HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Patients and Methods
Study Design and Participants
3. Results
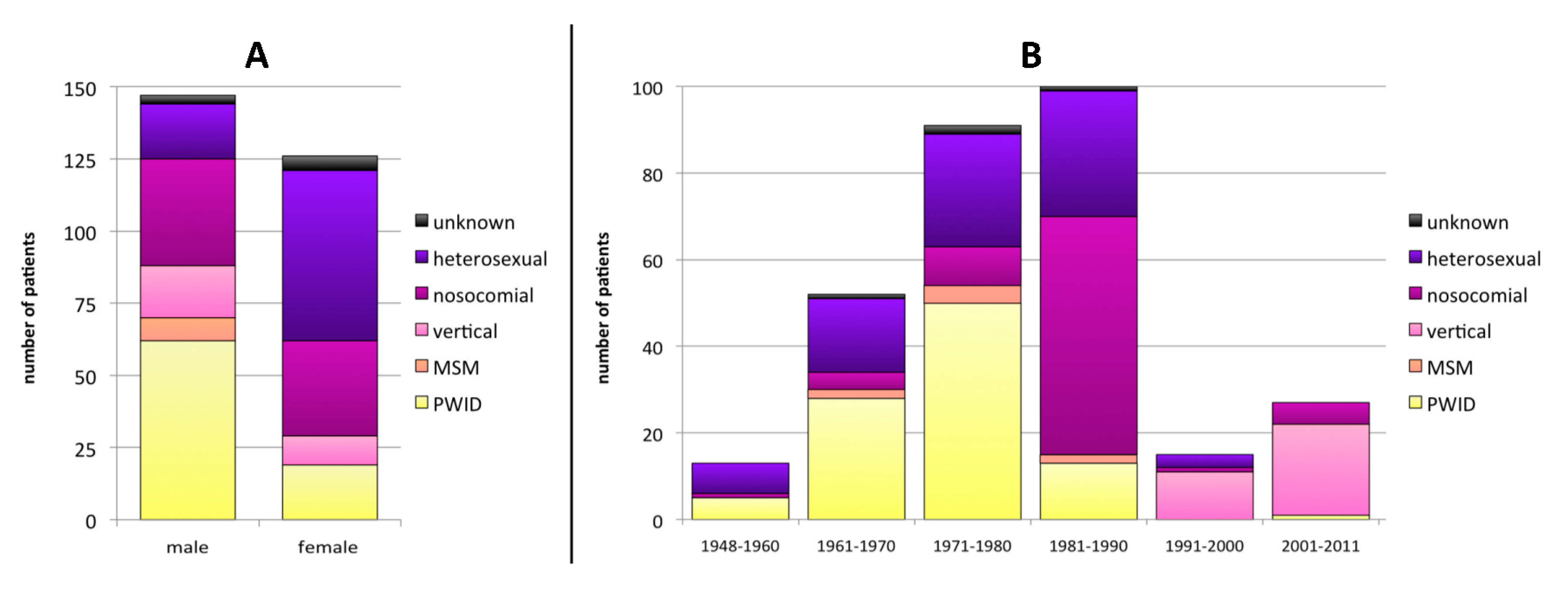
3.1. Baseline Characteristics
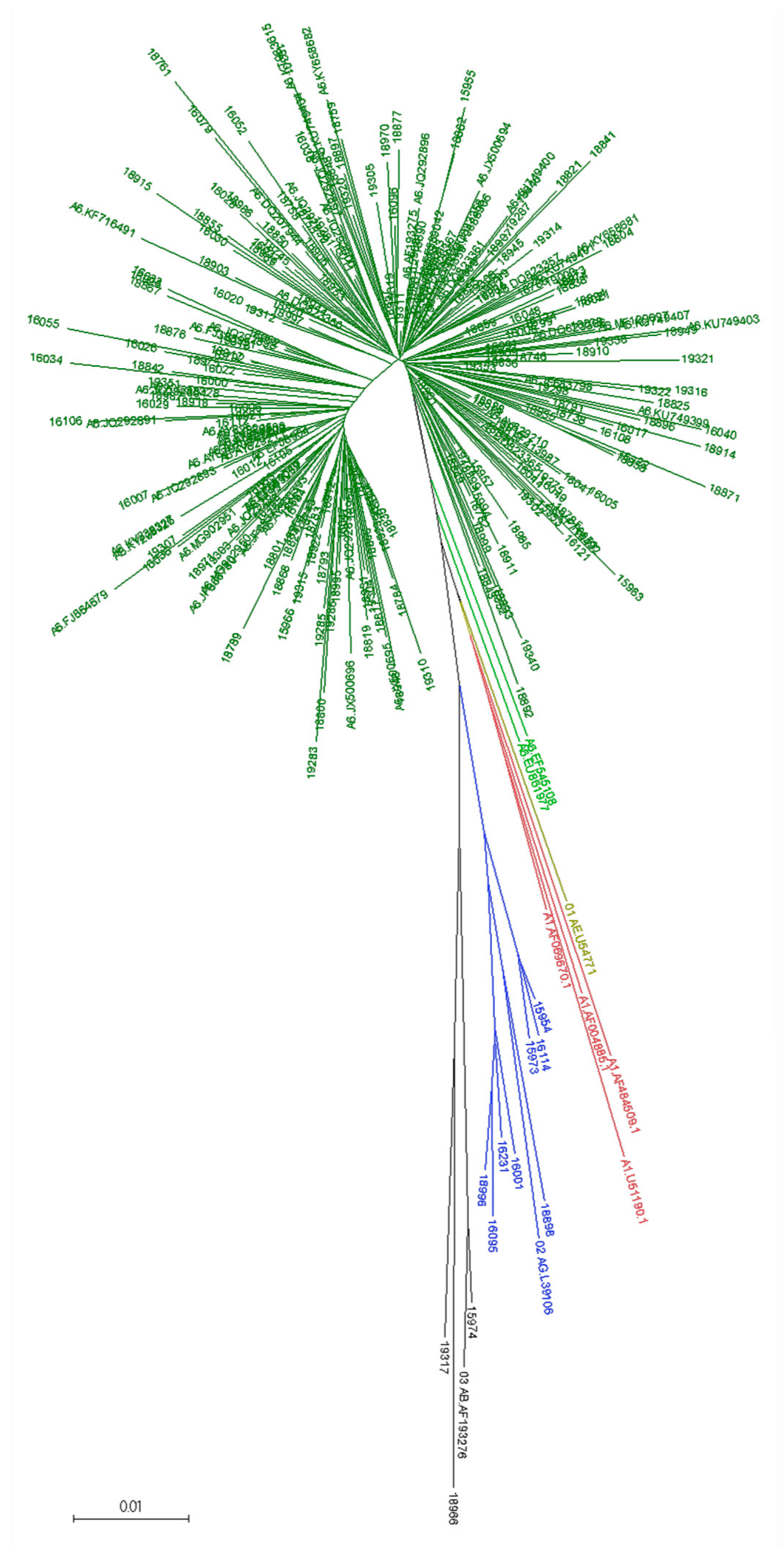
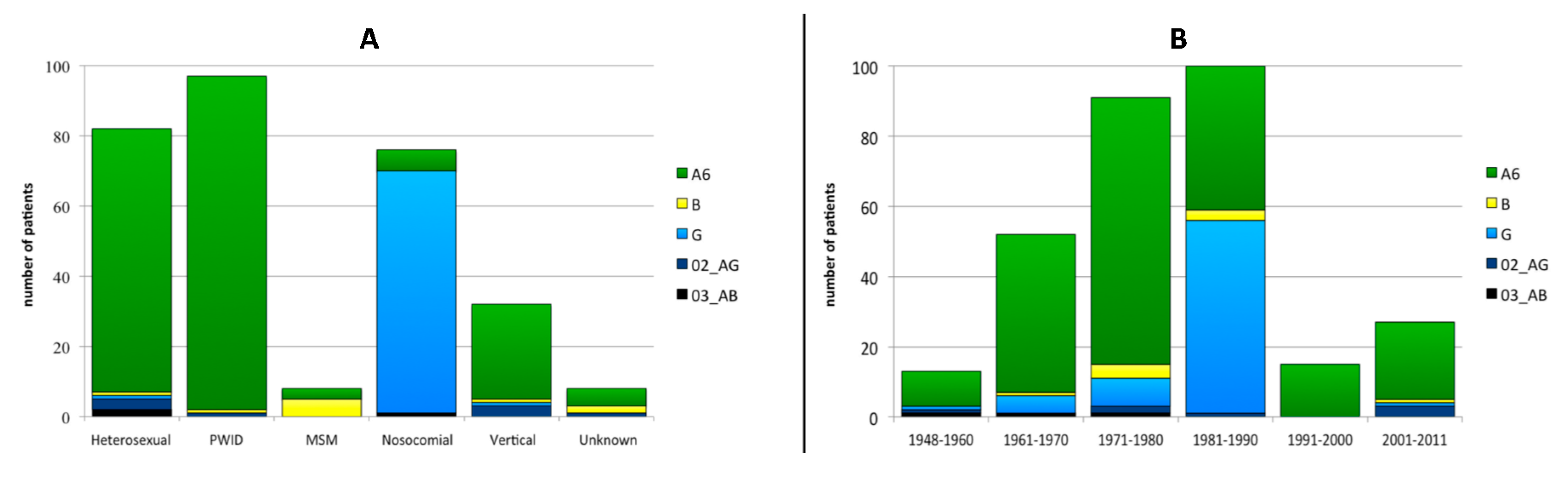
3.2. Viral Subtype Distribution
3.3. Resistance-Associated-Mutations (RAMs) and Drug-Susceptibility Profiles
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNAIDS DATA 2019. Available online: https://www.unaids.org/sites/default/files/media_asset/2019-UNAIDS-data_en.pdf (accessed on 2 February 2019).
- Beyrer, C.; Wirtz, A.L.; O’Hara, G.; Leon, N.; Kazatchkine, M. The expanding epidemic of HIV-1 in the Russian Federation. PLoS Med. 2017, 14, e1002462. [Google Scholar] [CrossRef]
- Bobkova, M. Current status of HIV-1 diversity and drug resistance monitoring in the former USSR. AIDS Rev. 2013, 15, 204–212. [Google Scholar] [PubMed]
- GBD 2017 HIV collaborators. Global, regional, and national incidence, prevalence, and mortality of HIV, 1980–2017, and forecasts to 2030, for 195 countries and territories: A systematic analysis for the Global Burden of Diseases, Injuries, and Risk Factors Study 2017. Lancet HIV 2019, 6, e831–e859. [Google Scholar] [CrossRef]
- Diez-Fuertes, F.; Cabello, M.; Thomson, M.M. Bayesian phylogeographic analyses clarify the origin of the HIV-1 subtype A variant circulating in former Soviet Union’s countries. Infect Genet. Evol. 2015, 33, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Novitsky, V.A.; Montano, M.A.; Essex, M. Molecular epidemiology of an HIV-1 subtype A subcluster among injection drug users in the Southern Ukraine. AIDS Res. Hum. Retrovir. 1998, 14, 1079–1085. [Google Scholar] [CrossRef]
- Thomson, M.M.; de Parga, E.V.; Vinogradova, A.; Sierra, M.; Yakovlev, A.; Rakhmanova, A.; Delgado, E.; Casado, G.; Munoz, M.; Carmona, R.; et al. New insights into the origin of the HIV type 1 subtype A epidemic in former Soviet Union’s countries derived from sequence analyses of preepidemically transmitted viruses. AIDS Res. Hum. Retrovir. 2007, 23, 1599–1604. [Google Scholar] [CrossRef]
- Bobkov, A.; Cheingsong-Popov, R.; Selimova, L.; Ladnaya, N.; Kazennova, E.; Kravchenko, A.; Fedotov, E.; Saukhat, S.; Zverev, S.; Pokrovsky, V.; et al. An HIV type 1 epidemic among injecting drug users in the former Soviet Union caused by a homogeneous subtype a strain. AIDS Res. Hum. Retrovir. 1997, 13, 1195–1201. [Google Scholar] [CrossRef]
- Rumyantseva, O.A.; Olkhovskiy, I.A.; Malysheva, M.A.; Ruzaeva, L.A.; Vasiliev, A.V.; Kazennova, E.V.; Bobkova, M.R.; Lukashov, V.V. Epidemiological networks and drug resistance of HIV type 1 in Krasnoyarsk region, Russia. AIDS Res. Hum. Retrovir. 2009, 25, 931–936. [Google Scholar] [CrossRef]
- Karkashadze, E.; Dvali, N.; Bolokadze, N.; Sharvadze, L.; Gabunia, P.; Karchava, M.; Tchelidze, T.; Tsertsvadze, T.; DeHovitz, J.; Del Rio, C.; et al. Epidemiology of human immunodeficiency virus (HIV) drug resistance in HIV patients with virologic failure of first-line therapy in the country of Georgia. J. Med. Virol. 2019, 91, 235–240. [Google Scholar] [CrossRef]
- Foley, B.T.; Leitner, T.; Paraskevis, D.; Peeters, M. Primate immunodeficiency virus classification and nomenclature: Review. Infect Genet. Evol. 2016, 46, 150–158. [Google Scholar] [CrossRef]
- Masharsky, A.E.; Klimov, N.A.; Kozlov, A.P. Molecular cloning and analysis of full-length genome of HIV type 1 strains prevalent in countries of the former Soviet Union. AIDS Res. Hum. Retrovir. 2003, 19, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Lapovok, I.; Laga, V.; Kazennova, E.; Bobkova, M. HIV Type 1 Integrase Natural Polymorphisms in Viral Variants Circulating in FSU Countries. Curr. HIV Res. 2017, 15, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Riva, C.; Romano, L.; Saladini, F.; Lai, A.; Carr, J.K.; Francisci, D.; Balotta, C.; Zazzi, M. Identification of a possible ancestor of the subtype A1 HIV Type 1 variant circulating in the former Soviet Union. AIDS Res. Hum. Retrovir. 2008, 24, 1319–1325. [Google Scholar] [CrossRef] [PubMed]
- Lapovok, I.A.; Lopatukhin, A.E.; Kireev, D.E.; Kazennova, E.V.; Lebedev, A.V.; Bobkova, M.R.; Kolomeets, A.N.; Turbina, G.I.; Shipulin, G.A.; Ladnaya, N.N.; et al. Molecular epidemiological analysis of HIV-1 variants circulating in Russia in 1987-2015. Ter. Arkh. 2017, 89, 44–49. [Google Scholar] [CrossRef]
- Pokrovskii, V.V.; Eramova, I.; Deulina, M.O.; Lipetikov, V.V.; Iashkulov, K.B.; Sliusareva, L.A.; Chemizova, N.M.; Savchenko, S.P. An intrahospital outbreak of HIV infection in Elista. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 1990, 17–23. [Google Scholar]
- Bobkov, A.; Cheingsong-Popov, R.; Garaev, M.; Rzhaninova, A.; Kaleebu, P.; Beddows, S.; Bachmann, M.H.; Mullins, J.I.; Louwagie, J.; Janssens, W.; et al. Identification of an env G subtype and heterogeneity of HIV-1 strains in the Russian Federation and Belarus. AIDS 1994, 8, 1649–1655. [Google Scholar] [CrossRef]
- Bobkov, A.; Garaev, M.M.; Rzhaninova, A.; Kaleebu, P.; Pitman, R.; Weber, J.N.; Cheingsong-Popov, R. Molecular epidemiology of HIV-1 in the former Soviet Union: Analysis of env V3 sequences and their correlation with epidemiologic data. AIDS 1994, 8, 619–624. [Google Scholar] [CrossRef]
- Murzakova, A.; Kireev, D.; Baryshev, P.; Lopatukhin, A.; Serova, E.; Shemshura, A.; Saukhat, S.; Kolpakov, D.; Matuzkova, A.; Suladze, A.; et al. Molecular Epidemiology of HIV-1 Subtype G in the Russian Federation. Viruses 2019, 11, 348. [Google Scholar] [CrossRef]
- Sierra, S.; Kaiser, R.; Lubke, N.; Thielen, A.; Schuelter, E.; Heger, E.; Daumer, M.; Reuter, S.; Esser, S.; Fatkenheuer, G.; et al. Prediction of HIV-1 coreceptor usage (tropism) by sequence analysis using a genotypic approach. J. Vis. Exp. 2011, 58, e3264. [Google Scholar] [CrossRef]
- Leitner, T.; Korber, B.; Daniels, M.; Calef, C.; Foley, B. HIV-1 Subtype and Circulating Recombinant Form (CRF) Reference Sequences. HIV Seq. Compend. 2005, 2005, 41–48. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.F.; Shafer, R.W. Web Resources for HIV type 1 Genotypic-Resistance Test Interpretation. Clin. Infect. Dis. 2006, 42, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Aibekova, L.; Foley, B.; Hortelano, G.; Raees, M.; Abdraimov, S.; Toichuev, R.; Ali, S. Molecular epidemiology of HIV-1 subtype A in former Soviet Union countries. PLoS ONE 2018, 13, e0191891. [Google Scholar] [CrossRef] [PubMed]
- Kazennova, E.; Laga, V.; Gromov, K.; Lebedeva, N.; Zhukova, E.; Pronin, A.; Grezina, L.; Dement’eva, N.; Shemshura, A.; Bobkova, M. Genetic Variants of HIV Type 1 in Men Who Have Sex with Men in Russia. AIDS Res. Hum. Retrovir. 2017, 33, 1061–1064. [Google Scholar] [CrossRef] [PubMed]
- Foley, B.; Leitner, T.; Apetrei, C.; Hahn, B.; Mizrachi, I.; Mullins, J.; Rambaut, A.; Wolinsky, S.; Korber, B. HIV Sequence Compendium 2018; Theoretical Biology and Biophysics Group T-6, Los Alamos National Laboratory: New Mexico, NM, USA, 2018. [Google Scholar]
- Miri, L.; Wakrim, L.; Kassar, H.; Hemminki, K.; Khyatti, M. Impact of immigration on HIV-1 molecular epidemiology in West Africa, Maghreb and Southern Europe. AIDS Rev. 2014, 16, 109–116. [Google Scholar]
- Perez-Parra, S.; Chueca, N.; Alvarez, M.; Pasquau, J.; Omar, M.; Collado, A.; Vinuesa, D.; Lozano, A.B.; Yebra, G.; Garcia, F. High prevalence and diversity of HIV-1 non-B genetic forms due to immigration in southern Spain: A phylogeographic approach. PLoS ONE 2017, 12, e0186928. [Google Scholar] [CrossRef]
- Pineda-Peña, A.C.; Pingarilho, M.; Li, G.; Vrancken, B.; Libin, P.; Gomes, P.; Camacho, R.J.; Theys, K.; Barroso Abecasis, A.; Portuguese, H.I.V.R.S.G. Drivers of HIV-1 transmission: The Portuguese case. PLoS ONE 2019, 14, e0218226. [Google Scholar] [CrossRef]
- Heipertz, R.A., Jr.; Ayemoba, O.; Sanders-Buell, E.; Poltavee, K.; Pham, P.; Kijak, G.H.; Lei, E.; Bose, M.; Howell, S.; O’Sullivan, A.M.; et al. Significant contribution of subtype G to HIV-1 genetic complexity in Nigeria identified by a newly developed subtyping assay specific for subtype G and CRF02_AG. Medicine (Baltimore) 2016, 95, e4346. [Google Scholar] [CrossRef]
- Gonzalez-Alba, J.M.; Holguin, A.; Garcia, R.; Garcia-Bujalance, S.; Alonso, R.; Suarez, A.; Delgado, R.; Cardenoso, L.; Gonzalez, R.; Garcia-Bermejo, I.; et al. Molecular surveillance of HIV-1 in Madrid, Spain: A phylogeographic analysis. J. Virol. 2011, 85, 10755–10763. [Google Scholar] [CrossRef]
- Karamov, E.; Epremyan, K.; Siniavin, A.; Zhernov, Y.; Cuevas, M.T.; Delgado, E.; Sanchez-Martinez, M.; Carrera, C.; Kornilaeva, G.; Turgiev, A.; et al. HIV-1 Genetic Diversity in Recently Diagnosed Infections in Moscow: Predominance of AFSU, Frequent Branching in Clusters, and Circulation of the Iberian Subtype G Variant. AIDS Res. Hum. Retrovir. 2018, 34, 629–634. [Google Scholar] [CrossRef]
- Sarabia, I.; Bosque, A. HIV-1 Latency and Latency Reversal: Does Subtype Matter? Viruses 2019, 11, 1104. [Google Scholar] [CrossRef] [PubMed]
- Daumer, M.; Awerkiew, S.; Aragon, S.S.; Kartashev, V.; Poplavskaja, T.; Klein, R.; Sichtig, N.; Thiele, B.; Lengauer, T.; Roomp, K.; et al. Short communication: Selection of thymidine analogue resistance mutational patterns in children infected from a common HIV type 1 subtype G source. AIDS Res. Hum. Retrovir. 2010, 26, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Wainberg, M.A.; Brenner, B.G. Role of HIV Subtype Diversity in the Development of Resistance to Antiviral Drugs. Viruses 2010, 2, 2493–2508. [Google Scholar] [CrossRef]
- Maldonado, J.O.; Mansky, L.M. The HIV-1 Reverse Transcriptase A62V Mutation Influences Replication Fidelity and Viral Fitness in the Context of Multi-Drug-Resistant Mutations. Viruses 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Dvali, N.; Parker, M.M.; Chkhartishvili, N.; Sharvadze, L.; Gochitashvili, N.; Abutidze, A.; Karchava, M.; DeHovitz, J.A.; Tsertsvadze, T. Characterization of HIV-1 subtypes and drug resistance mutations among individuals infected with HIV in Georgia. J. Med. Virol. 2012, 84, 1002–1008. [Google Scholar] [CrossRef]
- Kolomeets, A.N.; Varghese, V.; Lemey, P.; Bobkova, M.R.; Shafer, R.W. A uniquely prevalent nonnucleoside reverse transcriptase inhibitor resistance mutation in Russian subtype A HIV-1 viruses. AIDS 2014, 28, F1–F8. [Google Scholar] [CrossRef]
- Bacheler, L.T.; Anton, E.D.; Kudish, P.; Baker, D.; Bunville, J.; Krakowski, K.; Bolling, L.; Aujay, M.; Wang, X.V.; Ellis, D.; et al. Human immunodeficiency virus type 1 mutations selected in patients failing efavirenz combination therapy. Antimicrob. Agents Chemother. 2000, 44, 2475–2484. [Google Scholar] [CrossRef]
- Huang, W.; Gamarnik, A.; Limoli, K.; Petropoulos, C.J.; Whitcomb, J.M. Amino acid substitutions at position 190 of human immunodeficiency virus type 1 reverse transcriptase increase susceptibility to delavirdine and impair virus replication. J. Virol. 2003, 77, 1512–1523. [Google Scholar] [CrossRef]
- Brenner, B.G. Selective acquisition of G190S in HIV-1 subtype A from Russia leading to efavirenz and nevirapine treatment failure. AIDS 2014, 28, 2619–2621. [Google Scholar] [CrossRef]
- Avidor, B.; Turner, D.; Mor, Z.; Chalom, S.; Riesenberg, K.; Shahar, E.; Pollack, S.; Elbirt, D.; Sthoeger, Z.; Maayan, S.; et al. Transmission patterns of HIV-subtypes A/AE versus B: Inferring risk-behavior trends and treatment-efficacy limitations from viral genotypic data obtained prior to and during antiretroviral therapy. PLoS ONE 2013, 8, e57789. [Google Scholar] [CrossRef] [PubMed]
- Dvali, N.; Chkhartishvili, N.; Karchava, M.; Sharvadze, L.; Tsertsvadze, T. Distinct drug resistance profile of HIV-1 subtype A strain circulating in Georgia. Georgian Med. News 2015, 19–24. [Google Scholar]
- Davy-Mendez, T.; Eron, J.J.; Brunet, L.; Zakharova, O.; Dennis, A.M.; Napravnik, S. New antiretroviral agent use affects prevalence of HIV drug resistance in clinical care populations. AIDS 2018, 32, 2593–2603. [Google Scholar] [CrossRef] [PubMed]
- Schultze, A.; Phillips, A.N.; Paredes, R.; Battegay, M.; Rockstroh, J.K.; Machala, L.; Tomazic, J.; Girard, P.M.; Januskevica, I.; Gronborg-Laut, K.; et al. HIV resistance testing and detected drug resistance in Europe. AIDS 2015, 29, 1379–1389. [Google Scholar] [CrossRef]
- Knops, E.; Schuülter, E.; Luübke, N.; Oette, M.; Faätkenheuer, G.; Hower, M.; Knechten, H.; Mutz, A.; Esser, S.; Scholten, S.; et al. The RESINA data support the individualized therapy based on primary resistance testing. Eur. Resist. Abstr. 67 2015, 2015, 66–67. [Google Scholar]
- Franzetti, M.; De Luca, A.; Ceccherini-Silberstein, F.; Spagnuolo, V.; Nicastri, E.; Mussini, C.; Antinori, A.; Monno, L.; Vecchiet, J.; Fanti, I.; et al. Evolution of HIV-1 transmitted drug resistance in Italy in the 2007-2014 period: A weighted analysis. J. Clin. Virol. 2018, 106, 49–52. [Google Scholar] [CrossRef]
- Hauser, A.; Hofmann, A.; Hanke, K.; Bremer, V.; Bartmeyer, B.; Kuecherer, C.; Bannert, N. National molecular surveillance of recently acquired HIV infections in Germany, 2013 to 2014. Euro Surveill 2017, 22, 30436. [Google Scholar] [CrossRef]
- Rossetti, B.; Di Giambenedetto, S.; Torti, C.; Postorino, M.C.; Punzi, G.; Saladini, F.; Gennari, W.; Borghi, V.; Monno, L.; Pignataro, A.R.; et al. Evolution of transmitted HIV-1 drug resistance and viral subtypes circulation in Italy from 2006 to 2016. HIV Med. 2018, 19, 619–628. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| RAMs | Patients | ||||
|---|---|---|---|---|---|
| Total 1 | TN 2 | TE 3 | No Data 4 | p Value 5 | |
| PI | 261 | 102 | 110 | 49 | |
| D30N | 1 (0.4%) | 0 (0.0%) | 1 (0.9%) | 0 (0.0%) | ≥0.05 |
| M46IL | 13 (5.0%) | 2 (2.0%) | 9 (8.2%) | 2 (4.1%) | ≥0.05 |
| G48V | 2 (0.8%) | 2 (2.0%) | 0 (0.0%) | 0 (0.0%) | ≥0.05 |
| I50VL | 4 (1.5%) | 0 (0.0%) | 3 (2.7%) | 1 (2.0%) | ≥0.05 |
| I54AV | 8 (3.1%) | 1 (1.0%) | 6 (5.5%) | 1 (2.0%) | ≥0.05 |
| L76V | 2 (0.8%) | 1 (1.0%) | 1 (0.9%) | 0 (0.0%) | ≥0.05 |
| V82AFMST | 10 (3.8%) | 1 (1.0%) | 8 (7.3%) | 1 (2.0%) | 0,0363 |
| N88S | 1 (0.4%) | 0 (0.0%) | 0 (0.0%) | 1 (2.0%) | ≥0.05 |
| L90M | 10 (3.8%) | 1 (1.0%) | 8 (7.3%) | 1 (2.0%) | 0,0363 |
| NRTIs | 277 | 109 | 120 | 48 | |
| M41L | 22 (7.9%) | 1 (0.9%) | 19 (15.8%) | 2 (4.2%) | <0.0001 |
| E44D | 1 (0.4%) | 0 (0.0%) | 1 (0.8%) | 0 (0.0%) | ≥0.05 |
| A62V | 61 (22.0%) | 30 (27.5%) | 22 (18.3%) | 9 (18.8%) | ≥0.05 |
| D67G | 3 (1.1%) | 1 (0.9%) | 2 (1.7%) | 0 (0.0%) | ≥0.05 |
| D67N | 17 (6.1%) | 1 (0.9%) | 13 (10.8%) | 3 (6.3%) | 0.0016 |
| K65DEN | 4 (1.4%) | 1 (0.9%) | 3 (2.5%) | 0 (0.0%) | ≥0.05 |
| T69DN | 3 (1.1%) | 0 (0.0%) | 3 (2.5%) | 0 (0.0%) | ≥0.05 |
| T69G_SG | 1 (0.4%) | 0 (0.0%) | 1 (0.8%) | 0 (0.0%) | ≥0.05 |
| K70ER | 14 (5.1%) | 1 (0.9%) | 11 (9.2%) | 2 (4.2%) | 0.0058 |
| L74IV | 9 (3.2%) | 0 (0.0%) | 6 (5.0%) | 3 (6.3%) | ≥0.05 |
| V75AIM | 5 (1.8%) | 0 (0.0%) | 5 (4.2%) | 0 (0.0%) | ≥0.05 |
| Y115F | 2 (0.7%) | 0 (0.0%) | 1 (0.8%) | 1 (2.1%) | ≥0.05 |
| M184IV | 60 (21.7%) | 5 (4.6%) | 44 (36.7%) | 11 (22.9%) | <0.0001 |
| L210W | 8 (2.9%) | 0 (0.0%) | 8 (6.7%) | 0 (0.0%) | 0.0073 |
| T215CFILNSY | 38 (13.7%) | 1 (0.9%) | 33 (27.5%) | 4 (8.3%) | <0.0001 |
| K219ENQ | 16 (5.8%) | 1 (0.9%) | 12 (10.0%) | 3 (6.3%) | 0.0030 |
| NNRTI | 277 | 109 | 120 | 48 | |
| A98G | 2 (0.7%) | 0 (0.0%) | 2 (1.7%) | 0 (0.0%) | ≥0.05 |
| K101EHPQ | 11 (4.0%) | 0 (0.0%) | 9 (7.5%) | 2 (4.2%) | 0.0036 |
| L100F | 2 (0.7%) | 0 (0.0%) | 2 (1.7%) | 0 (0.0%) | ≥0.05 |
| K103NS | 28 (10.1%) | 0 (0.0%) | 24 (20.0%) | 4 (8.3%) | <0.0001 |
| V106A | 1 (0.4%) | 0 (0.0%) | 1 (0.8%) | 0 (0.0%) | ≥0.05 |
| V108I | 6 (2.2%) | 0 (0.0%) | 3 (2.5%) | 3 (6.3%) | ≥0.05 |
| E138AGHKQR | 20 (7.2%) | 5 (4.6%) | 13 (10.8%) | 2 (4.2%) | ≥0.05 |
| V179DE | 6 (2.2%) | 2 (1.8%) | 4 (3.3%) | 0 (0.0%) | ≥0.05 |
| Y181CFIV | 13 (4.7%) | 0 (0.0%) | 11 (9.2%) | 2 (4.2%) | 0.0009 |
| Y188LS | 4 (1.4%) | 1 (0.9%) | 3 (2.5%) | 0 (0.0%) | ≥0.05 |
| G190AS | 21 (7.6%) | 0 (0.0%) | 18 (15.0%) | 3 (6.3%) | <0.0001 |
| H221Y | 2 (0.7%) | 0 (0.0%) | 2 (1.7%) | 0 (0.0%) | ≥0.05 |
| P225H | 4 (1.4%) | 0 (0.0%) | 3 (2.5%) | 1 (2.1%) | ≥0.05 |
| F227L | 1 (0.4%) | 0 (0.0%) | 1 (0.8%) | 0 (0.0%) | ≥0.05 |
| M230L | 1 (0.4%) | 0 (0.0%) | 1 (0.8%) | 0 (0.0%) | ≥0.05 |
| K238T | 1 (0.4%) | 0 (0.0%) | 0 (0.0%) | 1 (2.1%) | ≥0.05 |
| Y318F | 1 (0.4%) | 0 (0.0%) | 1 (0.8%) | 0 (0.0%) | ≥0.05 |
| INSTI | 61 | 19 | 21 | 21 | |
| T6TK | 1 (1.6%) | 0 (0.0%) | 1 (4.8%) | 0 (0.0%) | ≥0.05 |
| G140A | 1 (1.6%) | 0 (0.0%) | 1 (4.8%) | 0 (0.0%) | ≥0.05 |
| Q148R | 1 (1.6%) | 0 (0.0%) | 1 (4.8%) | 0 (0.0%) | ≥0.05 |
| N155H | 1 (1.6%) | 0 (0.0%) | 1 (4.8%) | 0 (0.0%) | ≥0.05 |
| Patients | |||||
|---|---|---|---|---|---|
| Total 1 | TN 2 | TE 3 | No Data 4 | ||
| Available PR Sequences | 261 | 102 | 110 | 49 | |
| PI | IR | 11 (4.2%) | 6 (5.9%) | 5 (4.5%) | 0 (0.0%) |
| FR | 21 (8.0%) | 3 (2.9%) | 14 (12.7%) | 4 (8.2%) | |
| CR-resistance | 32 (12.3%) | 9 (8.8%) | 19 (17.3%) | 4 (8.2%) | |
| Available RT Sequences | 277 | 109 | 120 | 48 | |
| NRTI | IR | 9 (3.2%) | 2 (1.8%) | 6 (5.0%) | 1 (2.1%) |
| FR | 72 (26.0%) | 5 (4.6%) | 54 (45.0%) | 13 (27.1%) | |
| CR-resistance | 81 (29.2%) | 7 (6.4%) | 60 (50.0%) | 14 (29.2%) | |
| NNRTI | IR | 2 (0.7%) | 0 (0.0%) | 1 (0.8%) | 1 (2.1%) |
| FR | 54 (19.5%) | 0 (0.0%) | 46 (38.3%) | 8 (16.7%) | |
| CR-resistance | 56 (20.2%) | 0 (0.0%) | 47 (39.2%) | 9 (18.8%) | |
| NRTI + NNRTI | IR | 3 (1.1%) | 0 (0.0%) | 2 (1.7%) | 1 (2.1%) |
| FR | 30 (10.83%) | 0 (0.0%) | 26 (21.7%) | 4 (8.3%) | |
| CR-resistance | 33 (11.9%) | 0 (0.0%) | 28 (23.3%) | 5 (10.4%) | |
| Available PR + RT Sequences | 260 | 102 | 109 | 49 | |
| PI + NRTI | IR | 1 (0.4%) | 1 (1.0%) | 0 (0.0%) | 0 (0.0%) |
| FR | 9 (3.5%) | 2 (2.0%) | 5 (4.6%) | 2 (4.1%) | |
| CR-resistance | 10 (3.8%) | 3 (2.9%) | 5 (4.6%) | 2 (4.1%) | |
| PI + NNRTI | IR | 1 (0.4%) | 0 (0.0%) | 1 (0.9%) | 0 (0.0%) |
| FR | 1 (0.4%) | 0 (0.0%) | 1 (0.9%) | 0 (0.0%) | |
| CR-resistance | 2 (0.8%) | 0 (0.0%) | 2 (1.8%) | 0 (0.0%) | |
| PI + NRTI + NNRTI | IR | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) |
| FR | 5 (1.9%) | 0 (0.0%) | 3 (2.8%) | 2 (4.1%) | |
| CR-resistance | 5 (1.9%) | 0 (0.0%) | 3 (2.8%) | 2 (4.1%) | |
| Available IN Sequences | 61 | 19 | 21 | 21 | |
| INSTI | IR | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) |
| FR | 2 (3.3%) | 0 (0.0%) | 2 (9.5%) | 2 (4.1%) | |
| CR-resistance | 3 (3.3%) | 0 (0.0%) | 2 (9.5%) | 2 (4.1%) | |
| Available RT + IN Sequences | 52 | 18 | 17 | 17 | |
| NRTI + NNRTI + INSTI | IR | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) |
| FR | 2 (3.8%) | 0 (0.0%) | 2 (11.8%) | 0 (0.0%) | |
| CR-resistance | 2 (3.8%) | 0 (0.0%) | 2 (11.8%) | 0 (0.0%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlösser, M.; Kartashev, V.V.; Mikkola, V.H.; Shemshura, A.; Saukhat, S.; Kolpakov, D.; Suladze, A.; Tverdokhlebova, T.; Hutt, K.; Heger, E.; et al. HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia. Viruses 2020, 12, 475. https://doi.org/10.3390/v12040475
Schlösser M, Kartashev VV, Mikkola VH, Shemshura A, Saukhat S, Kolpakov D, Suladze A, Tverdokhlebova T, Hutt K, Heger E, et al. HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia. Viruses. 2020; 12(4):475. https://doi.org/10.3390/v12040475
Chicago/Turabian StyleSchlösser, Madita, Vladimir V. Kartashev, Visa H. Mikkola, Andrey Shemshura, Sergey Saukhat, Dmitriy Kolpakov, Alexandr Suladze, Tatiana Tverdokhlebova, Katharina Hutt, Eva Heger, and et al. 2020. "HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia" Viruses 12, no. 4: 475. https://doi.org/10.3390/v12040475
APA StyleSchlösser, M., Kartashev, V. V., Mikkola, V. H., Shemshura, A., Saukhat, S., Kolpakov, D., Suladze, A., Tverdokhlebova, T., Hutt, K., Heger, E., Knops, E., Böhm, M., Di Cristanziano, V., Kaiser, R., Sönnerborg, A., Zazzi, M., Bobkova, M., & Sierra, S. (2020). HIV-1 Sub-Subtype A6: Settings for Normalised Identification and Molecular Epidemiology in the Southern Federal District, Russia. Viruses, 12(4), 475. https://doi.org/10.3390/v12040475