Metabolomic Analysis of Cricket paralysis virus Infection in Drosophila S2 Cells Reveals Divergent Effects on Central Carbon Metabolism as Compared with Silkworm Bm5 Cells

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
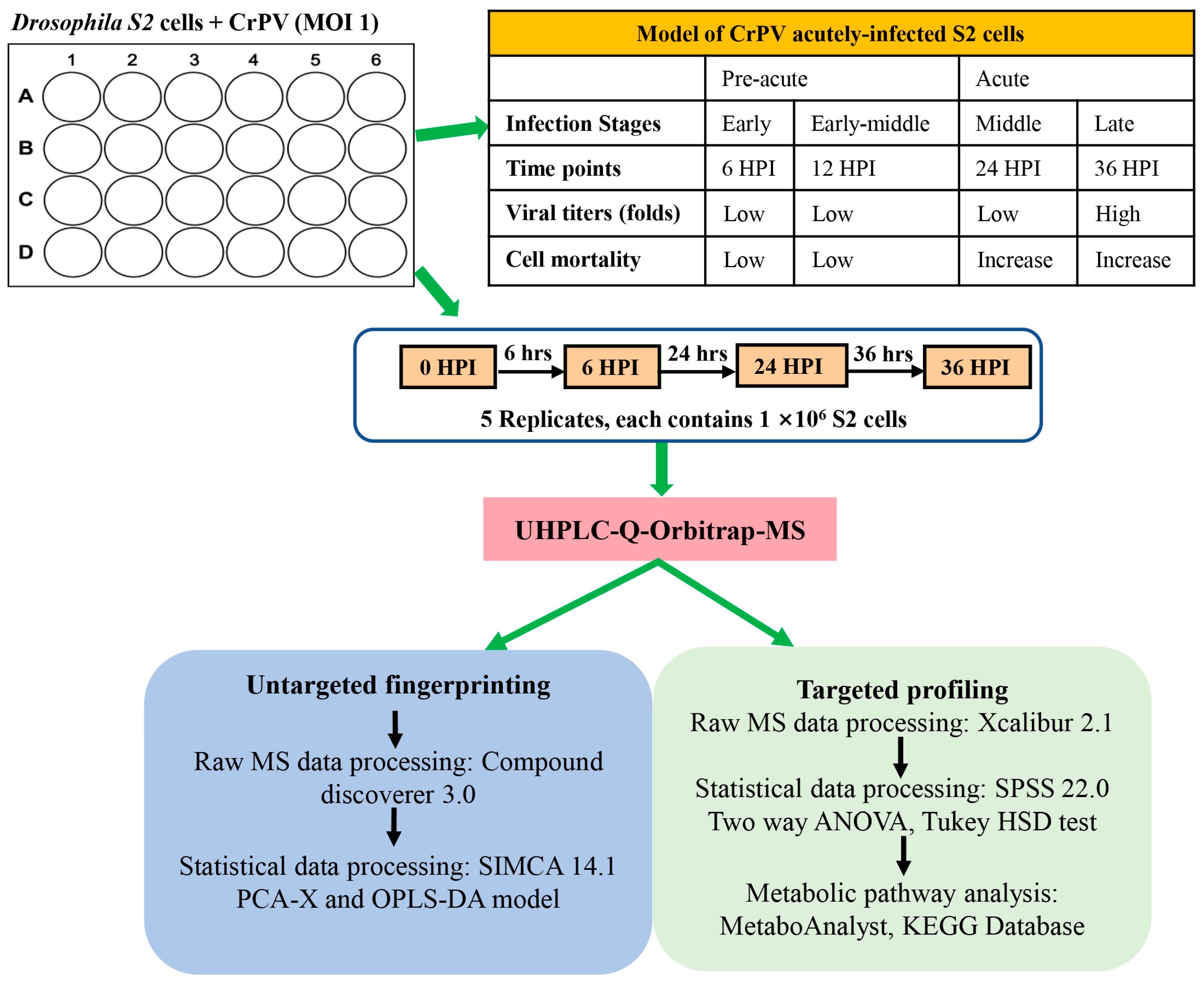
2.1. Cell Culture and Virus Infection
2.2. Experimental Design
2.3. Extraction of Intracellular Metabolites and Ultra-High Performance Liquid Chromatography Hyphenated to Quadrupole-Orbitrap High-Resolution Mass Spectrometry (UHPLC-Q-Orbitrap-HRMS) Analysis
2.4. Data Analysis
2.4.1. Untargeted Data Analysis
2.4.2. Targeted Data Analysis
3. Results
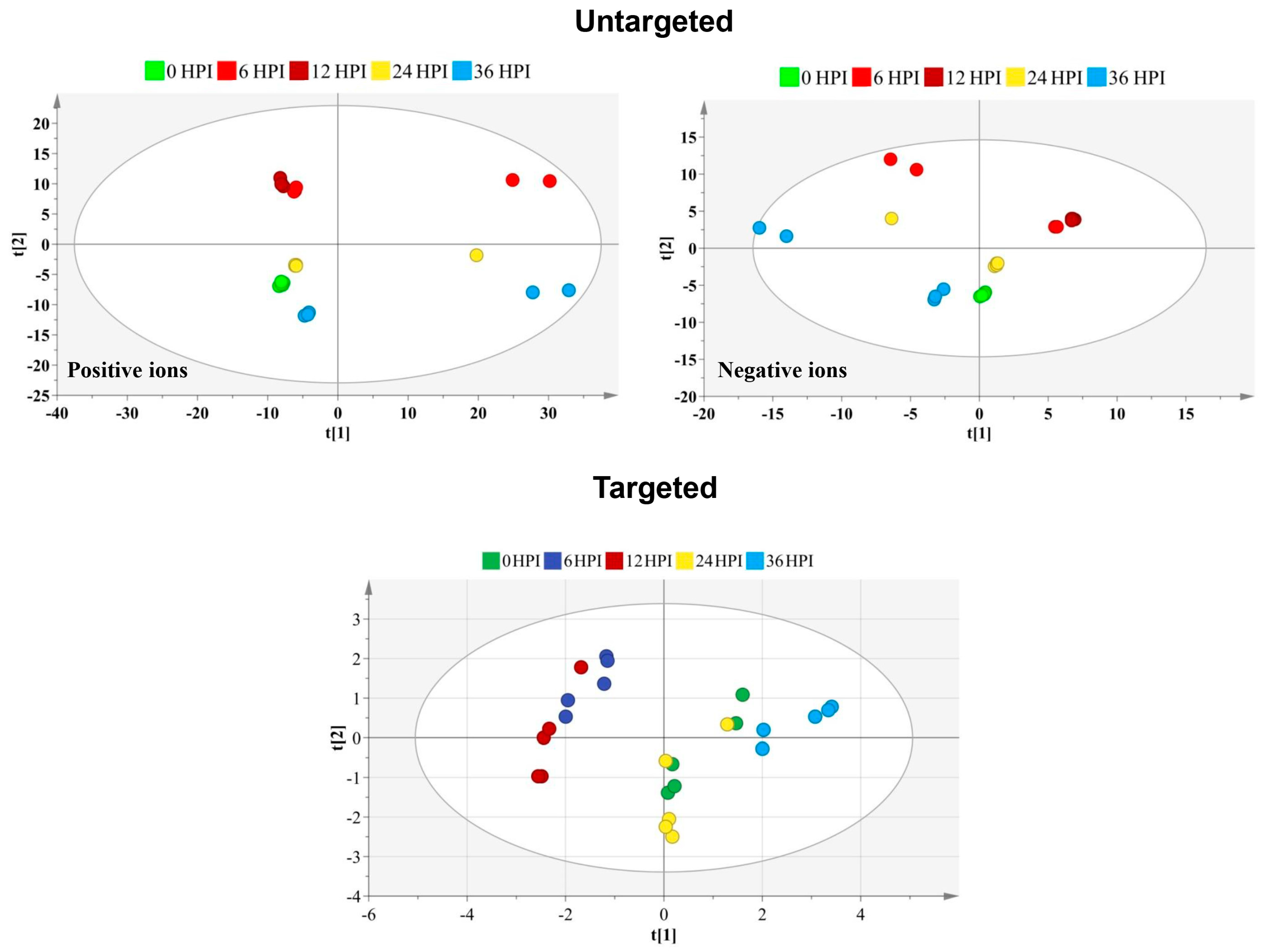
3.1. Global Metabolic Changes in S2 Cells Following CrPV Infection
3.2. Overview of Targeted Metabolite Profiling in S2 Cells Following CrPV Infection
3.3. Distinct Metabolome Changes Triggered by CrPV Infection in S2 Cells
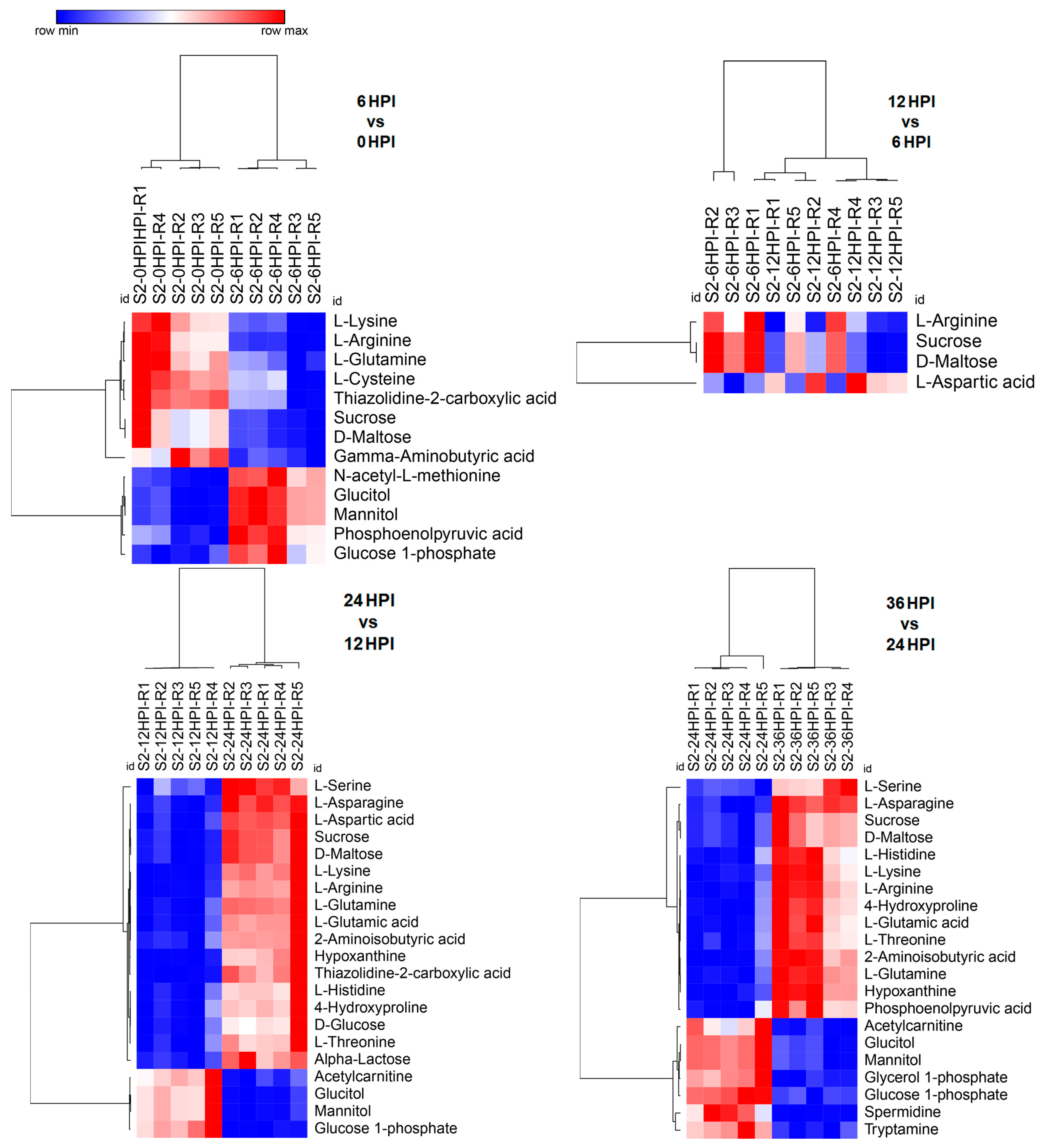
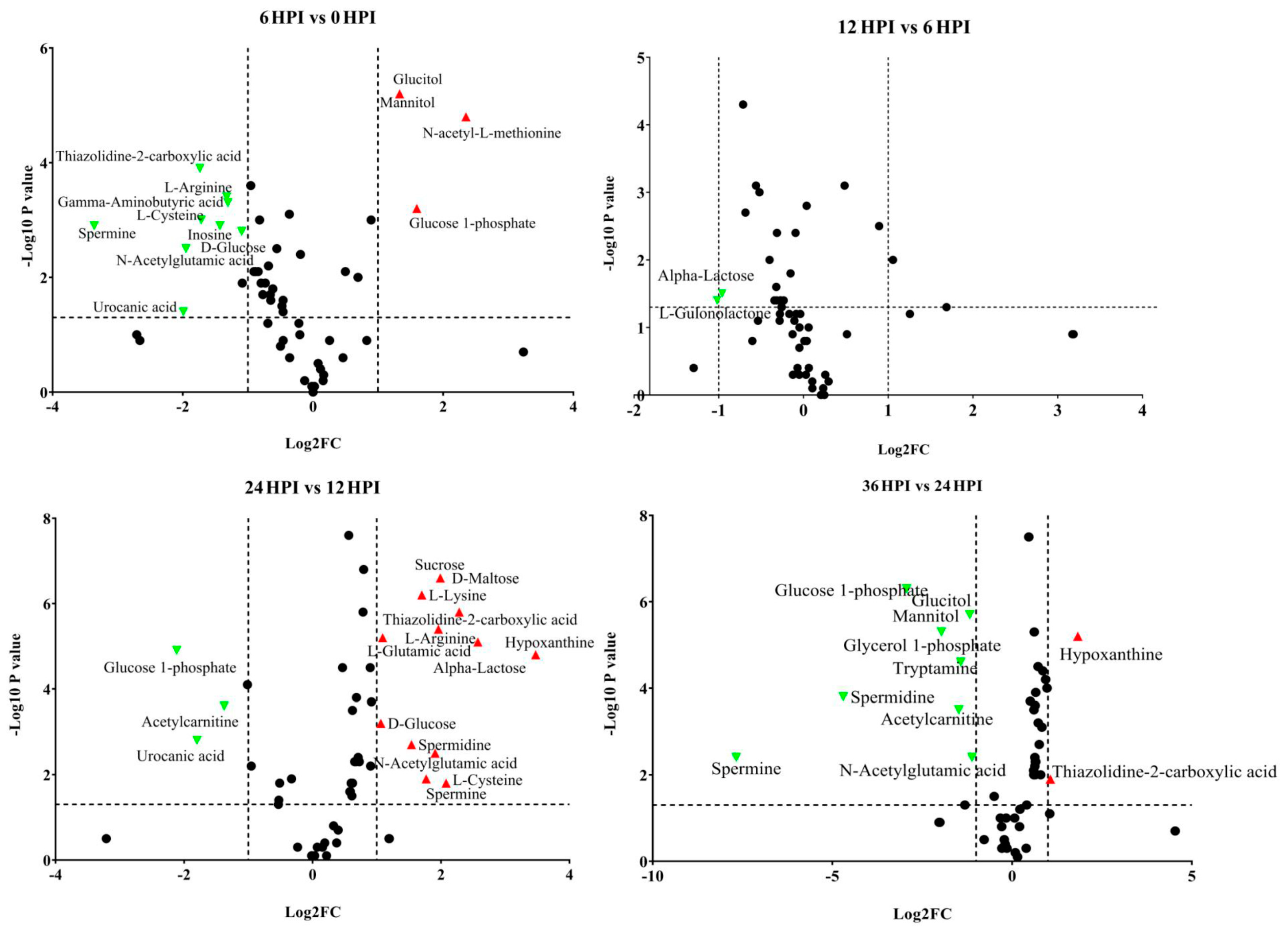
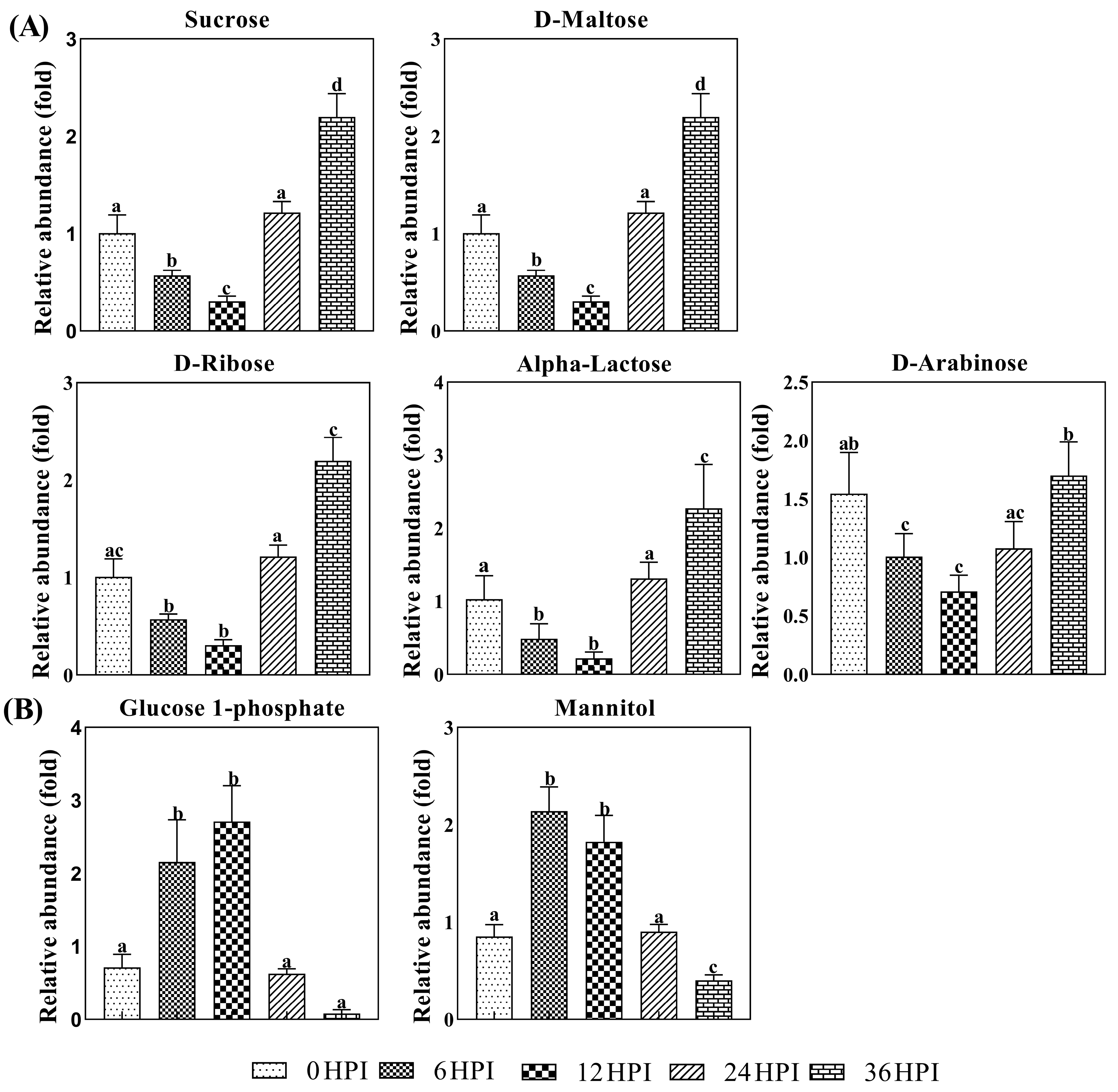
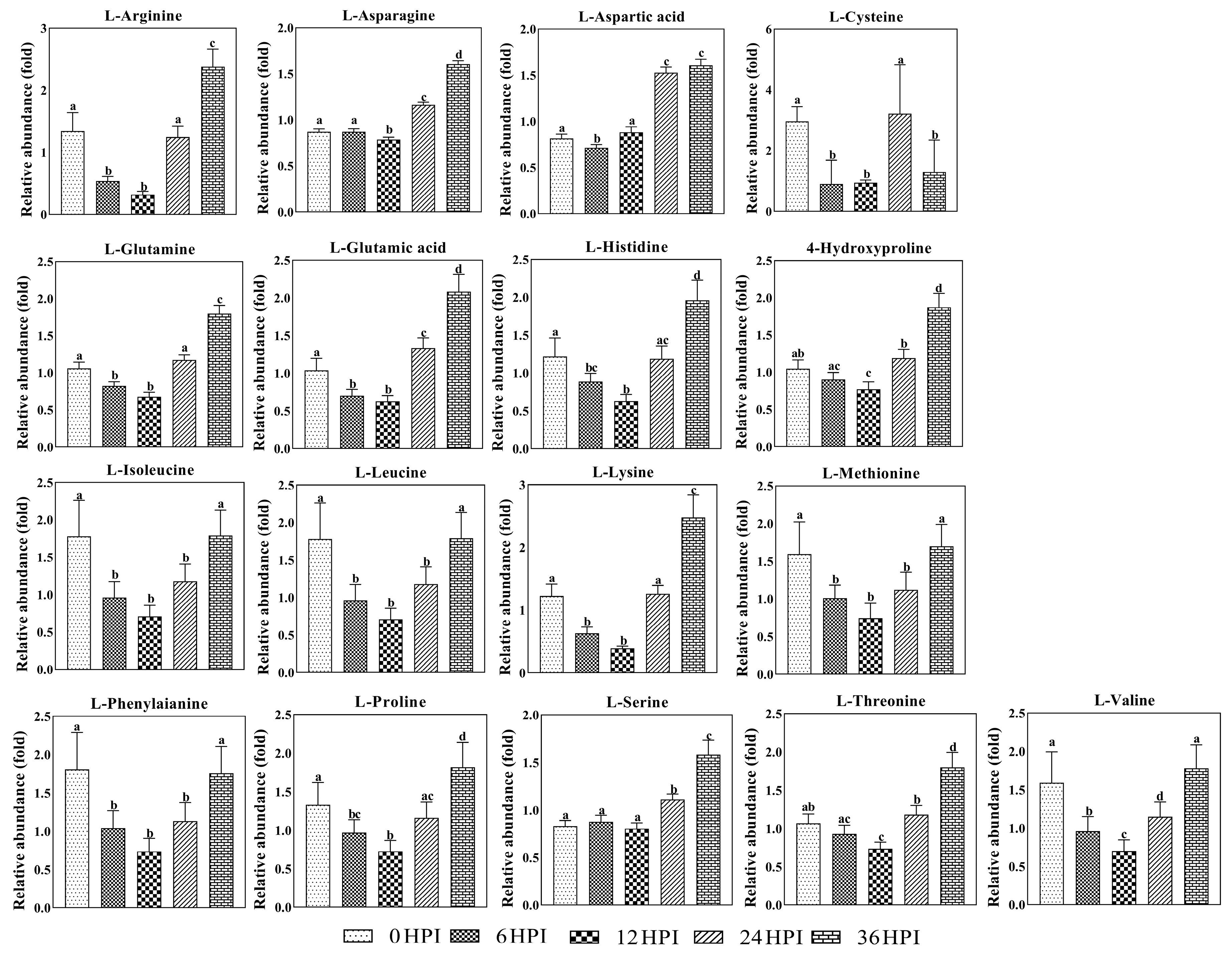
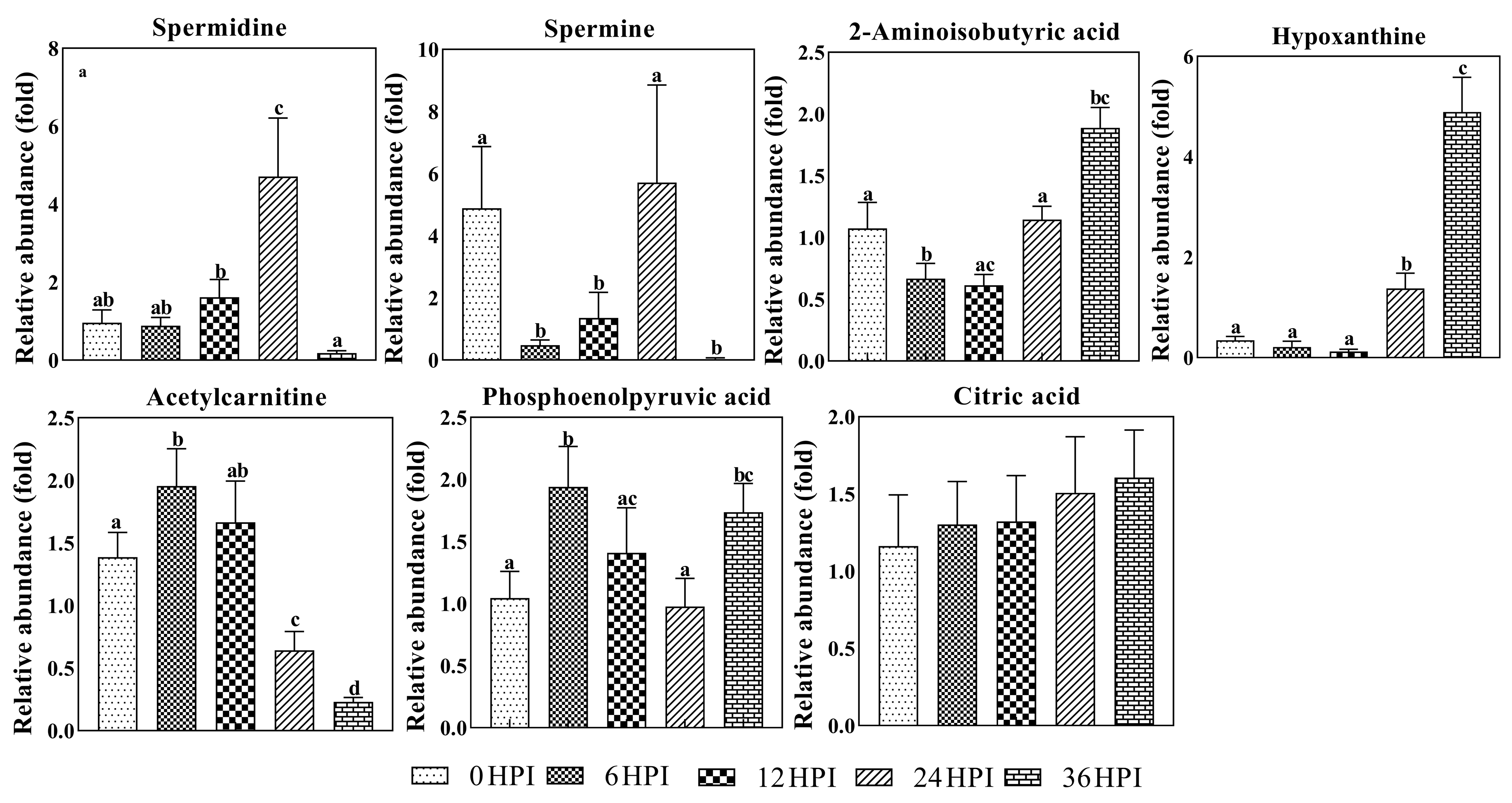
3.4. Abundance Analysis of Carbohydrates and Amino Acids during host-CrPV Infection
3.5. Key Metabolites Required for Acute Infection of CrPV in S2 Cells
3.6. Comparison of Metabolite Changes between Persistent Infection in Bm5 Cells and Acute Infection in S2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ertl, H.C.J. Viral Immunology. In Fundamental Immunology; Paul, W.E., Ed.; Wolters Kluwer: Alphen aan den Rijn, The Netherlands, 2012; pp. 937–972. [Google Scholar]
- Kane, M.; Golovkina, T. Common threads in persistent viral infections. J. Virol. 2010, 84, 4116–4123. [Google Scholar] [CrossRef] [PubMed]
- Arbour, N.; Côté, G.; Lachance, C.; Tardieu, M.; Cashman, N.R.; Talbot, P.J. Acute and persistent infection of human neural cell lines by human coronavirus OC43. J. Virol. 1999, 73, 3338–3350. [Google Scholar] [CrossRef] [PubMed]
- Sempere, R.N.; Arias, A. Establishment of a Cell Culture Model of Persistent Flaviviral Infection: Usutu Virus Shows Sustained Replication during Passages and Resistance to Extinction by Antiviral Nucleosides. Viruses 2019, 11, 560. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Canning, W.M.; Kauffman, R.S.; Sharpe, A.H.; Hallum, J.V.; Fields, B.N. Role of the Host Cell in Persistent Viral Infection: Coevolution of L Cells and Reovirus during Persistent Infection. Cell 1981, 25, 325–332. [Google Scholar] [CrossRef]
- Nomaguchi, M.; Fujita, M.; Miyasaki, Y.; Adachi, A. Viral tropism. Front Microbiol. 2012, 3, 281. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.; Kim, D.Y.; Trnka, M.J.; Kerr, C.H.; Lidsky, P.V.; Stanley, D.J.; Riversa, B.M.; Li, K.H.; Burlingame, A.L.; Jan, E.; et al. A Viral Protein Restricts Drosophila RNAi Immunity by Regulating Argonaute Activity and Stability. Cell Host Microbe 2018, 24, 542–557. [Google Scholar] [CrossRef]
- Merkling, S.H.; Riahi, H.; Overheul, G.J.; Schenck, A.; van Rij, R.P. Peroxisome-associated Sgroppino links fat metabolism with survival after RNA virus infection in Drosophila. Sci. Rep. 2019, 9, 2065. [Google Scholar] [CrossRef]
- Nayak, A.; Berry, B.; Tassetto, M.; Kunitomi, M.; Acevedo, A.; Deng, C.H.; Krutchinsky, A.; Gross, J.; Antoniewski, C.; Andino, R. Cricket paralysis virus antagonizes Argonaute 2 to modulate antiviral defense in Drosophila. Nat. Struct. Mol. Biol. 2010, 17, 547–554. [Google Scholar]
- Wang, L.; Cappelle, K.; Santos, D.; Vanden Broeck, J.; Smagghe, G.; Swevers, L. Short-term persistence precedes pathogenic infection: Infection kinetics of cricket paralysis virus in silkworm-derived Bm5 cells. J. Insect Physiol. 2019, 115, 1–11. [Google Scholar] [CrossRef]
- Santos, D.; Wynant, N.; Van den Brande, S.; Verdonckt, T.-W.; Mingels, L.; Peeters, P.; Kolliopoulou, A.; Swevers, L.; Vanden Broeck, J. Insights into RNAi-based antiviral immunity in Lepidoptera: acute and persistent infections in Bombyx mori and Trichoplusia ni cell lines. Sci. Rep. 2018, 8, 2423. [Google Scholar] [CrossRef]
- Santos, D.; Mingels, L.; Vogel, E.; Wang, L.; Christiaens, O.; Cappelle, K.; Wynant, N.; Gansemans, Y.; Van Nieuwerburgh, F.; Smagghe, G.; et al. Generation of virus- and dsRNA-derived siRNAs with species-dependent length in insects. Viruses 2019, 11, 738. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Swevers, L.; Rombouts, C.; Meeus, I.; Van Meulebroeck, L.; Vanhaecke, L.; Smagghe, G. A Metabolomics Approach to Unravel Cricket Paralysis Virus Infection in Silkworm Bm5 Cells. Viruses 2019, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, E.; Van Meulebroek, L.; Rombouts, C.; Huysman, S.; Verplanken, K.; Lapauw, B.; Wauters, J.; Hemeryck, L.Y.; Vanhaecke, L. A validated multi-matrix platform for metabolomic fingerprinting of human urine, feces and plasma using ultra-high performance liquid-chromatography coupled to hybrid orbitrap high-resolution mass spectrometry. Anal. Chim. Acta 2018, 1033, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Vanden Bussche, J.; Marzorati, M.; Laukens, D.; Vanhaecke, L. Validated high resolution mass spectrometry-based approach for metabolomic fingerprinting of the human gut phenotype. Anal. Chem. 2015, 87, 10927–10934. [Google Scholar] [CrossRef] [PubMed]
- Li, M.M.H.; MacDonald, M.R. Polyamines: small molecules with a big role in promoting virus infection. Cell Host & Microbe 2016, 20, 123–124. [Google Scholar]
- Mounce, B.C.; Olsen, M.E.; Vignuzzi, M.; Connor, J.H. Polyamines and their role in virus infection. Microbiol. Mol. Biol. Rev. 2017, 81, e00029-17. [Google Scholar] [CrossRef]
- Mendelson, S.D. Nutritional supplements and metabolic syndrome. In Metabolic syndrome and psychiatric illness—Interactions, pathophysiology, assessment and treatment; Mendelson, S.D., Ed.; Elsevier: Amsterdam, The Netherlands; pp. 141–186.
- Zerbe, R.L.; Robertson, G.L. Osmoregulation of thirst and vasopressin secretion in human subjects: effect of various solutes. Am. J. Physiol. 1983, 244, E607–E614. [Google Scholar] [CrossRef]
- Burg, M.B.; Kador, P.F. Sorbitol, osmoregulation, and the complications of diabetes. J. Clin. Invest. 1988, 81, 635–640. [Google Scholar] [CrossRef]
- Thoden, J.B.; Holden, H.M. The molecular architecture of glucose-1-phosphate uridylyltransferase. Protein Sci. 2007, 16, 432–440. [Google Scholar] [CrossRef]
- Bremer, J. The role of carnitine in intracellular metabolism. J. Clin. Chem. Clin. Biochem. 1990, 28, 297–301. [Google Scholar]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479–480, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Byers, N.M.; Fleshman, A.C.; Perera, R.; Molins, C.R. Metabolomic insights into human arboviral infections: Dengue, Chikungunya and Zika viruses. Viruses 2019, 11, 225. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, F.; Carinhas, N.; Carrondo, M.J.T.; Bernal, V.; Aves, P.M. Toward system-level understanding of baculovirus–host cell interactions: from molecular fundamental studies to large-scale proteomics approaches. Frontiers Microbiol. 2012, 3, 391. [Google Scholar] [CrossRef] [PubMed]
- Bernal, V.; Carinhas, N.; Yokomizo, A.Y.; Carrondo, M.J.T.; Alves, P.M. Cell density effect in the baculovirus–insect cells system: a quantitative analysis of energetic metabolism. Biotechnol. Bioeng. 2009, 104, 162–180. [Google Scholar] [CrossRef]
- Chambers, J.W.; Maguire, T.G.; Alwine, J.C. Glutamine metabolism is essential for human cytomegalovirus infection. J. Virol. 2010, 84, 1867–1873. [Google Scholar] [CrossRef]
- Eagle, H.; Habel, K. The nutritional requirements for the propagation of poliomyelitis virus by the HeLa cell. J. Exp. Med. 1956, 104, 271–287. [Google Scholar] [CrossRef]
- Lewis, V., Jr.; Scott, L.V. Nutritional requirements for the production of herpes simplex virus. I. Influence of glucose and glutamine of herpes simplex virus production by HeLa cells. J. Bacteriol. 1962, 83, 475–482. [Google Scholar] [CrossRef]
- Fontaine, K.A.; Camarda, R.; Lagunoff, M. Vaccinia Virus Requires Glutamine but Not Glucose for Efficient Replication. J. Virol. 2014, 88, 4366–4374. [Google Scholar] [CrossRef]
- Mounce, B.C.; Poirier, E.Z.; Passoni, G.; Simon-Loriere, E.; Cesaro, T.; Prot, M.; Stapleford, K.A.; Moratorio, G.; Sakuntabhai, A.; Levraud, J.-P.; et al. Interferon-Induced Spermidine-Spermine Acetyltransferase and Polyamine Depletion Restrict Zika and Chikungunya Viruses. Cell Host Microbe 2016, 20, 167–177. [Google Scholar] [CrossRef]
- Chotiwan, N.; Andre, B.G.; Sanchez-Vargas, I.; Islam, M.N.; Grabowski, J.M.; Hopf-Jannasch, A.; Gough, E.; Nakayasu, E.; Blair, C.D.; Belisle, J.T.; et al. Dynamic remodeling of lipids coincides with dengue virus replication in the midgut of Aedes aegypti mosquitoes. PLoS Pathog. 2018, 14, e1006853. [Google Scholar] [CrossRef]
- Lupberger, J.; Croonenborghs, T.; Suarez, A.A.R.; Van Renne, N.; Jühling, F.; Oudot, M.A.; Virzì, A.; Bandiera, S.; Jamey, C.; Meszaros, G.; et al. Combined analysis of metabolomes, proteomes and transcriptomes of Hepatitis C-infected cells and liver to identify pathways associated with disease development. Gastroenterology 2019, 157, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems biology and multi-omics integration: viewpoints from the metabolomics research community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | Total Number of Instances | Number of Model Components (to + tp) a | Model Characteristics b | Cross-Validated ANOVA c (p-Value) | Permutation d |
|---|---|---|---|---|---|
| 6 HPI vs. 0 HPI | 10 | 1+0 | R2Y = 0.998; Q2 = 0.995 | 1.25 × 10−8 | Good |
| 12 HPI vs. 0 HPI | 10 | 1+0 | R2Y = 1.000; Q2 = 0.998 | 1.98 × 10−10 | Good |
| 24 HPI vs. 0 HPI | 10 | 1+0 | R2Y = 0.998; Q2 = 0.993 | 2.34 × 10−8 | Good |
| 36 HPI vs. 0 HPI | 10 | 1+0 | R2Y = 0.997; Q2 = 0.992 | 5.29 × 10−8 | Good |
| 12 HPI vs. 6 HPI | 10 | 1+1 | R2Y = 0.985; Q2 = 0.940 | 3.23 × 10−3 | Good |
| 24 HPI vs. 6 HPI | 10 | 1+1 | R2Y = 0.997; Q2 = 0.990 | 3.42 × 10−5 | Good |
| 36 HPI vs. 6 HPI | 10 | 1+1 | R2Y = 0.998; Q2 = 0.997 | 1.94 × 10−6 | Good |
| 24 HPI vs. 12 HPI | 10 | 1+0 | R2Y = 0.998; Q2 = 0.993 | 2.46 × 10−8 | Good |
| 36 HPI vs. 12 HPI | 10 | 1+0 | R2Y = 0.993; Q 2= 0.99 | 8.96 × 10−8 | Good |
| 36 HPI vs. 24 HPI | 10 | 1+1 | R2Y = 0.995; Q2 = 0.986 | 8.63 × 10−5 | Good |
| Comparison | Pathway | Hits | Holm Adjusted P | FDR | Impact |
|---|---|---|---|---|---|
| 6 HPI vs. 0 HPI | None | ||||
| 12 HPI vs. 0 HPI | Galactose metabolism | 5 | 5.37 × 10−3 | 4.28 × 10−3 | 0.18 |
| Aminoacyl-tRNA biosynthesis | 7 | 8.45 × 10−3 | 4.28 × 10−3 | 0.00 | |
| Starch and sucrose metabolism | 4 | 1.39 × 10−2 | 4.77 × 10−3 | 0.21 | |
| 24 HPI vs. 0 HPI | None | ||||
| 36 HPI vs. 0 HPI | Arginine and proline metabolism | 7 | 8.43 × 10−4 | 8.43 × 10−4 | 0.33 |
| Aminoacyl-tRNA biosynthesis | 8 | 5.93 × 10−3 | 3.00 × 10−3 | 0.14 | |
| Alanine, aspartate and glutamate metabolism | 4 | 1.23 × 10−1 | 4.19 × 10−2 | 0.61 | |
| 12 HPI vs. 6 HPI | None | ||||
| 24 HPI vs. 12 HPI | Aminoacyl-tRNA biosynthesis | 9 | 1.05 × 10−4 | 1.05 × 10−4 | 0.14 |
| Galactose metabolism | 5 | 6.95 × 10−3 | 3.52 × 10−3 | 0.18 | |
| Starch and sucrose metabolism | 4 | 1.72 × 10−2 | 5.90 × 10−3 | 0.21 | |
| Arginine and proline metabolism | 5 | 3.90 × 10−2 | 1.01 × 10−2 | 0.29 | |
| Alanine, aspartate and glutamate metabolism | 4 | 5.78 × 10−2 | 1.22 × 10−3 | 0.61 | |
| 36 HPI vs. 24 HPI | Aminoacyl-tRNA biosynthesis | 8 | 1.95 × 10−3 | 1.95 × 10−3 | 0.14 |
| Arginine and proline metabolism | 5 | 5.12 × 10−2 | 2.59 × 10−2 | 0.33 |
| Amino acids | Carbohydrates | All Metabolites | ||
|---|---|---|---|---|
| Bm5 cells | 2 DPI (N = 34) | 19↑ 4↓ | 5↑ | 28↑ 6↓ |
| 1 WPI (N = 45) | 25↑ 1↓ | 9↑ | 41↑ 4↓ | |
| 2 WPI (N = 42) | 21↑ 5↓ | 6↑ 1↓ | 32↑ 10↓ | |
| 3 WPI (N = 43) | 20↑ 6↓ | 7↑ 1↓ | 32↑ 11↓ | |
| S2 cells | 6 HPI (N = 13) | 1↑ 4↓ | 3↑ 2↓ | 5↑ 8↓ |
| 12 HPI (N = 21) | 1↑ 9↓ | 3↑ 5↓ | 4↑ 17↓ | |
| 24 HPI (N = 6) | 4↑ 1↓ | 4↑ 1↓ | ||
| 36 HPI (N = 24) | 10↑ 4↓ | 2↑ 3↓ | 14↑ 10↓ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.-L.; Swevers, L.; Van Meulebroek, L.; Meeus, I.; Vanhaecke, L.; Smagghe, G. Metabolomic Analysis of Cricket paralysis virus Infection in Drosophila S2 Cells Reveals Divergent Effects on Central Carbon Metabolism as Compared with Silkworm Bm5 Cells. Viruses 2020, 12, 393. https://doi.org/10.3390/v12040393
Wang L-L, Swevers L, Van Meulebroek L, Meeus I, Vanhaecke L, Smagghe G. Metabolomic Analysis of Cricket paralysis virus Infection in Drosophila S2 Cells Reveals Divergent Effects on Central Carbon Metabolism as Compared with Silkworm Bm5 Cells. Viruses. 2020; 12(4):393. https://doi.org/10.3390/v12040393
Chicago/Turabian StyleWang, Luo-Luo, Luc Swevers, Lieven Van Meulebroek, Ivan Meeus, Lynn Vanhaecke, and Guy Smagghe. 2020. "Metabolomic Analysis of Cricket paralysis virus Infection in Drosophila S2 Cells Reveals Divergent Effects on Central Carbon Metabolism as Compared with Silkworm Bm5 Cells" Viruses 12, no. 4: 393. https://doi.org/10.3390/v12040393
APA StyleWang, L.-L., Swevers, L., Van Meulebroek, L., Meeus, I., Vanhaecke, L., & Smagghe, G. (2020). Metabolomic Analysis of Cricket paralysis virus Infection in Drosophila S2 Cells Reveals Divergent Effects on Central Carbon Metabolism as Compared with Silkworm Bm5 Cells. Viruses, 12(4), 393. https://doi.org/10.3390/v12040393