Changes in MicroRNA Expression during Rabbit Hemorrhagic Disease Virus (RHDV) Infection

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Rabbit Hemorrhagic Disease Virus (RHDV)
2.2. Tissue Samples
2.3. Serum Samples
2.4. RNA Isolation from Tissue Samples
2.5. RNA Isolation from Serum Samples
2.6. miRNA Polyadenylation and Reverse Transcription (RT) Reaction
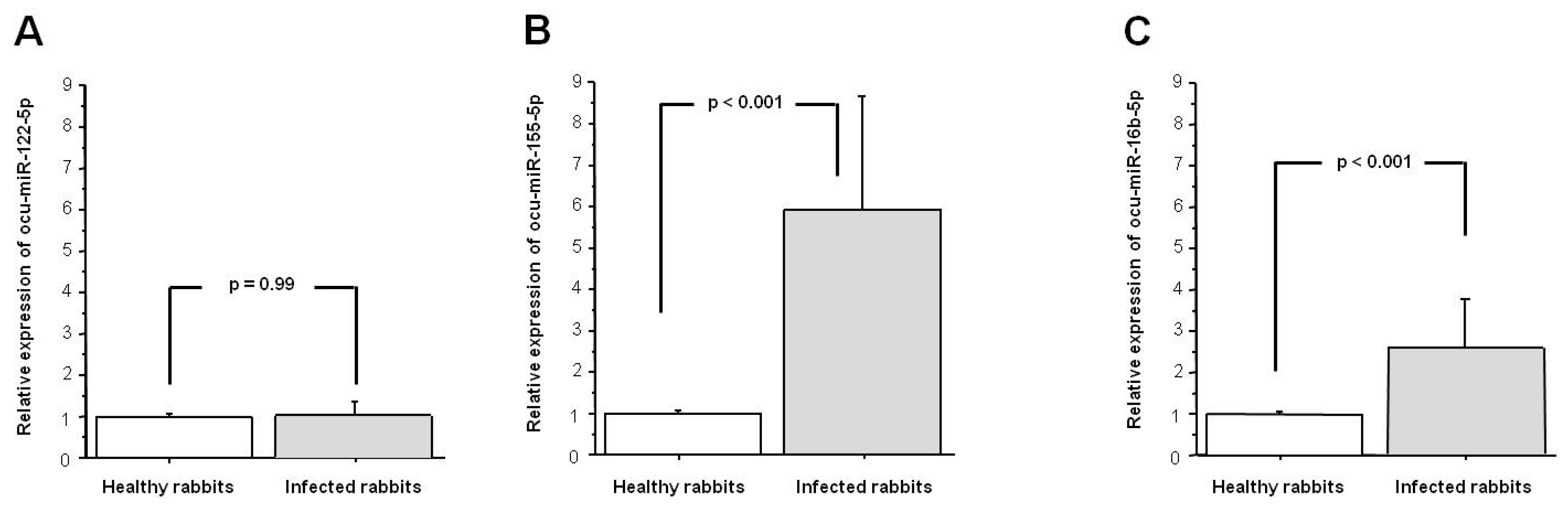
2.7. Quantification of ocu-miR-122-5p, ocu-miR-155-5p, and ocu-miR-16b-5p in Liver Samples Using Quantitative Real-Time PCR (qPCR) and Data Analysis
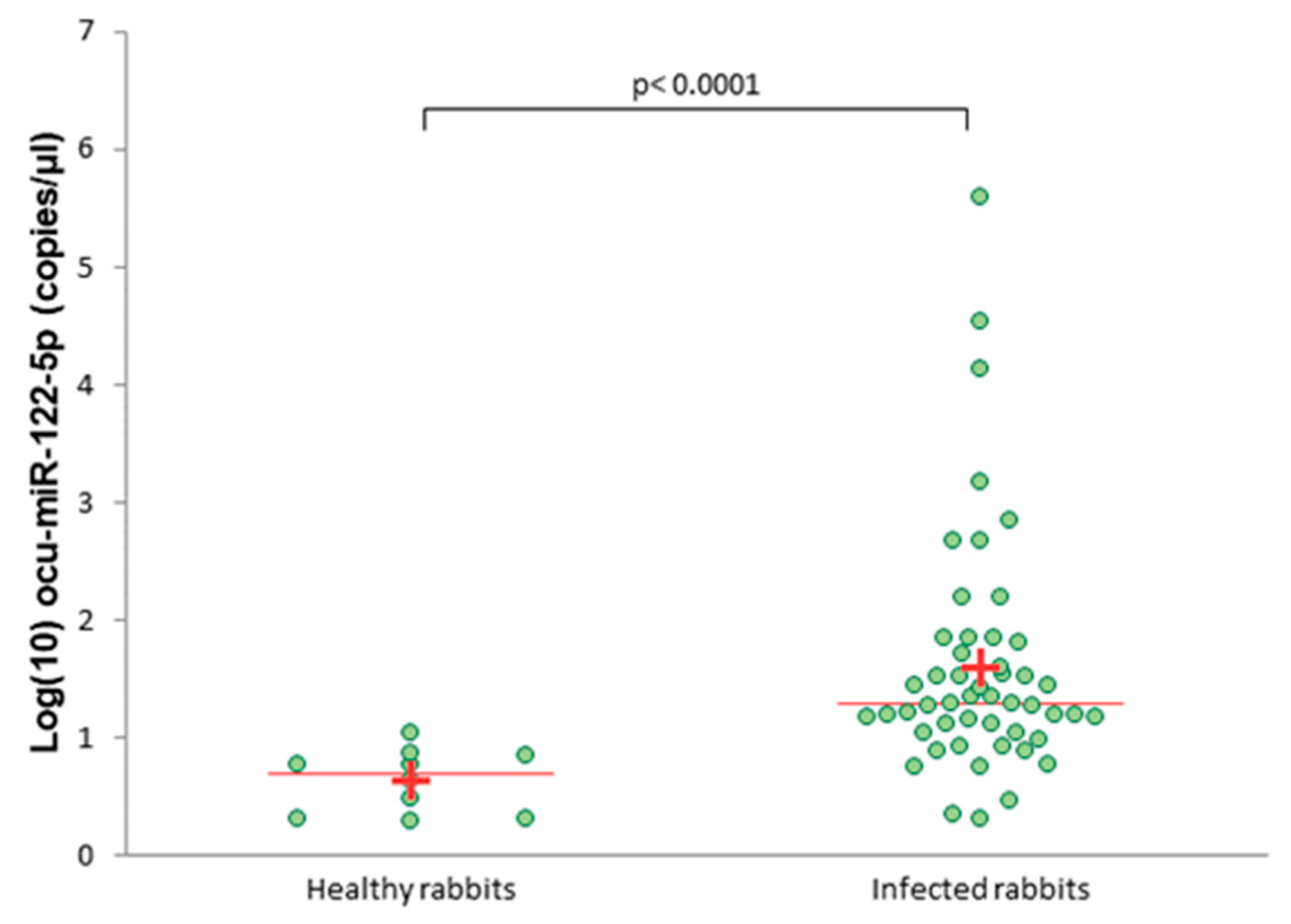
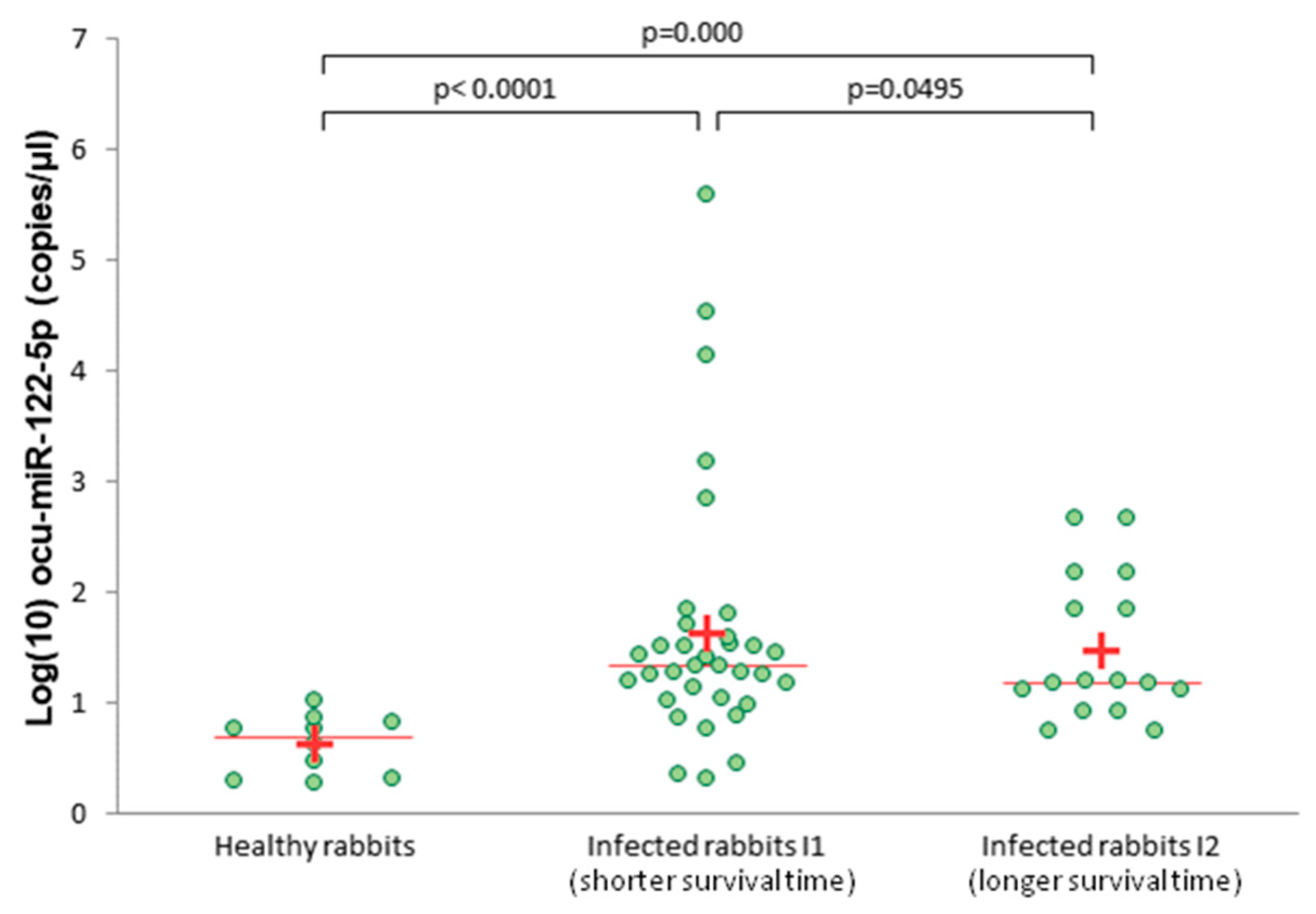
2.8. Quantity of miRNAs in Serum Samples Measured by ddPCR and Data Analysis
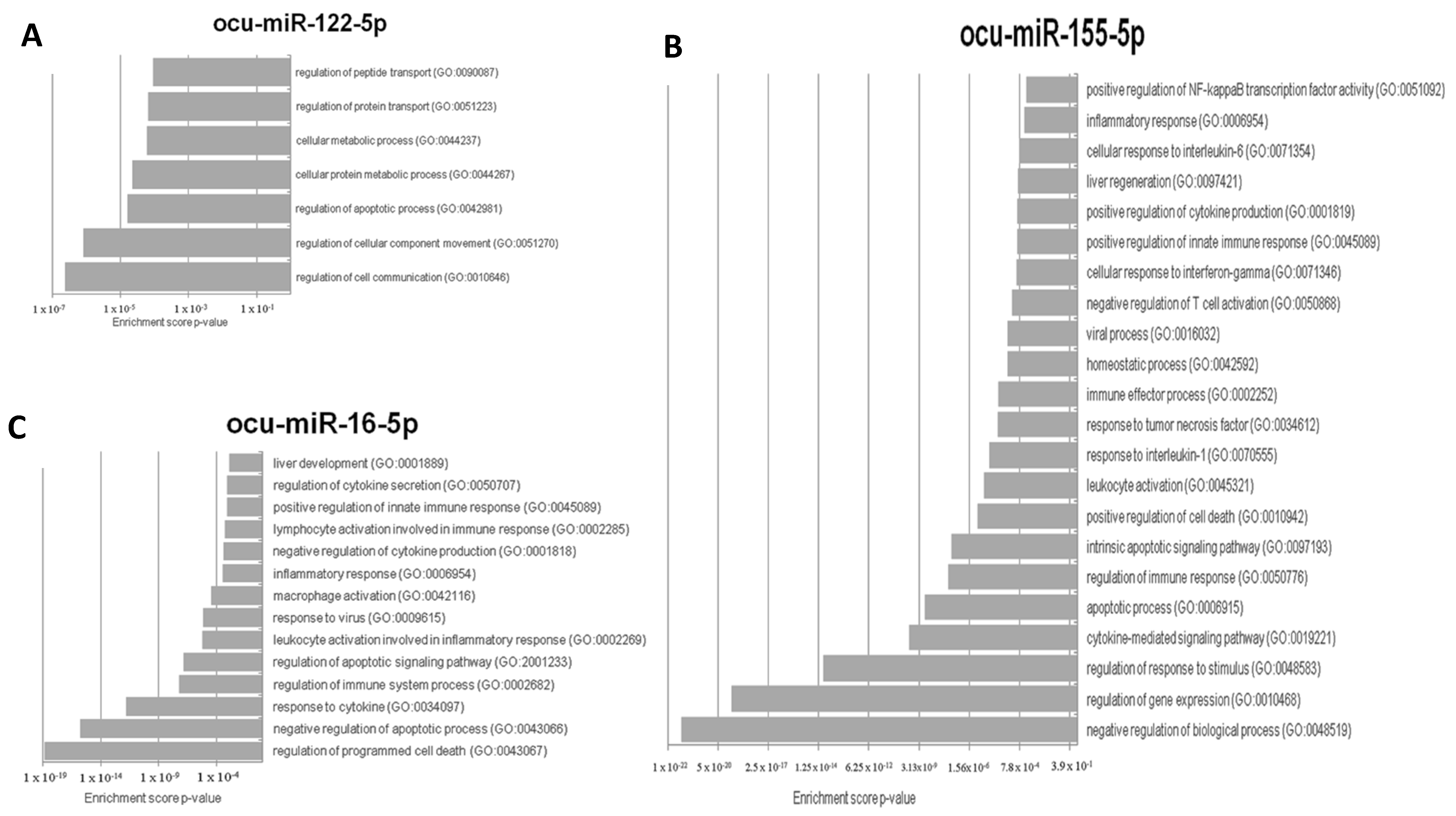
2.9. In Silico Prediction of miR-122-5p, miR-155-5p, and miR-16b-5p Target Genes in Oryctolagus cuniculus
2.10. Statistical Analysis
3. Results
3.1. Clinical Signs of RHD
3.2. Tissue Level of ocu-miR-122-5p, ocu-miR-155-5p, and ocu-miR-16b-5p during RHDV Infection
3.3. Serum Level of ocu-miR-122-5p in Infected Rabbits Versus Healthy Rabbits
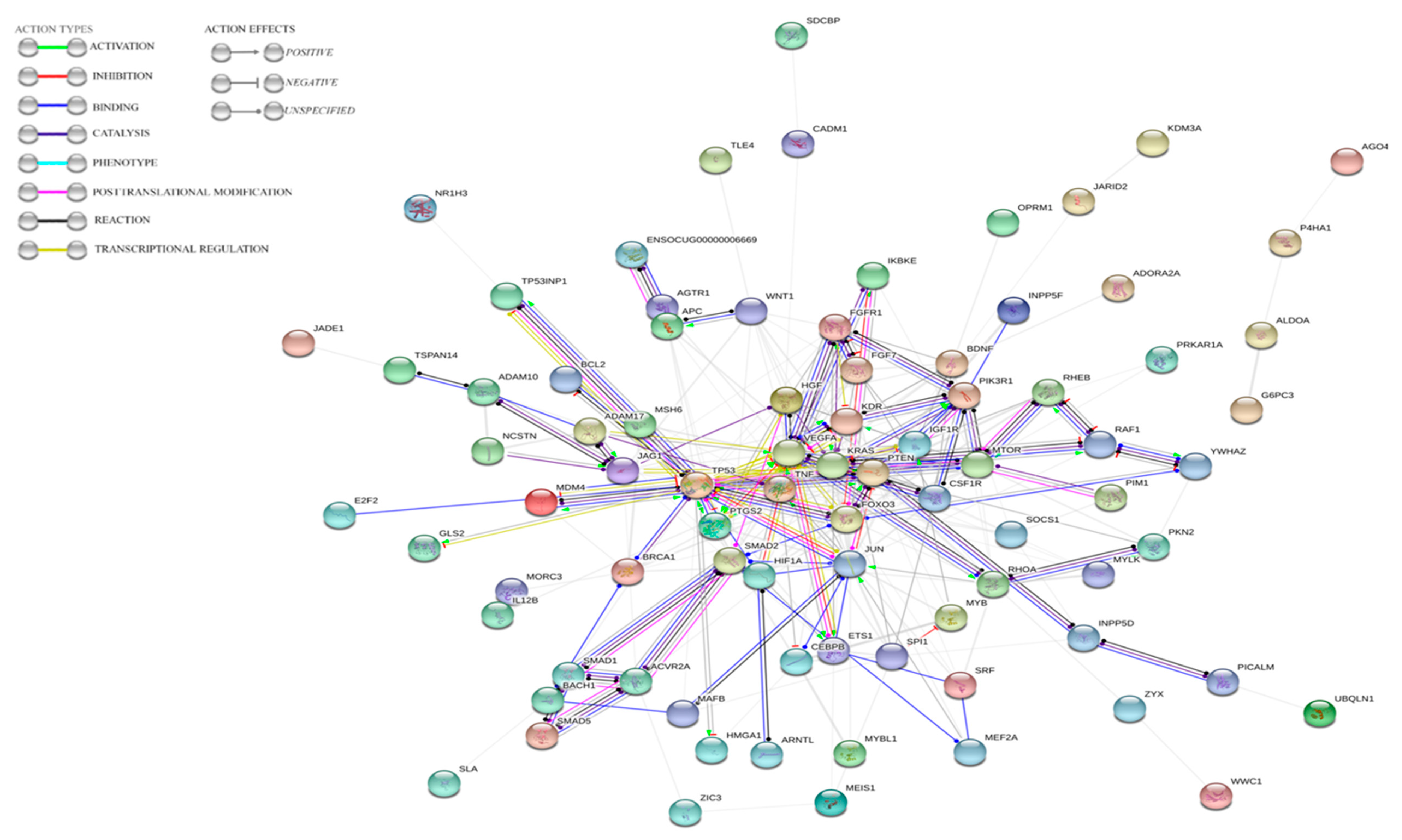
3.4. Prediction of Target Genes for the Three Selected MiRNAs in Oryctolagus cuniculus
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; Le Gall-Recule, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Celio, A.P.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Xue, H.P.; Pu, B.Q.; Qian, N.H. A new viral disease in rabbits. Anim. Husb. Vet. Med. 1984, 16, 253–255. [Google Scholar]
- Hukowska-Szematowicz, B. Genetic variability and phylogenetic analysis of Lagovirus europaeus strains GI.1 (RHDV) and GI.2 (RHDV2) based on the RNA-dependent RNA polymerase (RdRp) coding gene. Acta Biochim. Pol. 2020, 67, 111–122. [Google Scholar] [CrossRef]
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, B.J.; Tai, J.H.; Park, J.H.; Lee, Y.S. Apoptosis in rabbit haemorrhagic disease. J. Comp. Pathol. 2000, 123, 135–140. [Google Scholar] [CrossRef]
- Prieto, J.M.; Fernandez, F.; Alvarez, V.; Espi, A.; Garcia Marin, J.F.; Alvarez, M.; Martin, J.M.; Parra, F. Immunohistochemical localisation of rabbit haemorrhagic disease virus VP-60 antigen in early infection of young and adult rabbits. Res. Vet. Sci. 2000, 68, 181–187. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, Y.S.; Itakura, C. Pathogenesis of acute necrotic hepatitis in rabbit hemorrhagic disease. Lab. Anim. Sci. 1995, 45, 445–449. [Google Scholar]
- Alonso, C.; Oviedo, J.M.; Martin-Alonso, J.M.; Diaz, E.; Boga, J.A.; Parra, F. Programmed cell death in the pathogenesis of rabbit hemorrhagic disease. Arch. Virol. 1998, 143, 321–332. [Google Scholar] [CrossRef]
- San-Miguel, B.; Alvarez, M.; Culebras, J.M.; Gonzalez-Gallego, J.; Tunon, M.J. N-acetyl-cysteine protects liver from apoptotic death in an animal model of fulminant hepatic failure. Apoptosis 2006, 11, 1945–1957. [Google Scholar] [CrossRef]
- Niedzwiedzka-Rystwej, P.; Deptula, W. Lymphocyte subpopulations and apoptosis of immune cells in rabbits experimentally infected with a strain of the RHD virus having a variable haemagglutination capacity. Pol. J. Vet. Sci. 2012, 15, 43–49. [Google Scholar] [CrossRef]
- Marques, R.M.; Costa, E.S.A.; Aguas, A.P.; Teixeira, L.; Ferreira, P.G. Early acute depletion of lymphocytes in calicivirus-infected adult rabbits. Vet. Res. Commun. 2010, 34, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiedzka-Rystwej, P.; Deptula, W. Apoptosis of peripheral blood leukocytes from rabbits infected with non-haemagglutinating strains of rabbit haemorrhagic disease virus (RHDV). Vet. Immunol. Immunopathol. 2012, 149, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Hukowska-Szematowicz, B. Genetic and immunological characteristic of European strains of RHD (rabbit haemorrhagic disease) virus. Pol. J. Environ. Stud. 2013, 2, 1–114. [Google Scholar]
- Trzeciak-Ryczek, A.; Tokarz-Deptula, B.; Deptula, W. The importance of liver lesions and changes to biochemical and coagulation factors in the pathogenesis of RHD. Acta Biochim. Pol. 2015, 62, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, X.; Hu, B.; Fan, Z.; Song, Y.; Wei, H.; Qiu, R.; Xu, W.; Zhu, W.; Wang, F. Rabbit Hemorrhagic Disease Virus Non-structural Protein 6 Induces Apoptosis in Rabbit Kidney Cells. Front. Microbiol. 2018, 9, 3308. [Google Scholar] [CrossRef]
- Tunon, M.J.; San Miguel, B.; Crespo, I.; Riezu-Boj, J.I.; Larrea, E.; Alvarez, M.; Gonzalez, I.; Bustos, M.; Gonzalez-Gallego, J.; Prieto, J. Cardiotrophin-1 promotes a high survival rate in rabbits with lethal fulminant hepatitis of viral origin. J. Virol. 2011, 85, 13124–13132. [Google Scholar] [CrossRef]
- Tunon, M.J.; San Miguel, B.; Crespo, I.; Jorquera, F.; Santamaria, E.; Alvarez, M.; Prieto, J.; Gonzalez-Gallego, J. Melatonin attenuates apoptotic liver damage in fulminant hepatic failure induced by the rabbit hemorrhagic disease virus. J. Pineal Res. 2011, 50, 38–45. [Google Scholar] [CrossRef]
- Tokarz-Deptula, B. Immunity phenomena in rabbits infected with the RHD virus (rabbit haemorrhagic disease). Pol. J. Environ. Sci. 2009, 7, 1–81. [Google Scholar]
- Trzeciak-Ryczek, A.; Tokarz-Deptula, B.; Deptula, W. Expression of IL-1beta, IL-2, IL-10, TNF-beta and GM-CSF in peripheral blood leukocytes of rabbits experimentally infected with rabbit haemorrhagic disease virus. Vet. Microbiol. 2016, 186, 71–81. [Google Scholar] [CrossRef]
- Trzeciak-Ryczek, A.; Tokarz-Deptula, B.; Deptula, W. Expression of IL-1Ra, IL-6, IL-8, IL-18, TNF-alpha and IFN-gamma genes in peripheral blood leukocytes of rabbits infected with RHDV (Rabbit Haemorrhagic Disease Virus). Dev. Comp. Immunol. 2017, 76, 310–315. [Google Scholar] [CrossRef]
- Semerjyan, A.B.; Sargsyan, M.A.; Arzumanyan, H.H.; Hakobyan, L.H.; Abroyan, L.O.; Semerjyan, Z.B.; Avetisyan, A.S.; Karalova, E.M.; Manukyan, D.M.; Matevosyan, H.S.; et al. Immune cell pathology in rabbit hemorrhagic disease. Vet. World 2019, 12, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Ardekani, A.M.; Naeini, M.M. The Role of MicroRNAs in Human Diseases. Avicenna J. Med. Biotechnol. 2010, 2, 161–179. [Google Scholar] [PubMed]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, biogenesis, and their evolving role in animal development and disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yan, I.; Wen, H.J.; Patel, T. microRNAs in liver disease: From diagnostics to therapeutics. Clin. Biochem. 2013, 46, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Pandey, R.K.; Prajapati, P.; Prajapati, V.K. Circulating MicroRNAs: Potential and Emerging Biomarkers for Diagnosis of Human Infectious Diseases. Front. Microbiol. 2016, 7, 1274. [Google Scholar] [CrossRef]
- Loosen, S.H.; Schueller, F.; Trautwein, C.; Roy, S.; Roderburg, C. Role of circulating microRNAs in liver diseases. World J. Hepatol. 2017, 9, 586–594. [Google Scholar] [CrossRef]
- Dong, H.; Gao, Q.; Peng, X.; Sun, Y.; Han, T.; Zhao, B.; Liu, Y.; Wang, C.; Song, X.; Wu, J.; et al. Circulating MicroRNAs As Potential Biomarkers for Veterinary Infectious Diseases. Front. Vet. Sci. 2017, 4, 186. [Google Scholar] [CrossRef]
- Starkey Lewis, P.J.; Dear, J.; Platt, V.; Simpson, K.J.; Craig, D.G.; Antoine, D.J.; French, N.S.; Dhaun, N.; Webb, D.J.; Costello, E.M.; et al. Circulating microRNAs as potential markers of human drug-induced liver injury. Hepatology 2011, 54, 1767–1776. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, S.; Marzolf, B.; Troisch, P.; Brightman, A.; Hu, Z.; Hood, L.E.; Galas, D.J. Circulating microRNAs, potential biomarkers for drug-induced liver injury. Proc. Natl. Acad. Sci. USA 2009, 106, 4402–4407. [Google Scholar] [CrossRef]
- Igaz, P. Circulating microRNA in Disease Diagnostics and Their Potential Biological Relevance; Springer International Publishing: Basel, Switzerland, 2015. [Google Scholar]
- Jopling, C. Liver-specific microRNA-122: Biogenesis and function. RNA Biol. 2012, 9, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Bandiera, S.; Pfeffer, S.; Baumert, T.F.; Zeisel, M.B. miR-122—A key factor and therapeutic target in liver disease. J. Hepatol. 2015, 62, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Schueller, F.; Roy, S.; Vucur, M.; Trautwein, C.; Luedde, T.; Roderburg, C. The Role of miRNAs in the Pathophysiology of Liver Diseases and Toxicity. Int. J. Mol. Sci. 2018, 19, 261. [Google Scholar] [CrossRef]
- Mashima, R. Physiological roles of miR-155. Immunology 2015, 145, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, G.; Biswas, R. MicroRNA-155: A Master Regulator of Inflammation. J. Interferon Cytokine Res. 2019, 39, 321–330. [Google Scholar] [CrossRef]
- Zhang, G.; Yan, C.; Chen, D.; Wu, X.; Zhang, Y.; Zhan, Q.; An, F. Up-regulation of miR-155 contributes to TNF-mediated hepatocyte apoptosis in acute liver failure. Turk. J. Gastroenterol. 2019, 30, 475–484. [Google Scholar] [CrossRef] [PubMed]
- An, F.; Gong, B.; Wang, H.; Yu, D.; Zhao, G.; Lin, L.; Tang, W.; Yu, H.; Bao, S.; Xie, Q. miR-15b and miR-16 regulate TNF mediated hepatocyte apoptosis via BCL2 in acute liver failure. Apoptosis 2012, 17, 702–716. [Google Scholar] [CrossRef]
- Zhan, X.H.; Xu, Q.Y.; Tian, R.; Yan, H.; Zhang, M.; Wu, J.; Wang, W.; He, J. MicroRNA16 regulates glioma cell proliferation, apoptosis and invasion by targeting Wip1-ATM-p53 feedback loop. Oncotarget 2017, 8, 54788–54798. [Google Scholar] [CrossRef]
- Niedzwiedzka-Rystwej, P.; Deptula, W. Non-specific immunity in rabbits infected with 10 strains of the rabbit haemorrhagic disease virus with different biological properties. Cent. Eur. J. Biol. 2010, 5, 613–632. [Google Scholar] [CrossRef]
- Lardizabal, M.N.; Nocito, A.L.; Daniele, S.M.; Ornella, L.A.; Palatnik, J.F.; Veggi, L.M. Reference genes for real-time PCR quantification of microRNAs and messenger RNAs in rat models of hepatotoxicity. PLoS ONE 2012, 7, e36323. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Choi, Y.; Dienes, H.P.; Krawczynski, K. Kinetics of miR-122 expression in the liver during acute HCV infection. PLoS ONE 2013, 8, e76501. [Google Scholar] [CrossRef]
- Sharapova, T.; Devanarayan, V.; LeRoy, B.; Liguori, M.J.; Blomme, E.; Buck, W.; Maher, J. Evaluation of miR-122 as a Serum Biomarker for Hepatotoxicity in Investigative Rat Toxicology Studies. Vet. Pathol. 2016, 53, 211–221. [Google Scholar] [CrossRef]
- Starckx, S.; Batheja, A.; Verheyen, G.R.; Jonghe, S.D.; Steemans, K.; Dijck, B.V.; Singer, M.; Bogdan, N.; Snoeys, J.; Vinken, P.; et al. Evaluation of miR-122 and other biomarkers in distinct acute liver injury in rats. Toxicol. Pathol. 2013, 41, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.S.; Hwang, K.S.; Jeong, Y.M.; Ryu, J.I.; Choi, T.Y.; Bae, M.A.; Son, W.C.; You, K.H.; Son, H.Y.; Kim, C.H. Expression of miRNA-122 Induced by Liver Toxicants in Zebrafish. Biomed. Res. Int. 2016, 2016, 1473578. [Google Scholar] [CrossRef] [PubMed]
- Oosthuyzen, W.; Ten Berg, P.W.L.; Francis, B.; Campbell, S.; Macklin, V.; Milne, E.; Gow, A.G.; Fisher, C.; Mellanby, R.J.; Dear, J.W. Sensitivity and specificity of microRNA-122 for liver disease in dogs. J. Vet. Intern. Med. 2018, 32, 1637–1644. [Google Scholar] [CrossRef] [PubMed]
- Dear, J.W.; Antoine, D.J.; Starkey-Lewis, P.; Goldring, C.E.; Park, B.K. Early detection of paracetamol toxicity using circulating liver microRNA and markers of cell necrosis. Br. J. Clin. Pharmacol. 2014, 77, 904–905. [Google Scholar] [CrossRef] [PubMed]
- Bihrer, V.; Friedrich-Rust, M.; Kronenberger, B.; Forestier, N.; Haupenthal, J.; Shi, Y.; Peveling-Oberhag, J.; Radeke, H.H.; Sarrazin, C.; Herrmann, E.; et al. Serum miR-122 as a biomarker of necroinflammation in patients with chronic hepatitis C virus infection. Am. J. Gastroenterol. 2011, 106, 1663–1669. [Google Scholar] [CrossRef] [PubMed]
- McCrae, J.C.; Sharkey, N.; Webb, D.J.; Vliegenthart, A.D.; Dear, J.W. Ethanol consumption produces a small increase in circulating miR-122 in healthy individuals. Clin. Toxicol. 2016, 54, 53–55. [Google Scholar] [CrossRef]
- Ding, X.; Ding, J.; Ning, J.; Yi, F.; Chen, J.; Zhao, D.; Zheng, J.; Liang, Z.; Hu, Z.; Du, Q. Circulating microRNA-122 as a potential biomarker for liver injury. Mol. Med. Rep. 2012, 5, 1428–1432. [Google Scholar] [CrossRef]
- Robinson, S.; Follo, M.; Haenel, D.; Mauler, M.; Stallmann, D.; Tewari, M.; Duerschmied, D.; Peter, K.; Bode, C.; Ahrens, I.; et al. Droplet digital PCR as a novel detection method for quantifying microRNAs in acute myocardial infarction. Int. J. Cardiol. 2018, 257, 247–254. [Google Scholar] [CrossRef]
- Blaya, D.; Aguilar-Bravo, B.; Hao, F.; Casacuberta-Serra, S.; Coll, M.; Perea, L.; Vallverdu, J.; Graupera, I.; Pose, E.; Llovet, L.; et al. Expression of microRNA-155 in inflammatory cells modulates liver injury. Hepatology 2018, 68, 691–706. [Google Scholar] [CrossRef]
- Bala, S.; Marcos, M.; Kodys, K.; Csak, T.; Catalano, D.; Mandrekar, P.; Szabo, G. Up-regulation of microRNA-155 in macrophages contributes to increased tumor necrosis factor (TNF{alpha}) production via increased mRNA half-life in alcoholic liver disease. J. Biol. Chem. 2011, 286, 1436–1444. [Google Scholar] [CrossRef]
- Sanchez-Campos, S.; Alvarez, M.; Culebras, J.M.; Gonzalez-Gallego, J.; Tunon, M.J. Pathogenic molecular mechanisms in an animal model of fulminant hepatic failure: Rabbit hemorrhagic viral disease. J. Lab. Clin. Med. 2004, 144, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, A.; Chung, R.T. Acute liver failure: Mechanisms of hepatocyte injury and regeneration. Semin. Liver Dis. 2008, 28, 167–174. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNAs | Sequences |
|---|---|
| miRNAs tested | |
| ocu-miR-122-5p | 5′UGGAGUGUGACAAUGGUGUUUG3′ |
| ocu-miR-155-5p | 5′UUAAUGCUAAUCGUGAUAGGGGUU3′ |
| ocu-miR-16b-5p | 5′UAGCAGCACGUAAAUAUUGGCGU3′ |
| reference miRNAs | |
| ocu-let-7a-5p | 5′UGAGGUAGUAGGUUGUAUAGUU3′ |
| ocu-miR-103a-3p | 5′AGCAGCAUUGUACAGGGCUAUGA3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hukowska-Szematowicz, B.; Maciejak-Jastrzębska, A.; Blatkiewicz, M.; Maciak, K.; Góra, M.; Janiszewska, J.; Burzyńska, B. Changes in MicroRNA Expression during Rabbit Hemorrhagic Disease Virus (RHDV) Infection. Viruses 2020, 12, 965. https://doi.org/10.3390/v12090965
Hukowska-Szematowicz B, Maciejak-Jastrzębska A, Blatkiewicz M, Maciak K, Góra M, Janiszewska J, Burzyńska B. Changes in MicroRNA Expression during Rabbit Hemorrhagic Disease Virus (RHDV) Infection. Viruses. 2020; 12(9):965. https://doi.org/10.3390/v12090965
Chicago/Turabian StyleHukowska-Szematowicz, Beata, Agata Maciejak-Jastrzębska, Małgorzata Blatkiewicz, Karolina Maciak, Monika Góra, Joanna Janiszewska, and Beata Burzyńska. 2020. "Changes in MicroRNA Expression during Rabbit Hemorrhagic Disease Virus (RHDV) Infection" Viruses 12, no. 9: 965. https://doi.org/10.3390/v12090965
APA StyleHukowska-Szematowicz, B., Maciejak-Jastrzębska, A., Blatkiewicz, M., Maciak, K., Góra, M., Janiszewska, J., & Burzyńska, B. (2020). Changes in MicroRNA Expression during Rabbit Hemorrhagic Disease Virus (RHDV) Infection. Viruses, 12(9), 965. https://doi.org/10.3390/v12090965