
Compelling Evidence for the Activity of Antiviral Peptides against SARS-CoV-2


Abstract
1. Introduction
1.1. SARS-CoV-2 and the Role of Antiviral Therapy Including AMPs
1.2. Defensins as Antiviral AMPs

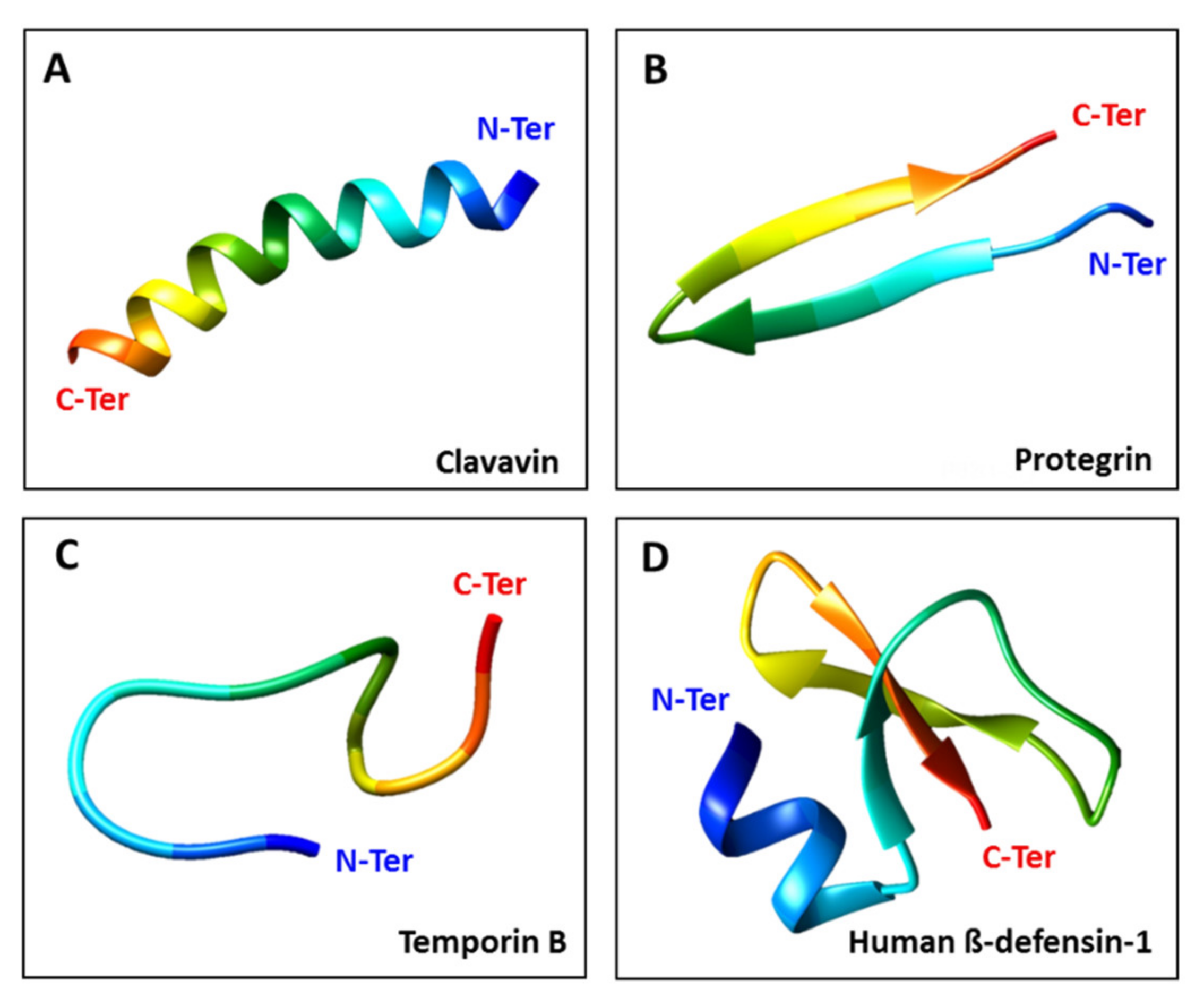{kind=link}
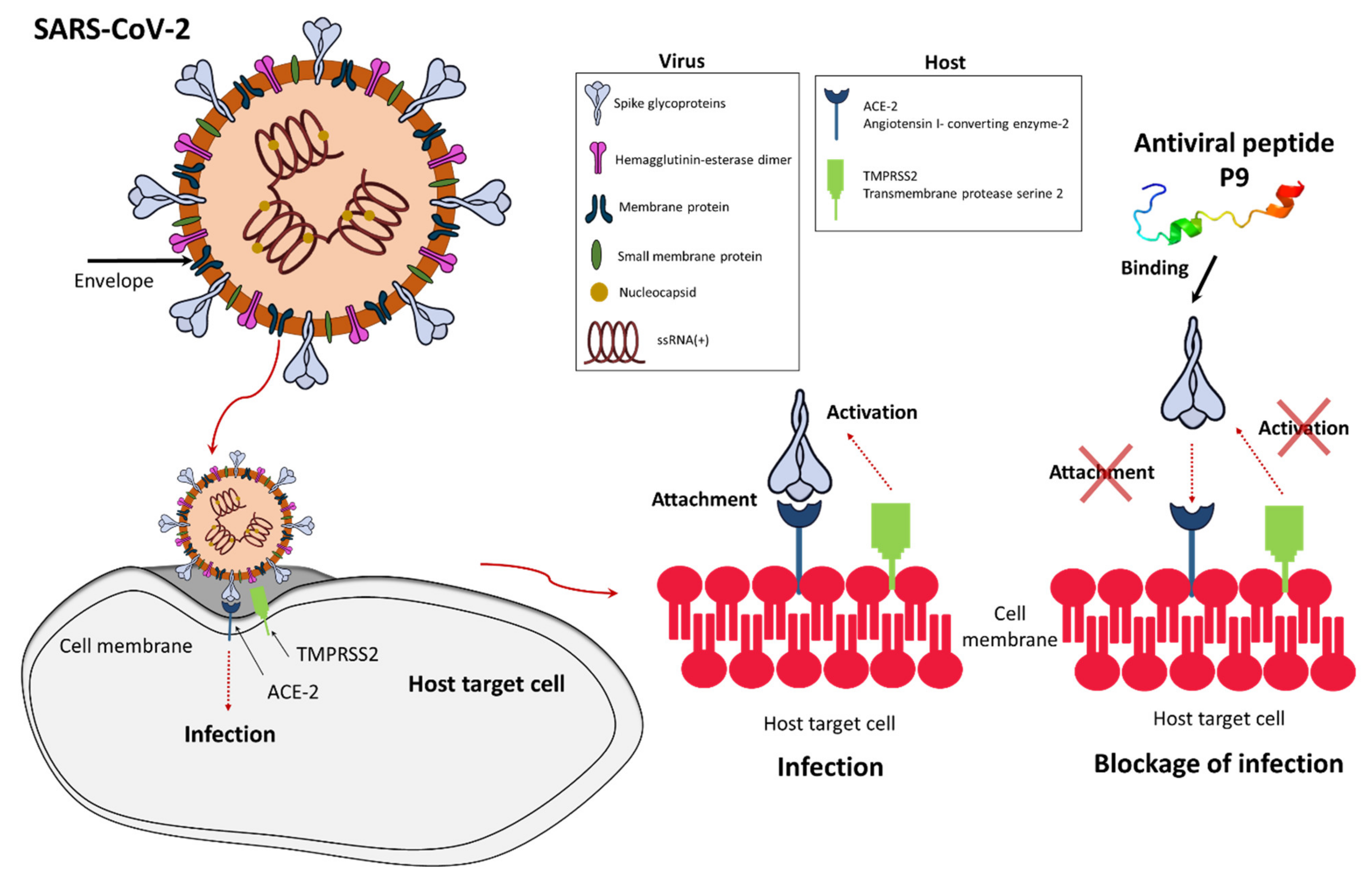{kind=link}
| Antiviral Peptides | Source | Group | Sequence | Activity | Type | Mechanism of Action | Reference |
|---|---|---|---|---|---|---|---|
| P9R (β-defensin-4 derivative) | Mus musculus | Mouse | NGAICWGPCPTAFRQIGNCGRFRVRCCRIR | SARS-CoV-2, MERS-CoV, SARS-CoV, A(H1N1)pdm09, A(H7N9) virus, and the non-enveloped rhinovirus | RNA, Enveloped/Non-enveloped | Inhibits viral replication by binding to different viruses and then inhibits virus–host endosomal acidification to prevent the endosomal release of pH-dependent viruses | [44] |
| P9 (β-defensin-4) | Mus musculus | Mouse | NGAICWGPCPTAFRQIGNCGHFKVRCCKIR | SARS-CoV, MERS-CoV, IAV (H1N1, H3N2, H5N1, H7N7, H7N9) | RNA, Enveloped | High-affinity binding to viral glycoproteins | [54] |
| θ-defensin retrocylin 2 | Synthetic construct | ND | GICRCICGRRICRCICGR | HIV, HSV, Influenza virus, Sindbis virus, Baculovirus | RNA, Enveloped/DNA virus | Inhibits viral fusion and entry by crosslinking membrane glycoproteins | [36,57] |
| MP7-NH2 (Mastoparan derivative) | Vespula lewisii | Insect | INLKALAALAKALL | VSV | RNA, Enveloped | Interact with the lipid component of virus membranes and thereby reduce infectivity of enveloped viruses | [58] |
| HEdefensin | Haemaphysalis longicornis | Tick | EEESEVAHLRVRRGFGCPLNQGACHRHCRSIRRRGGYCSGIIKQTCTCYRN | LGTV | RNA, Enveloped | Target virus membrane | [59] |
| longicin P4 | Haemaphysalis longicornis | Tick | SIGRRGGYCAGIIKQTCTCYR | TBEV surrogate LGTV | RNA, Enveloped | ND | [60] |
| Mucroporin-M1 | Lychas mucronatus | Scorpion | LFRLIKSLIKRLVSAFK | SARS-CoV, H5N1, Measles virus | RNA, Enveloped | Virus envelope interaction | [48] |
| Ev37 | Euscorpiops validus | Scorpion | GLINEKKVQQYLDEKLPNGVVKGALKSLVHKAAKNQNLCAFNVDTVGMCDADCKRQGKAKGVCHGTKCKCDVELSYKK | DENV-2, HCV, ZIKV, HSV-1 | RNA, Enveloped | Blocks the release of the viral genome from the endosome to the cytoplasm and then restricts viral late entry | [61] |
| BanLec | Musa acuminata | Plant | NGAIKVGAWGGNGGSAFDMG | HIV, HCP, Influenza virus | RNA, Enveloped | ND | [62] |
| Circulin-A | Chassalia parviflora | Plant | GIPCGESCVWIPCISAALGCSCKNKVCYRN | HIV | RNA, Enveloped | ND | [63] |
| Coccinin | Phaseolus coccineus | Plant | KQTENLADTY | HIV | RNA, Enveloped | HIV-1 reverse transcriptase inhibitory | [64] |
| Cycloviolins | Leonia cymosa | Plant | GVIPCGESCVFIPCISAAIGCSCKNKVCYRN | HIV | RNA, Enveloped | ND | [65] |
| Maculatin-1.1 | Litoria genimaculata | Frog | GLFGVLAKVAAHVVPAIAEHF | HIV | RNA, Enveloped | ND | [66] |
| Caerin-1.1 | Litoria splendida | Frog | GLLSVLGSVAKHVLPHVVPVIAEHL | HIV | RNA, Enveloped | ND | [67] |
| Urumin | Hydrophylax bahuvistara | Frog | IPLRGAFINGRWDSQCHRFSNGAIACA | H1 hemagglutinin-bearing human IAV | RNA, Enveloped | Destroys virions, targets the hemagglutinin stalk region | [68] |
| Tachyplesin I | Tachypleus tridentatus | Horseshoe crab | KWCFRVCYRGICYRRCR | VSV, IAV H1N1 | RNA, Enveloped | Inactivates the VSV by destroying its envelope subunits | [69] |
| Cyanovirin-N | Nostoc ellipsosporum | Cyanobacterium | LGKFSQTCYNSAIQGSVLTSTCERTNGGYNTSSIDLNSVIENVDGSLKWQPSNFIETCRNTQLAGSSELAAECKTRAQQFVSTKINLDDHIANIDGTLKYE | HIV-1 | RNA, Enveloped | Binds viral surface envelope glycoprotein gp120 | [70] |
1.3. Other Natural AMPs with Antiviral Properties
1.4. Advantages and Disadvantages of AMPs
2. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fu, L.; Wang, B.; Yuan, T.; Chen, X.; Ao, Y.; Fitzpatrick, T.; Li, P.; Zhou, Y.; Lin, Y.; Duan, Q.; et al. Clinical Characteristics of Coronavirus Disease 2019 (COVID-19) in China: A Systematic Review and Meta-Analysis. J. Infect. 2020, 80, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef]
- Khan, S.; Siddique, R.; Shereen, M.A.; Ali, A.; Liu, J.; Bai, Q.; Bashir, N.; Xue, M. Emergence of a Novel Coronavirus, Severe Acute Respiratory Syndrome Coronavirus 2: Biology and Therapeutic Options. J. Clin. Microbiol. 2020, 58. [Google Scholar] [CrossRef]
- WHO. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 30 March 2021).
- COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). Available online: https://coronavirus.jhu.edu/map.html (accessed on 30 March 2021).
- Su, L.; Ma, X.; Yu, H.; Zhang, Z.; Bian, P.; Han, Y.; Sun, J.; Liu, Y.; Yang, C.; Geng, J.; et al. The Different Clinical Characteristics of Corona Virus Disease Cases between Children and Their Families in China—The Character of Children with COVID-19. Emerg. Microbes Infect. 2020, 9, 707–713. [Google Scholar] [CrossRef]
- Li, Y.-D.; Chi, W.-Y.; Su, J.-H.; Ferrall, L.; Hung, C.-F.; Wu, T.-C. Coronavirus Vaccine Development: From SARS and MERS to COVID-19. J. Biomed. Sci. 2020, 27, 104. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, M.; Yin, L.; Wang, K.; Zhou, Y.; Zhou, M.; Lu, Y. COVID-19 Treatment: Close to a Cure? A Rapid Review of Pharmacotherapies for the Novel Coronavirus (SARS-CoV-2). Int. J. Antimicrob. Agents 2020, 56, 106080. [Google Scholar] [CrossRef] [PubMed]
- Eastman, R.T.; Roth, J.S.; Brimacombe, K.R.; Simeonov, A.; Shen, M.; Patnaik, S.; Hall, M.D. Remdesivir: A Review of Its Discovery and Development Leading to Emergency Use Authorization for Treatment of COVID-19. ACS Cent. Sci. 2020. [Google Scholar] [CrossRef]
- Khalili, J.S.; Zhu, H.; Mak, N.S.A.; Yan, Y.; Zhu, Y. Novel Coronavirus Treatment with Ribavirin: Groundwork for an Evaluation Concerning COVID-19. J. Med. Virol. 2020, 92, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Arabi, Y.M.; Chrousos, G.P.; Meduri, G.U. The Ten Reasons Why Corticosteroid Therapy Reduces Mortality in Severe COVID-19. Intensive Care Med. 2020, 46, 2067–2070. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, J.; Zhou, Y.; Zhao, X.; Zhao, Q.; Liu, J. The Effect of Corticosteroid Treatment on Patients with Coronavirus Infection: A Systematic Review and Meta-Analysis. J. Infect. 2020, 81, e13–e20. [Google Scholar] [CrossRef]
- Galeotti, C.; Kaveri, S.V.; Bayry, J. Intravenous Immunoglobulin Immunotherapy for Coronavirus Disease-19 (COVID-19). Clin. Transl. Immunol. 2020, 9, e1198. [Google Scholar] [CrossRef]
- Oren, O.; Yang, E.H.; Gluckman, T.J.; Michos, E.D.; Blumenthal, R.S.; Gersh, B.J. Use of Chloroquine and Hydroxychloroquine in COVID-19 and Cardiovascular Implications. Circ. Arrhythm. Electrophysiol. 2020, 13, e008688. [Google Scholar] [CrossRef]
- Guillén, L.; Padilla, S.; Fernández, M.; Agulló, V.; García, J.A.; Telenti, G.; García-Abellán, J.; Botella, Á.; Gutiérrez, F.; Masiá, M. Preemptive Interleukin-6 Blockade in Patients with COVID-19. Sci. Rep. 2020, 10, 16826. [Google Scholar] [CrossRef] [PubMed]
- Nasonov, E.; Samsonov, M. The Role of Interleukin 6 Inhibitors in Therapy of Severe COVID-19. Biomed. Pharmacother. 2020, 131, 110698. [Google Scholar] [CrossRef] [PubMed]
- Sinha, P.; Mostaghim, A.; Bielick, C.G.; McLaughlin, A.; Hamer, D.H.; Wetzler, L.M.; Bhadelia, N.; Fagan, M.A.; Linas, B.P.; Assoumou, S.A.; et al. Early Administration of Interleukin-6 Inhibitors for Patients with Severe COVID-19 Disease Is Associated with Decreased Intubation, Reduced Mortality, and Increased Discharge. Int. J. Infect. Dis. IJID 2020, 99, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wu, Z.; Li, J.-W.; Zhao, H.; Wang, G.-Q. Cytokine Release Syndrome in Severe COVID-19: Interleukin-6 Receptor Antagonist Tocilizumab May Be the Key to Reduce Mortality. Int. J. Antimicrob. Agents 2020, 55, 105954. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, J.P. Neutralizing Monoclonal Antibodies for COVID-19 Treatment and Prevention. Biomed. J. 2021, 44, 7–17. [Google Scholar] [CrossRef]
- Wagner, E.; Hewlett, M. WW|Retrovirus. In Basic Virology, 3rd ed.; Blackwell: Oxford, UK, 1999. [Google Scholar]
- Lou, Z.; Sun, Y.; Rao, Z. Current Progress in Antiviral Strategies. Trends Pharmacol. Sci. 2014, 35, 86–102. [Google Scholar] [CrossRef]
- Bakovic, A.; Risner, K.; Bhalla, N.; Alem, F.; Chang, T.L.; Weston, W.; Harness, J.A.; Narayanan, A. Brilacidin, a COVID-19 Drug Candidate, Exhibits Potent In Vitro Antiviral Activity Against SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef]
- Elnagdy, S.; AlKhazindar, M. The Potential of Antimicrobial Peptides as an Antiviral Therapy against COVID-19. ACS Pharmacol. Transl. Sci. 2020. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, W. Defensins in Innate Immunity. Curr. Opin. Hematol. 2014, 21, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-Microbial Peptides: From Invertebrates to Vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide Antimicrobial Agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The Antimicrobial Peptides and Their Potential Clinical Applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- De Vries, R.D.; Schmitz, K.S.; Bovier, F.T.; Predella, C.; Khao, J.; Noack, D.; Haagmans, B.L.; Herfst, S.; Stearns, K.N.; Drew-Bear, J.; et al. Intranasal Fusion Inhibitory Lipopeptide Prevents Direct-Contact SARS-CoV-2 Transmission in Ferrets. Science 2021, 371, 1379–1382. [Google Scholar] [CrossRef]
- Galdiero, S.; Falanga, A.; Tarallo, R.; Russo, L.; Galdiero, E.; Cantisani, M.; Morelli, G.; Galdiero, M. Peptide Inhibitors against Herpes Simplex Virus Infections. J. Pept. Sci. 2013, 19, 148–158. [Google Scholar] [CrossRef]
- Qureshi, A.; Thakur, N.; Kumar, M. VIRsiRNApred: A Web Server for Predicting Inhibition Efficacy of SiRNAs Targeting Human Viruses. J. Transl. Med. 2013, 11, 305. [Google Scholar] [CrossRef] [PubMed]
- Zapata, W.; Aguilar-Jiménez, W.; Feng, Z.; Weinberg, A.; Russo, A.; Potenza, N.; Estrada, H.; Rugeles, M.T. Identification of Innate Immune Antiretroviral Factors during in Vivo and in Vitro Exposure to HIV-1. Microbes Infect. 2016, 18, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.S.; Wiens, M.E.; Smith, J.G. Antiviral Mechanisms of Human Defensins. J. Mol. Biol. 2013, 425, 4965–4980. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, E.; Galen, B.; Lu, W.; Wang, W.; Ouyang, Y.; Keller, M.J.; Lehrer, R.I.; Herold, B.C. Human Alpha- and Beta-Defensins Block Multiple Steps in Herpes Simplex Virus Infection. J. Immunol. 2006, 177, 8658–8666. [Google Scholar] [CrossRef]
- Demirkhanyan, L.H.; Marin, M.; Padilla-Parra, S.; Zhan, C.; Miyauchi, K.; Jean-Baptiste, M.; Novitskiy, G.; Lu, W.; Melikyan, G.B. Multifaceted Mechanisms of HIV-1 Entry Inhibition by Human α-Defensin. J. Biol. Chem. 2012, 287, 28821–28838. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C. Describing the Mechanism of Antimicrobial Peptide Action with the Interfacial Activity Model. ACS Chem. Biol. 2010, 5, 905–917. [Google Scholar] [CrossRef] [PubMed]
- Leikina, E.; Delanoe-Ayari, H.; Melikov, K.; Cho, M.-S.; Chen, A.; Waring, A.J.; Wang, W.; Xie, Y.; Loo, J.A.; Lehrer, R.I.; et al. Carbohydrate-Binding Molecules Inhibit Viral Fusion and Entry by Crosslinking Membrane Glycoproteins. Nat. Immunol. 2005, 6, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E. Antivirals: Past, Present and Future. Biochem. Pharmacol. 2013, 85, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Ison, M.G. Antiviral Treatments. Clin. Chest Med. 2017, 38, 139–153. [Google Scholar] [CrossRef]
- Mulder, K.C.L.; Lima, L.A.; Miranda, V.J.; Dias, S.C.; Franco, O.L. Current Scenario of Peptide-Based Drugs: The Key Roles of Cationic Antitumor and Antiviral Peptides. Front. Microbiol. 2013, 4, 321. [Google Scholar] [CrossRef]
- Daher, K.A.; Selsted, M.E.; Lehrer, R.I. Direct Inactivation of Viruses by Human Granulocyte Defensins. J. Virol. 1986, 60, 1068–1074. [Google Scholar] [CrossRef]
- Gwyer Findlay, E.; Currie, S.M.; Davidson, D.J. Cationic Host Defence Peptides: Potential as Antiviral Therapeutics. BioDrugs Clin. Immunother. Biopharm. Gene Ther. 2013, 27, 479–493. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, D.; Li, W.; Wang, B.; Jiang, Z.; Li, M. Antiviral Activity of Recombinant Mouse β-Defensin 3 against Influenza A Virus in Vitro and in Vivo. Antivir. Chem. Chemother. 2012, 22, 255–262. [Google Scholar] [CrossRef]
- Park, M.S.; Kim, J.I.; Lee, I.; Park, S.; Bae, J.-Y.; Park, M.-S. Towards the Application of Human Defensins as Antivirals. Biomol. Ther. 2018, 26, 242–254. [Google Scholar] [CrossRef]
- Zhao, H.; To, K.K.W.; Sze, K.-H.; Yung, T.T.-M.; Bian, M.; Lam, H.; Yeung, M.L.; Li, C.; Chu, H.; Yuen, K.-Y. A Broad-Spectrum Virus- and Host-Targeting Peptide against Respiratory Viruses Including Influenza Virus and SARS-CoV-2. Nat. Commun. 2020, 11, 4252. [Google Scholar] [CrossRef]
- Zhao, H.; To, K.K.W.; Chu, H.; Ding, Q.; Zhao, X.; Li, C.; Shuai, H.; Yuan, S.; Zhou, J.; Kok, K.-H.; et al. Dual-Functional Peptide with Defective Interfering Genes Effectively Protects Mice against Avian and Seasonal Influenza. Nat. Commun. 2018, 9, 2358. [Google Scholar] [CrossRef]
- Yu, Y.; Deng, Y.-Q.; Zou, P.; Wang, Q.; Dai, Y.; Yu, F.; Du, L.; Zhang, N.-N.; Tian, M.; Hao, J.-N.; et al. A Peptide-Based Viral Inactivator Inhibits Zika Virus Infection in Pregnant Mice and Fetuses. Nat. Commun. 2017, 8, 15672. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Q.; Zhu, Y.; Chan, K.-H.; Qin, L.; Li, Y.; Wang, Q.; Chan, J.F.-W.; Du, L.; Yu, F.; et al. Structure-Based Discovery of Middle East Respiratory Syndrome Coronavirus Fusion Inhibitor. Nat. Commun. 2014, 5, 3067. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, Z.; Zhou, D.; Chen, Y.; Hong, W.; Cao, L.; Yang, J.; Zhang, Y.; Shi, W.; Cao, Z.; et al. Virucidal Activity of a Scorpion Venom Peptide Variant Mucroporin-M1 against Measles, SARS-CoV and Influenza H5N1 Viruses. Peptides 2011, 32, 1518–1525. [Google Scholar] [CrossRef]
- Liu, S.; Lu, H.; Niu, J.; Xu, Y.; Wu, S.; Jiang, S. Different from the HIV Fusion Inhibitor C34, the Anti-HIV Drug Fuzeon (T-20) Inhibits HIV-1 Entry by Targeting Multiple Sites in Gp41 and Gp120. J. Biol. Chem. 2005, 280, 11259–11273. [Google Scholar] [CrossRef]
- Gomes, B.; Augusto, M.T.; Felício, M.R.; Hollmann, A.; Franco, O.L.; Gonçalves, S.; Santos, N.C. Designing Improved Active Peptides for Therapeutic Approaches against Infectious Diseases. Biotechnol. Adv. 2018, 36, 415–429. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.-T.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A Pan-Coronavirus Fusion Inhibitor Targeting the HR1 Domain of Human Coronavirus Spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Zhu, Y.; Liu, M.; Lan, Q.; Xu, W.; Wu, Y.; Ying, T.; Liu, S.; Shi, Z.; Jiang, S.; et al. Fusion Mechanism of 2019-NCoV and Fusion Inhibitors Targeting HR1 Domain in Spike Protein. Cell. Mol. Immunol. 2020, 17, 765–767. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, J.; Zhang, K.; Chu, H.; Liu, D.; Poon, V.K.-M.; Chan, C.C.-S.; Leung, H.-C.; Fai, N.; Lin, Y.-P.; et al. A Novel Peptide with Potent and Broad-Spectrum Antiviral Activities against Multiple Respiratory Viruses. Sci. Rep. 2016, 6, 22008. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S.; Li, D.; Wei, D.-Q.; Zhao, J.; Wang, J. Human Intestinal Defensin 5 Inhibits SARS-CoV-2 Invasion by Cloaking ACE2. Gastroenterology 2020, 159, 1145–1147. [Google Scholar] [CrossRef] [PubMed]
- Mahendran, A.S.K.; Lim, Y.S.; Fang, C.-M.; Loh, H.-S.; Le, C.F. The Potential of Antiviral Peptides as COVID-19 Therapeutics. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef]
- Daly, N.L.; Chen, Y.-K.; Rosengren, K.J.; Marx, U.C.; Phillips, M.L.; Waring, A.J.; Wang, W.; Lehrer, R.I.; Craik, D.J. Retrocyclin-2: Structural Analysis of a Potent Anti-HIV Theta-Defensin. Biochemistry 2007, 46, 9920–9928. [Google Scholar] [CrossRef]
- Sample, C.J.; Hudak, K.E.; Barefoot, B.E.; Koci, M.D.; Wanyonyi, M.S.; Abraham, S.; Staats, H.F.; Ramsburg, E.A. A Mastoparan-Derived Peptide Has Broad-Spectrum Antiviral Activity against Enveloped Viruses. Peptides 2013, 48, 96–105. [Google Scholar] [CrossRef]
- Talactac, M.R.; Yada, Y.; Yoshii, K.; Hernandez, E.P.; Kusakisako, K.; Maeda, H.; Galay, R.L.; Fujisaki, K.; Mochizuki, M.; Tanaka, T. Characterization and Antiviral Activity of a Newly Identified Defensin-like Peptide, HEdefensin, in the Hard Tick Haemaphysalis Longicornis. Dev. Comp. Immunol. 2017, 68, 98–107. [Google Scholar] [CrossRef]
- Talactac, M.R.; Yoshii, K.; Maeda, H.; Kusakisako, K.; Hernandez, E.P.; Tsuji, N.; Fujisaki, K.; Galay, R.L.; Tanaka, T.; Mochizuki, M. Virucidal Activity of Haemaphysalis longicornis Longicin P4 Peptide against Tick-Borne Encephalitis Virus Surrogate Langat Virus. Parasit. Vectors 2016, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Lang, Y.; Ji, Z.; Xia, Z.; Han, Y.; Cheng, Y.; Liu, G.; Sun, F.; Zhao, Y.; Gao, M.; et al. A Scorpion Venom Peptide Ev37 Restricts Viral Late Entry by Alkalizing Acidic Organelles. J. Biol. Chem. 2019, 294, 182–194. [Google Scholar] [CrossRef]
- Swanson, M.D.; Boudreaux, D.M.; Salmon, L.; Chugh, J.; Winter, H.C.; Meagher, J.L.; André, S.; Murphy, P.V.; Oscarson, S.; Roy, R.; et al. Engineering a Therapeutic Lectin by Uncoupling Mitogenicity from Antiviral Activity. Cell 2015, 163, 746–758. [Google Scholar] [CrossRef]
- Nguyen, G.K.T.; Lim, W.H.; Nguyen, P.Q.T.; Tam, J.P. Novel Cyclotides and Uncyclotides with Highly Shortened Precursors from Chassalia chartacea and Effects of Methionine Oxidation on Bioactivities. J. Biol. Chem. 2012, 287, 17598–17607. [Google Scholar] [CrossRef]
- Ngai, P.H.K.; Ng, T.B. Coccinin, an Antifungal Peptide with Antiproliferative and HIV-1 Reverse Transcriptase Inhibitory Activities from Large Scarlet Runner Beans. Peptides 2004, 25, 2063–2068. [Google Scholar] [CrossRef]
- Hallock, Y.F.; Sowder, R.C.; Pannell, L.K.; Hughes, C.B.; Johnson, D.G.; Gulakowski, R.; Cardellina, J.H.; Boyd, M.R. Cycloviolins A–D, Anti-HIV Macrocyclic Peptides from Leonia cymosa. J. Org. Chem. 2000, 65, 124–128. [Google Scholar] [CrossRef]
- Rozek, T.; Waugh, R.J.; Steinborner, S.T.; Bowie, J.H.; Tyler, M.J.; Wallace, J.C. The Maculatin Peptides from the Skin Glands of the Tree Frog Litoria genimaculata: A Comparison of the Structures and Antibacterial Activities of Maculatin 1.1 and Caerin 1.1. J. Pept. Sci. 1998, 4, 111–115. [Google Scholar] [CrossRef]
- Stone, D.J.M.; Waugh, R.J.; Bowie, J.H.; Wallace, J.C.; Tyler, M.J. Peptides from Australian Frogs. Structures of the Caerins and Caeridin 1 from Litoria splendida. J. Chem. Soc. Perkin Trans. 1992, 3173–3178. [Google Scholar] [CrossRef]
- Holthausen, D.J.; Lee, S.H.; Kumar, V.T.; Bouvier, N.M.; Krammer, F.; Ellebedy, A.H.; Wrammert, J.; Lowen, A.C.; George, S.; Pillai, M.R.; et al. An Amphibian Host Defense Peptide Is Virucidal for Human H1 Hemagglutinin-Bearing Influenza Viruses. Immunity 2017, 46, 587–595. [Google Scholar] [CrossRef]
- Murakami, T.; Niwa, M.; Tokunaga, F.; Miyata, T.; Iwanaga, S. Direct Virus Inactivation of Tachyplesin I and Its Isopeptides from Horseshoe Crab Hemocytes. Chemotherapy 1991, 37, 327–334. [Google Scholar] [CrossRef]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of Cyanovirin-N, a Novel Human Immunodeficiency Virus-Inactivating Protein That Binds Viral Surface Envelope Glycoprotein Gp120: Potential Applications to Microbicide Development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.-J.; Nikapitiya, C.; Lee, H.-C.; Park, M.-E.; Kim, J.-H.; Kim, T.-H.; Yoon, J.-E.; Cho, W.-K.; Ma, J.Y.; Kim, C.-J.; et al. Inhibition of Highly Pathogenic Avian Influenza (HPAI) Virus by a Peptide Derived from VFLIP through Its Direct Destabilization of Viruses. Sci. Rep. 2017, 7, 4875. [Google Scholar] [CrossRef]
- Hussain, M.; Galvin, H.D.; Haw, T.Y.; Nutsford, A.N.; Husain, M. Drug Resistance in Influenza A Virus: The Epidemiology and Management. Infect. Drug Resist. 2017, 10, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Pillay, D.; Zambon, M. Antiviral Drug Resistance. BMJ 1998, 317, 660–662. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.Y.; Yang, J.-R. Analysis and Prediction of Highly Effective Antiviral Peptides Based on Random Forests. PLoS ONE 2013, 8, e70166. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.Y.; Murakami, E.; Zorca, S.M.; Johnson, A.A.; Johnson, K.A.; Schinazi, R.F.; Furman, P.A.; Anderson, K.S. Relationship between Antiviral Activity and Host Toxicity: Comparison of the Incorporation Efficiencies of 2′,3′-Dideoxy-5-Fluoro-3′-Thiacytidine-Triphosphate Analogs by Human Immunodeficiency Virus Type 1 Reverse Transcriptase and Human Mitochondrial DNA Polymerase. Antimicrob. Agents Chemother. 2004, 48, 1300–1306. [Google Scholar] [CrossRef] [PubMed]
- Carr, A. Toxicity of Antiretroviral Therapy and Implications for Drug Development. Nat. Rev. Drug Discov. 2003, 2, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Sala, A.; Ardizzoni, A.; Ciociola, T.; Magliani, W.; Conti, S.; Blasi, E.; Cermelli, C. Antiviral Activity of Synthetic Peptides Derived from Physiological Proteins. Intervirology 2018, 61, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, G.; Gabrani, R. Antiviral Peptides: Identification and Validation. Int. J. Pept. Res. Ther. 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhang, S.; Peng, J.; Li, Y.; Yang, Q. Synthetic Surfactin Analogues Have Improved Anti-PEDV Properties. PLoS ONE 2019, 14, e0215227. [Google Scholar] [CrossRef] [PubMed]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral Peptides as Promising Therapeutic Drugs. Cell. Mol. Life Sci. CMLS 2019, 76, 3525–3542. [Google Scholar] [CrossRef]
- Di, L. Strategic Approaches to Optimizing Peptide ADME Properties. AAPS J. 2015, 17, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Shartouny, J.R.; Jacob, J. Mining the Tree of Life: Host Defense Peptides as Antiviral Therapeutics. Semin. Cell Dev. Biol. 2019, 88, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Pattabiraman, V.R.; Bode, J.W. Rethinking Amide Bond Synthesis. Nature 2011, 480, 471–479. [Google Scholar] [CrossRef] [PubMed]
- McGregor, D.P. Discovering and Improving Novel Peptide Therapeutics. Curr. Opin. Pharmacol. 2008, 8, 616–619. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonk, M.; Růžek, D.; Vilcinskas, A. Compelling Evidence for the Activity of Antiviral Peptides against SARS-CoV-2. Viruses 2021, 13, 912. https://doi.org/10.3390/v13050912
Tonk M, Růžek D, Vilcinskas A. Compelling Evidence for the Activity of Antiviral Peptides against SARS-CoV-2. Viruses. 2021; 13(5):912. https://doi.org/10.3390/v13050912
Chicago/Turabian StyleTonk, Miray, Daniel Růžek, and Andreas Vilcinskas. 2021. "Compelling Evidence for the Activity of Antiviral Peptides against SARS-CoV-2" Viruses 13, no. 5: 912. https://doi.org/10.3390/v13050912
APA StyleTonk, M., Růžek, D., & Vilcinskas, A. (2021). Compelling Evidence for the Activity of Antiviral Peptides against SARS-CoV-2. Viruses, 13(5), 912. https://doi.org/10.3390/v13050912