Chronic Hepatitis B Infection Is Associated with Increased Molecular Degree of Inflammatory Perturbation in Peripheral Blood

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Study Design
2.3. Laboratory Measurements
2.4. Adaptation of Molecular Degree of Perturbation to Examine Plasma Concentrations of Biomarkers
- n = Number of data points
- xi = Each of the value of data
- = Mean of the data points
- σ = Standard deviation
2.5. Statistical Analysis
3. Results
3.1. Characteristics of Participants
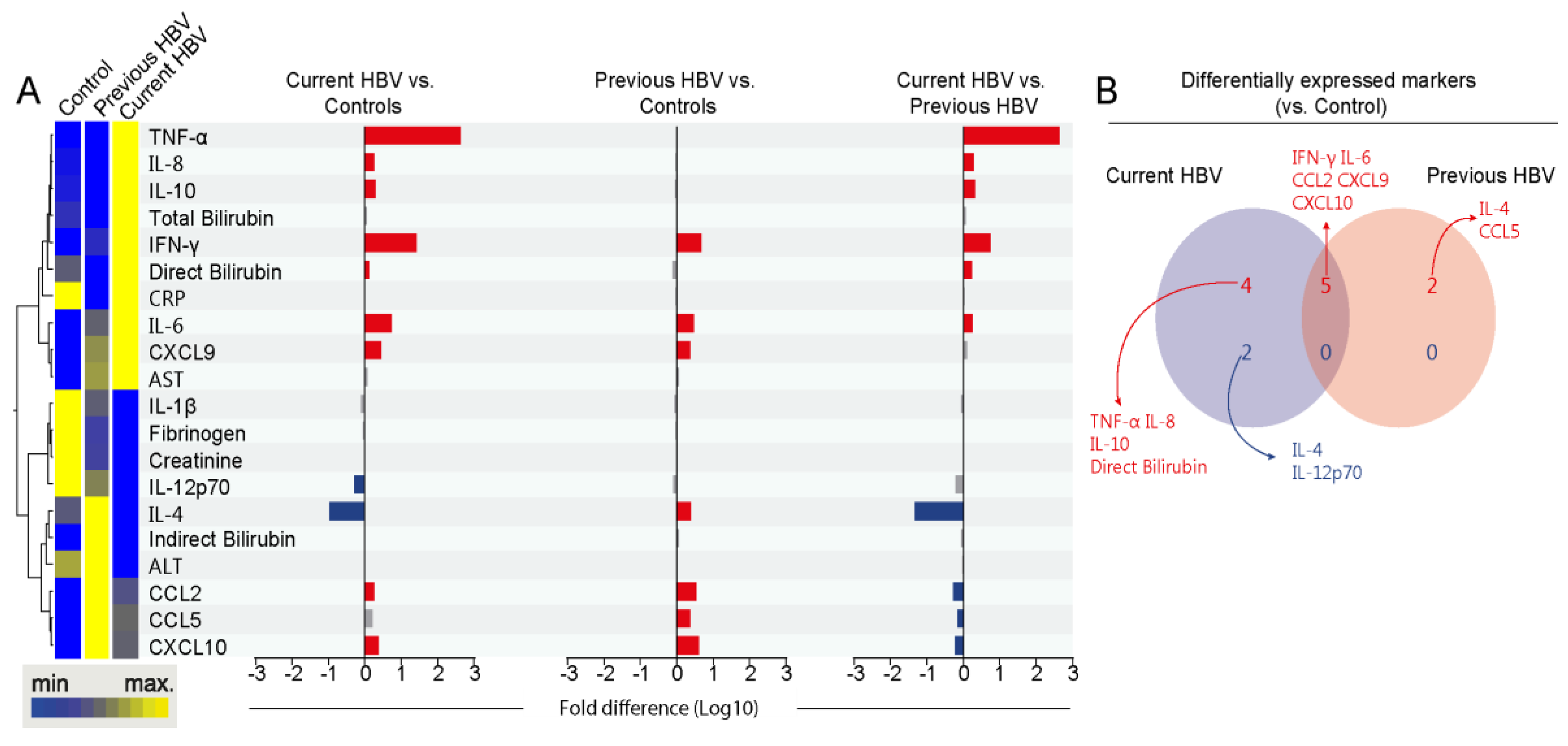
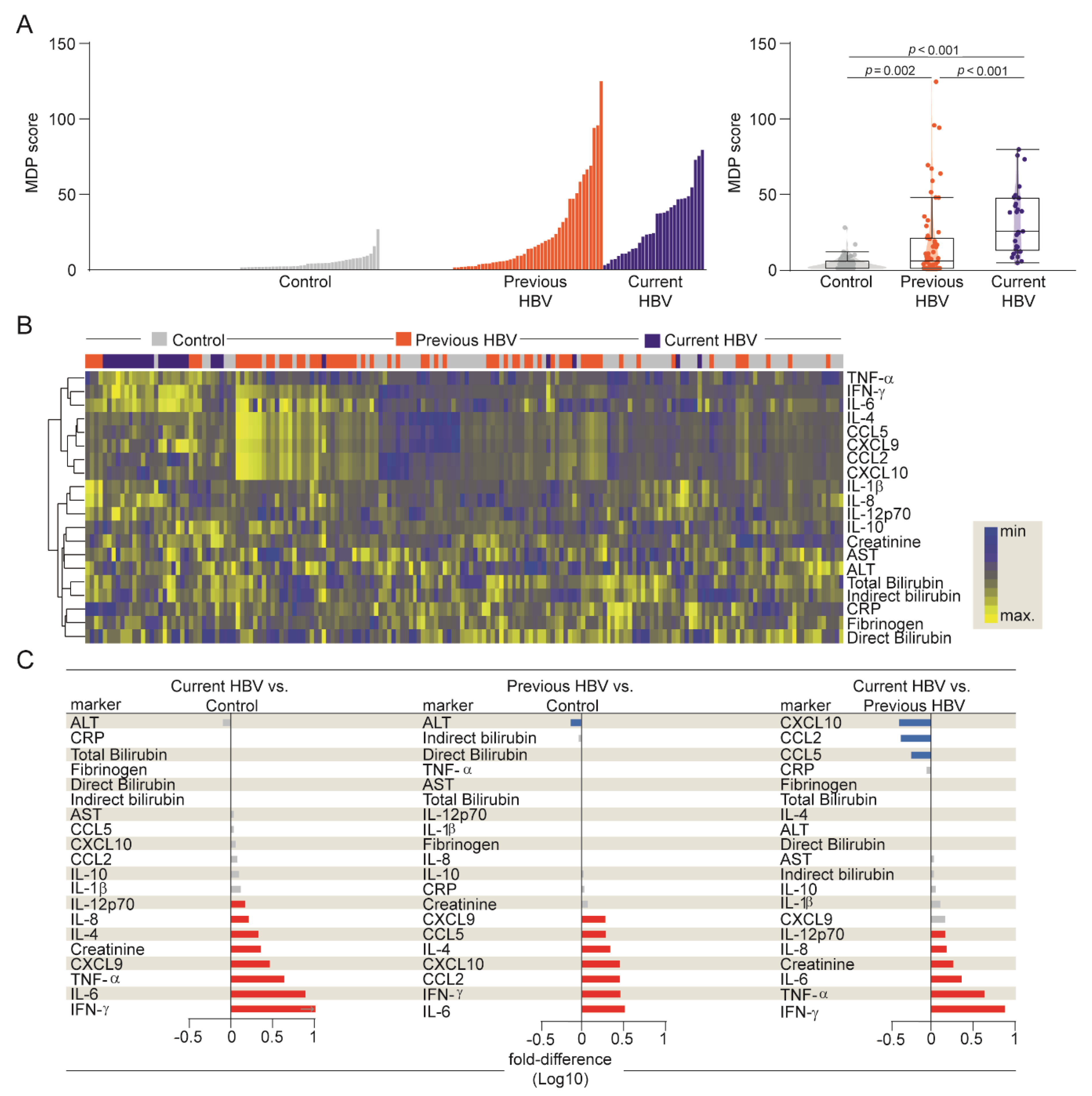
3.2. Differences in Systemic Inflammation between the Study Subgroups
3.3. HBV Infection is Associated with an Increased Degree of Inflammatory Perturbation
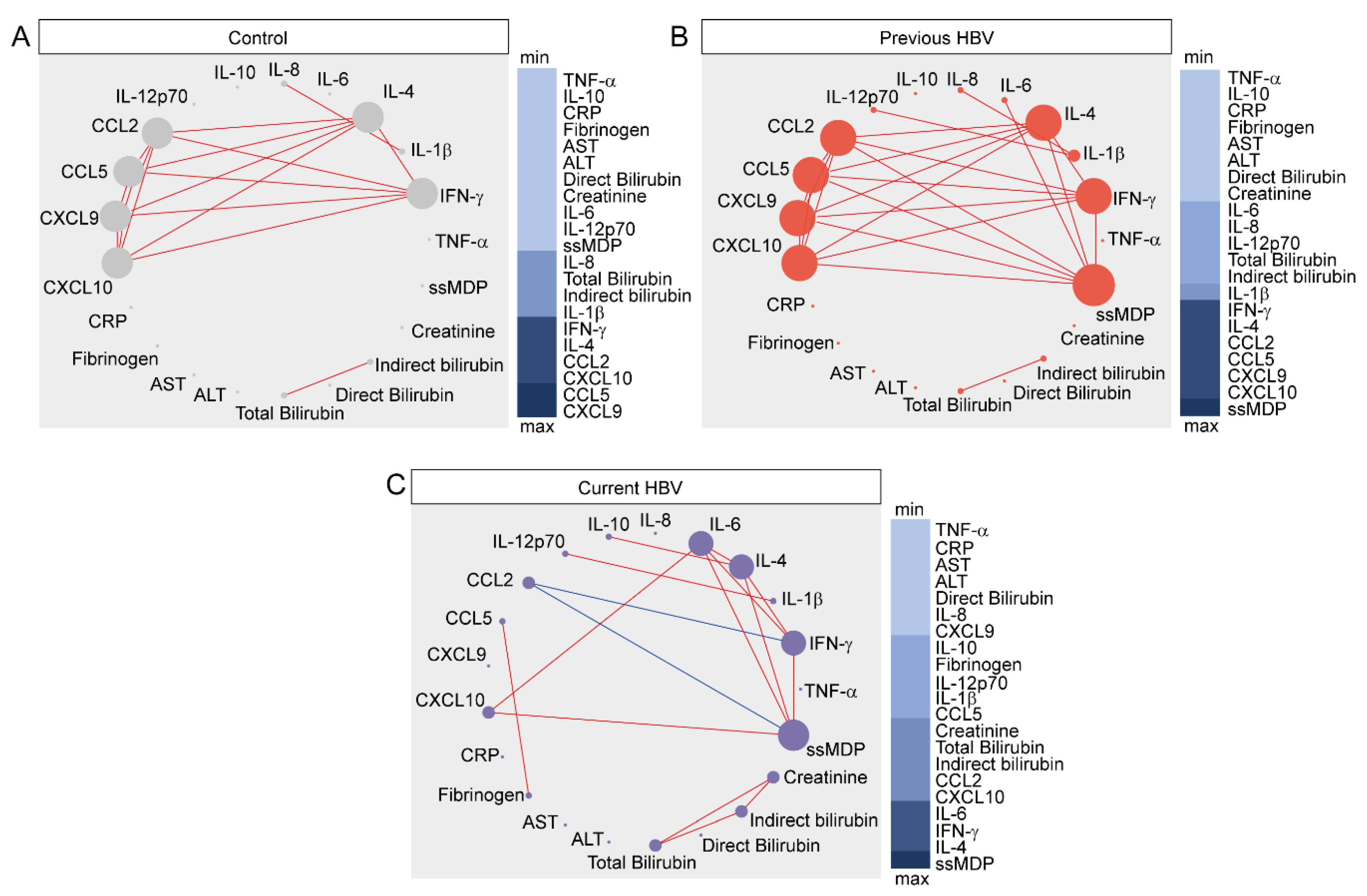
3.4. Network Analysis of Inflammatory Imbalance in Chronic HBV Infection
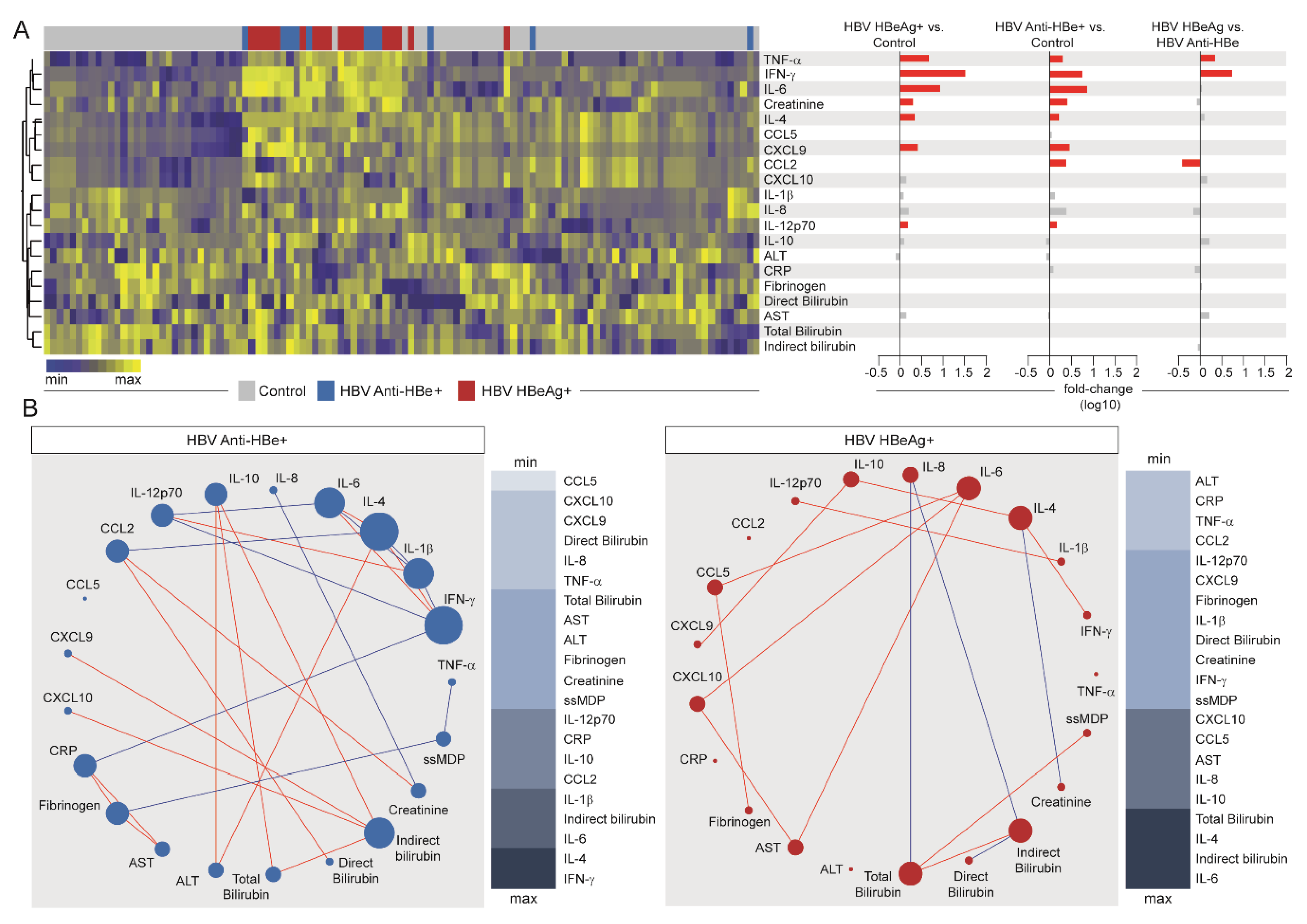
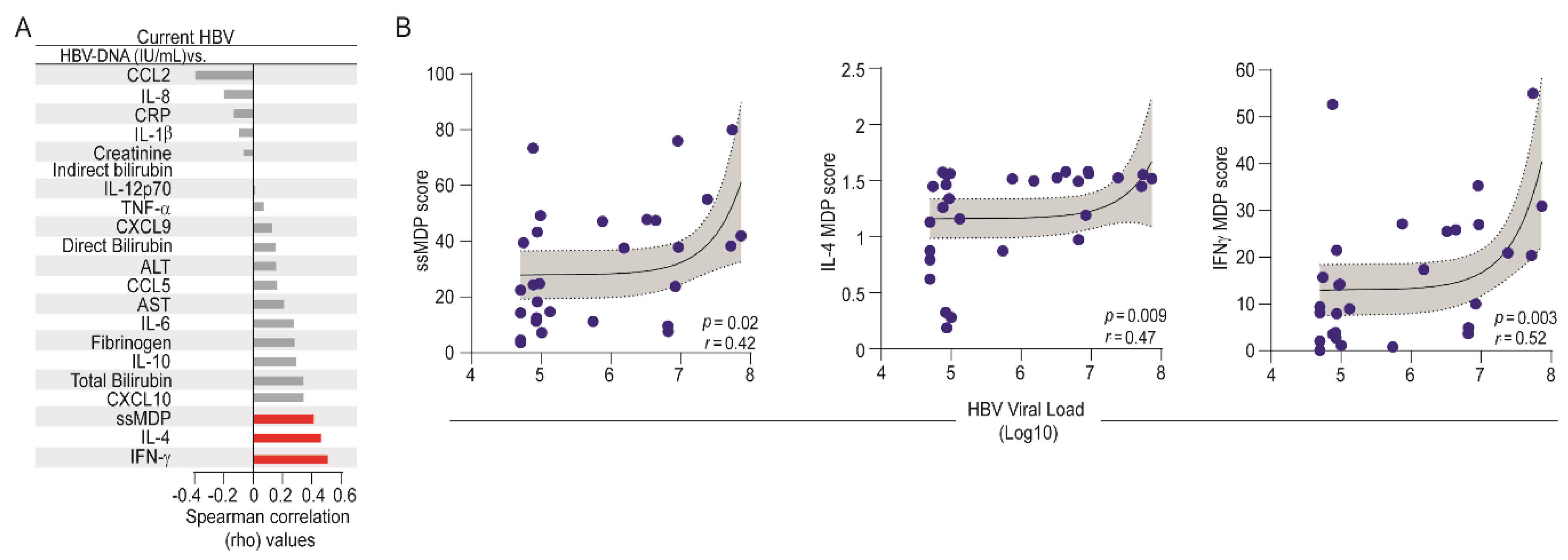
3.5. Influence of HBV Viral Replication on the Systemic Inflammatory Imbalance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tarocchi, M.; Polvani, S.; Marroncini, G.; Galli, A. Molecular mechanism of hepatitis B virus-induced hepatocarcinogenesis. World J. Gastroenterol. 2014, 20, 11630–11640. [Google Scholar] [CrossRef]
- WHO. Global Hepatitis Report 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Naghavi, M.; Abajobir, A.A.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abera, S.F.; Aboyans, V.; Adetokunboh, O.O.; Afshin, A.; Agrawal, A.; et al. Global, regional, and national age-sex specific mortality for 264 causes of death, 1980–2016: A systematic analysis for the global burden of disease study 2016. Lancet 2017, 390, 1151–1210. [Google Scholar] [CrossRef] [Green Version]
- Ganem, D.; Prince, A.M. Hepatitis B virus infection—Natural history and clinical consequences. N. Engl. J. Med. 2004, 350, 1118–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuen, M.F.; Chen, D.S.; Dusheiko, G.M.; Janssen, H.L.A.; Lau, D.T.Y.; Locarnini, S.A.; Peters, M.G.; Lai, C.L. Hepatitis B virus infection. Nat. Rev. Dis. Prim. 2018, 4, 18035. [Google Scholar] [CrossRef] [PubMed]
- Trépo, C.; Chan, H.L.Y.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- Mason, W.S.; Gill, U.S.; Litwin, S.; Zhou, Y.; Peri, S.; Pop, O.; Hong, M.; Naik, S.; Quaglia, A.; Bertoletti, A.; et al. HBV DNA integration and clonal hepatocyte expansion in chronic hepatitis B patients considered immune tolerant. Gastroenterology 2016, 151, 986–998.e4. [Google Scholar] [CrossRef] [Green Version]
- Bertoletti, A.; Ferrari, C. Innate and adaptive immune responses in chronic hepatitis B virus infections: Towards restoration of immune control of viral infection. Gut 2012, 61, 1754. [Google Scholar] [CrossRef]
- Boni, C.; Fisicaro, P.; Valdatta, C.; Amadei, B.; Di Vincenzo, P.; Giuberti, T.; Laccabue, D.; Zerbini, A.; Cavalli, A.; Missale, G.; et al. Characterization of hepatitis B virus (HBV)-specific T-cell dysfunction in chronic HBV infection. J. Virol. 2007, 81, 4215–4225. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J. T cell exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Wherry, E.J.; Blattman, J.N.; Murali-Krishna, K.; Van Der Most, R.; Ahmed, R. Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment. J. Virol. 2003, 77, 4911–4927. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Raziorrouh, B.; Schraut, W.; Gerlach, T.; Nowack, D.; Grüner, N.H.; Ulsenheimer, A.; Zachoval, R.; Wächtler, M.; Spannagl, M.; Haas, J.; et al. The immunoregulatory role of CD244 in chronic hepatitis B infection and its inhibitory potential on virus-specific CD8+ T-cell function. Hepatology 2010, 52, 1934–1947. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.G.; Trifilo, M.J.; Edelmann, K.H.; Teyton, L.; McGAVERN, D.B.; Oldstone, M.B. Interleukin-10 determines viral clearance or persistence in vivo. Nat. Med. 2006, 12, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Ellis, G.; Pallant, C.; Lopes, A.R.; Khanna, P.; Peppa, D.; Chen, A.; Blair, P.; Dusheiko, G.; Gill, U.; et al. IL-10-producing regulatory B cells in the pathogenesis of chronic hepatitis B virus infection. J. Immunol. 2012, 189, 3925–3935. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.L.; Yang, J.-R.; Lin, S.Z.; Ma, H.; Guo, F.; Yang, R.F.; Zhang, H.H.; Han, J.C.; Wei, L.; Pan, X.B. Serum viral duplex-linear DNA proportion increases with the progression of liver disease in patients infected with HBV. Gut 2015, 65, 502–511. [Google Scholar] [CrossRef]
- Vinhaes, C.L.; Arriaga, M.B.; De Almeida, B.L.; Oliveira, J.V.; Santos, C.S.I.; Calcagno, J.; Carvalho, T.X.; Giovanetti, M.; Alcantara, L.C.J.; De Siqueira, I.C.; et al. Newborns with zika virus-associated microcephaly exhibit marked systemic inflammatory imbalance. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Oliveira-De-Souza, D.; Vinhaes, C.L.; Arriaga, M.B.; Kumar, N.P.; Angulo, J.M.C.; Shi, R.; Wei, W.; Yuan, X.; Zhang, G.; Cai, Y.; et al. Molecular degree of perturbation of plasma inflammatory markers associated with tuberculosis reveals distinct disease profiles between Indian and Chinese populations. Sci. Rep. 2019, 9, 8002. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-De-Souza, D.; Vinhaes, C.L.; Arriaga, M.B.; Kumar, N.P.; Queiroz, A.T.L.; Fukutani, K.F.; Babu, S.; Andrade, B.B. Aging increases the systemic molecular degree of inflammatory perturbation in patients with tuberculosis. Sci. Rep. 2020, 10, 11358. [Google Scholar] [CrossRef]
- Vinhaes, C.L.; Oliveira-De-Souza, D.; Silveira-Mattos, P.S.; Nogueira, B.; Shi, R.; Wei, W.; Yuan, X.; Zhang, G.; Cai, Y.; Barry, C.E.; et al. Changes in inflammatory protein and lipid mediator profiles persist after antitubercular treatment of pulmonary and extrapulmonary tuberculosis: A prospective cohort study. Cytokine 2019, 123, 154759. [Google Scholar] [CrossRef]
- Prada-Medina, C.A.; Fukutani, K.F.; Kumar, N.P.; Gil-Santana, L.; Babu, S.; Lichtenstein, F.; West, K.; Sivakumar, S.; Menon, P.A.; Viswanathan, V.; et al. Systems immunology of diabetes-tuberculosis comorbidity reveals signatures of disease complications. Sci. Rep. 2017, 7, 1999. [Google Scholar] [CrossRef]
- Barretto, J.R.; Boa-Sorte, N.; Vinhaes, C.L.; Malta-Santos, H.; Rebouças-Silva, J.; Ramos, C.F.; Torres-Nascimento, M.A.S.; Borges, V.M.; Andrade, B.B. Heightened plasma levels of transforming growth factor beta (TGF-β) and increased degree of systemic biochemical perturbation characterizes hepatic steatosis in overweight pediatric patients: A cross-sectional study. Nutrients 2020, 12, 1650. [Google Scholar] [CrossRef] [PubMed]
- Andrade, B.B.; Santos, C.J.N.; Camargo, L.M.; Souza-Neto, S.M.; Reis-Filho, A.; Clarêncio, J.; Mendonça, V.R.R.; Luz, N.F.; Camargo, E.P.; Barral, A.; et al. Hepatitis B infection is associated with asymptomatic malaria in the brazilian amazon. PLoS ONE 2011, 6, e19841. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.A.B.; Netto, M.B.; Andrade, B.B. Distinct inflammatory profile underlies pathological increases in creatinine levels associated with Plasmodium vivax malaria clinical severity. PLoS Neglected Trop. Dis. 2018, 12, e0006306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, L.A.B.; Moraes, M.O.A.; Queiroga-Barros, M.R.; Fukutani, K.F.; Netto, M.B.; Andrade, B.B. Chronic hepatitis B virus infection drives changes in systemic immune activation profile in patients coinfected with Plasmodium vivax malaria. PLoS Neglected Trop. Dis. 2019, 13, e0007535. [Google Scholar] [CrossRef] [PubMed]
- Andrade, B.B.; Reis-Filho, A.; Souza-Neto, S.M.; Clarêncio, J.; Camargo, L.M.A.; Barral, A.; Netto, M.B. Severe Plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar. J. 2010, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Mendonça, V.R.; Queiroz, A.T.L.; Lopes, F.M.; Andrade, B.B.; Netto, M.B. Networking the host immune response in Plasmodium vivax malaria. Malar. J. 2013, 12, 69. [Google Scholar] [CrossRef] [Green Version]
- Gozzelino, R.; Andrade, B.B.; Larsen, R.; Luz, N.F.; Vanoaica, L.; Seixas, E.; Coutinho, A.; Cardoso, S.; Rebelo, S.; Poli, M.; et al. Metabolic adaptation to tissue iron overload confers tolerance to malaria. Cell Host Microbe 2012, 12, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; Reis-Filho, A.; Barros, A.M.; Souza-Neto, S.M.; Nogueira, L.D.L.; Fukutani, K.F.; Camargo, E.P.; Camargo, L.M.A.; Barral, A.; Duarte, A.; et al. Towards a precise test for malaria diagnosis in the Brazilian Amazon: Comparison among field microscopy, a rapid diagnostic test, nested PCR, and a computational expert system based on artificial neural networks. Malar. J. 2010, 9, 117. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; Rocha, B.C.; Filho, A.R.; Camargo, L.M.A.; Tadei, W.P.; Moreira, L.A.; Barral, A.; Netto, M.B. Anti-Anopheles darlingi saliva antibodies as marker of Plasmodium vivax infection and clinical immunity in the Brazilian Amazon. Malar. J. 2009, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; Araújo-Santos, T.; Luz, N.F.; Khouri, R.; Bozza, M.T.; Camargo, L.M.A.; Barral, A.; Borges, V.M.; Netto, M.B. Heme impairs prostaglandin E2 and TGF-β production by human mononuclear cells via cu/zn superoxide dismutase: Insight into the pathogenesis of severe malaria. J. Immunol. 2010, 185, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; Reis-Filho, A.; Souza-Neto, S.M.; Raffaele-Netto, I.; Camargo, L.M.A.; Barral, A.; Netto, M.B. Plasma superoxide dismutase-1 as a surrogate marker of vivax malaria severity. PLoS Neglected Trop. Dis. 2010, 4, e650. [Google Scholar] [CrossRef] [PubMed]
- Jamil, B.; Shahid, F.; Hasan, Z.; Nasir, N.; Razzaki, T.; Dawood, G.; Hussain, R. Interferonγ/IL10 ratio defines the disease severity in pulmonary and extra pulmonary tuberculosis. Tuberculosis 2007, 87, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Protzer, U. Control of hepatitis B virus by cytokines. Viruses 2017, 9, 18. [Google Scholar] [CrossRef]
- Raman, D.; Sobolik-Delmaire, T.; Richmond, A. Chemokines in health and disease. Exp. Cell Res. 2011, 317, 575–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Guo, S.; Hibbert, J.M.; Jain, V.; Singh, N.; Wilson, N.O.; Stiles, J.K. CXCL10/IP-10 in infectious diseases pathogenesis and potential therapeutic implications. Cytokine Growth Factor Rev. 2011, 22, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Mohs, A.; Kuttkat, N.; Reißing, J.; Zimmermann, H.W.; Sonntag, R.; Proudfoot, A.; Youssef, S.; De Bruin, A.; Cubero, F.J.; Trautwein, C. Functional role of CCL5/RANTES for HCC progression during chronic liver disease. J. Hepatol. 2017, 66, 743–753. [Google Scholar] [CrossRef]
- Hokeness, K.L.; Kuziel, W.A.; Biron, C.A.; Salazar-Mather, T.P. Monocyte chemoattractant protein-1 and ccr2 interactions are required for ifn-α/β-induced inflammatory responses and antiviral defense in liver. J. Immunol. 2005, 174, 1549–1556. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 2011, 11, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Hösel, M.; Quasdorff, M.; Ringelhan, M.; Kashkar, H.; Debey-Pascher, S.; Sprinzl, M.F.; Bockmann, J.-H.; Arzberger, S.; Webb, D.; Von Olshausen, G.; et al. Hepatitis B virus activates signal transducer and activator of transcription 3 supporting hepatocyte survival and virus replication. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 339–363. [Google Scholar] [CrossRef]
- Karpus, W.J.; Lukacs, N.W.; Kennedy, K.J.; Smith, W.S.; Hurst, S.D.; Barrett, T. Differential CC chemokine-induced enhancement of T helper cell cytokine production. J. Immunol. 1997, 158, 4129. [Google Scholar]
- Estevez, J.; Chen, V.L.; Podlaha, O.; Li, B.; Le, A.; Vutien, P.; Chang, E.T.; Rosenberg-Hasson, Y.; Jiang, Z.; Pflanz, S.; et al. Differential serum cytokine profiles in patients with chronic hepatitis b, c, and hepatocellular carcinoma. Sci. Rep. 2017, 7, 11867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, T.W.; Yevsa, T.; Woller, N.; Hoenicke, L.; Wuestefeld, T.; Dauch, D.; Hohmeyer, A.; Gereke, M.; Rudalska, R.; Potapova, A.; et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature 2011, 479, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Poon, R.T.; Hughes, J.; Li, Q.Y.; Yu, W.C.; Fan, S.T. Desensitization of T lymphocyte function by CXCR3 ligands in human hepatocellular carcinoma. World J. Gastroenterol. 2005, 11, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Zacharakis, G.; Koskinas, J.; Kotsiou, S.; Tzara, F.; Vafeiadis, N.; Papoutselis, M.; Maltezos, E.; Sivridis, E.; Papoutselis, K. The role of serial measurement of serum HBV DNA levels in patients with chronic HBeAg(−) hepatitis B infection: Association with liver disease progression. A prospective cohort study. J. Hepatol. 2008, 49, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Manion, M.; Andrade, B.B.; DerSimonian, R.; Gu, W.; Rupert, A.; Musselwhite, L.W.; Sierra-Madero, J.G.; Belaunzaran-Zamudio, P.F.; Sanne, I.; Lederman, M.M.; et al. Country of residence is associated with distinct inflammatory biomarker signatures in HIV-infected patients. J. Virus Erad. 2017, 3, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Lian, Y.; Gu, L.; Chen, L.; Li, X.; Zhou, L.; Huang, Y.; Wang, J.; Huang, Y. Correlations between cytokines produced by T cells and clinical-virological characteristics in untreated chronic hepatitis B patients. BMC Infect. Dis. 2019, 19, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.J.; Yang, H.I.; Iloeje, U.H. The reveal-HBV study group hepatitis B virus DNA levels and outcomes in chronic hepatitis B. Hepatology 2009, 49, S72–S84. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinhaes, C.L.; Cruz, L.A.B.; Menezes, R.C.; Carmo, T.A.; Arriaga, M.B.; Queiroz, A.T.L.; Barral-Netto, M.; Andrade, B.B. Chronic Hepatitis B Infection Is Associated with Increased Molecular Degree of Inflammatory Perturbation in Peripheral Blood. Viruses 2020, 12, 864. https://doi.org/10.3390/v12080864
Vinhaes CL, Cruz LAB, Menezes RC, Carmo TA, Arriaga MB, Queiroz ATL, Barral-Netto M, Andrade BB. Chronic Hepatitis B Infection Is Associated with Increased Molecular Degree of Inflammatory Perturbation in Peripheral Blood. Viruses. 2020; 12(8):864. https://doi.org/10.3390/v12080864
Chicago/Turabian StyleVinhaes, Caian L., Luís A. B. Cruz, Rodrigo C. Menezes, Thomas A. Carmo, María B. Arriaga, Artur T. L. Queiroz, Manoel Barral-Netto, and Bruno B. Andrade. 2020. "Chronic Hepatitis B Infection Is Associated with Increased Molecular Degree of Inflammatory Perturbation in Peripheral Blood" Viruses 12, no. 8: 864. https://doi.org/10.3390/v12080864
APA StyleVinhaes, C. L., Cruz, L. A. B., Menezes, R. C., Carmo, T. A., Arriaga, M. B., Queiroz, A. T. L., Barral-Netto, M., & Andrade, B. B. (2020). Chronic Hepatitis B Infection Is Associated with Increased Molecular Degree of Inflammatory Perturbation in Peripheral Blood. Viruses, 12(8), 864. https://doi.org/10.3390/v12080864