3390/v13071232
Jarzebska NT, Frei J, Lauchli S, French LE, Guenova E, Gouttefangeas C, Kündig TM, Mellett M, Pascolo S. Lipofection with Synthetic mRNA as a Simple Method for T-Cell Immunomonitoring. Viruses. 2021; 13(7):1232. https://doi.org/10.3390/v13071232
Chicago/Turabian StyleJarzebska, Natalia Teresa, Julia Frei, Severin Lauchli, Lars E. French, Emmanuella Guenova, Cécile Gouttefangeas, Thomas M. Kündig, Mark Mellett, and Steve Pascolo. 2021. "Lipofection with Synthetic mRNA as a Simple Method for T-Cell Immunomonitoring" Viruses 13, no. 7: 1232. https://doi.org/10.3390/v13071232
APA StyleJarzebska, N. T., Frei, J., Lauchli, S., French, L. E., Guenova, E., Gouttefangeas, C., Kündig, T. M., Mellett, M., & Pascolo, S. (2021). Lipofection with Synthetic mRNA as a Simple Method for T-Cell Immunomonitoring. Viruses, 13(7), 1232. https://doi.org/10.3390/v13071232