
Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Subjects
2.2. Ethics Statement
2.3. Dengue Diagnosis
2.4. Measurement of Serum Cytokine Levels
2.5. Flow Cytometry for CD4+IL-17 (Interleukin 17)-Producing Cells
2.6. Statistical Analysis
3. Results
3.1. Patient Characteristics
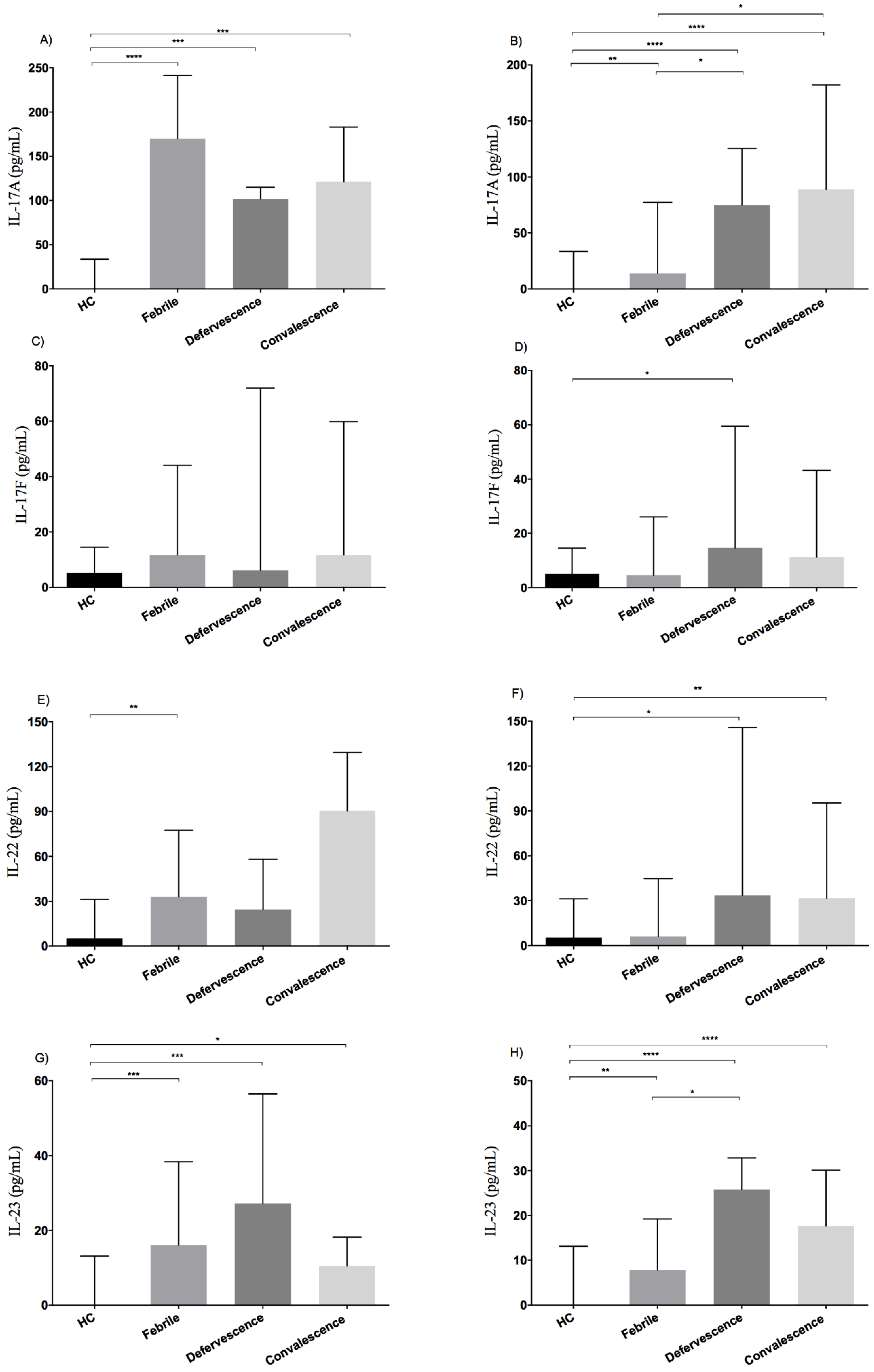
3.2. Elevated Serum Levels of IL-17A, IL-17F, IL-22 and IL-23 in Dengue Patients
3.3. Dynamics of Production of IL-17-Related Cytokines at Different Phases of Illness
3.4. Serum Levels of IL-17A and IL-22 Are Associated with Dengue Infection Severity
3.5. High Serum Levels of IL-17A and IL-23 Are Correlated during Dengue Infections
3.6. CD4+ IL-17-Producing Cells in DENV-Infected Patients Are Associated with Disease Severity
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.G.; Kouri, G. Dengue and dengue hemorrhagic fever in the Americas: Lessons and challenges. J. Clin. Virol. 2003, 27, 1–13. [Google Scholar] [CrossRef]
- Halstead, S.B. Pathogenesis of dengue: Challenges to molecular biology. Science 1988, 239, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Endy, T.P.; Nisalak, A.; Chunsuttitwat, S.; Vaughn, D.W.; Green, S.; Ennis, F.A.; Rothman, A.L.; Libraty, D.H. Relationship of preexisting dengue virus (DV) neutralizing antibody levels to viremia and severity of disease in a prospective cohort study of DV infection in Thailand. J. Infect. Dis. 2004, 189, 990–1000. [Google Scholar] [CrossRef] [Green Version]
- Guilarde, A.O.; Turchi, M.D.; Siqueira, J.B., Jr.; Feres, V.C.; Rocha, B.; Levi, J.E.; Souza, V.A.; Boas, L.S.; Pannuti, C.S.; Martelli, C.M. Dengue and dengue hemorrhagic fever among adults: Clinical outcomes related to viremia, serotypes, and antibody response. J. Infect. Dis. 2008, 197, 817–824. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, D.W.; Green, S.; Kalayanarooj, S.; Innis, B.L.; Nimmannitya, S.; Suntayakorn, S.; Endy, T.P.; Raengsakulrach, B.; Rothman, A.L.; Ennis, F.A.; et al. Dengue viremia titer, antibody response pattern, and virus serotype correlate with disease severity. J. Infect. Dis. 2000, 181, 2–9. [Google Scholar] [CrossRef]
- Chen, L.C.; Lei, H.Y.; Liu, C.C.; Shiesh, S.C.; Chen, S.H.; Liu, H.S.; Lin, Y.S.; Wang, S.T.; Shyu, H.W.; Yeh, T.M. Correlation of serum levels of macrophage migration inhibitory factor with disease severity and clinical outcome in dengue patients. Am. J. Trop. Med. Hyg. 2006, 74, 142–147. [Google Scholar] [CrossRef]
- Green, S.; Vaughn, D.W.; Kalayanarooj, S.; Nimmannitya, S.; Suntayakorn, S.; Nisalak, A.; Rothman, A.L.; Ennis, F.A. Elevated plasma interleukin-10 levels in acute dengue correlate with disease severity. J. Med Virol. 1999, 59, 329–334. [Google Scholar] [CrossRef]
- Pinto, L.M.; Oliveira, S.A.; Braga, E.L.; Nogueira, R.M.; Kubelka, C.F. Increased pro-inflammatory cytokines (TNF-alpha and IL-6) and anti-inflammatory compounds (sTNFRp55 and sTNFRp75) in Brazilian patients during exanthematic dengue fever. Mem. Inst. Oswaldo Cruz 1999, 94, 387–394. [Google Scholar] [CrossRef]
- Bozza, F.A.; Cruz, O.G.; Zagne, S.M.; Azeredo, E.L.; Nogueira, R.M.; Assis, E.F.; Bozza, P.T.; Kubelka, C.F. Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC Infect. Dis. 2008, 8, 86. [Google Scholar] [CrossRef] [Green Version]
- Mangada, M.M.; Rothman, A.L. Altered cytokine responses of dengue-specific CD4+ T cells to heterologous serotypes. J. Immunol. 2005, 175, 2676–2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatch, S.; Endy, T.P.; Thomas, S.; Mathew, A.; Potts, J.; Pazoles, P.; Libraty, D.H.; Gibbons, R.; Rothman, A.L. Intracellular cytokine production by dengue virus-specific T cells correlates with subclinical secondary infection. J. Infect. Dis. 2011, 203, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Rathakrishnan, A.; Wang, S.M.; Hu, Y.; Khan, A.M.; Ponnampalavanar, S.; Lum, L.C.; Manikam, R.; Sekaran, S.D. Cytokine expression profile of dengue patients at different phases of illness. PLoS ONE 2012, 7, e52215. [Google Scholar] [CrossRef] [PubMed]
- Mabalirajan, U.; Kadhiravan, T.; Sharma, S.K.; Banga, A.; Ghosh, B. Th(2) immune response in patients with dengue during defervescence: Preliminary evidence. Am. J. Trop. Med. Hyg. 2005, 72, 783–785. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Gao, W.; Awasthi, A.; Jager, A.; Strom, T.B.; Oukka, M.; Kuchroo, V.K. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature 2007, 448, 484–487. [Google Scholar] [CrossRef]
- Manel, N.; Unutmaz, D.; Littman, D.R. The differentiation of human T(H)-17 cells requires transforming growth factor-beta and induction of the nuclear receptor RORgammat. Nat. Immunol. 2008, 9, 641–649. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Anderson, D.E.; Baecher-Allan, C.; Hastings, W.D.; Bettelli, E.; Oukka, M.; Kuchroo, V.K.; Hafler, D.A. IL-21 and TGF-beta are required for differentiation of human T(H)17 cells. Nature 2008, 454, 350–352. [Google Scholar] [CrossRef]
- Aggarwal, S.; Ghilardi, N.; Xie, M.H.; de Sauvage, F.J.; Gurney, A.L. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 2003, 278, 1910–1914. [Google Scholar] [CrossRef] [Green Version]
- Wilke, C.M.; Bishop, K.; Fox, D.; Zou, W. Deciphering the role of Th17 cells in human disease. Trends Immunol. 2011, 32, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Van de Veerdonk, F.L.; Gresnigt, M.S.; Kullberg, B.J.; van der Meer, J.W.; Joosten, L.A.; Netea, M.G. Th17 responses and host defense against microorganisms: An overview. BMB Rep. 2009, 42, 776–787. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Li, J.; Zheng, Y.; Luo, Y.; Zhou, H.; Yao, Y.; Chen, X.; Chen, Z.; He, M. A randomized case-control study of dynamic changes in peripheral blood Th17/Treg cell balance and interleukin-17 levels in highly active antiretroviral-treated HIV type 1/AIDS patients. AIDS Res. Hum. Retrovir. 2012, 28, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, A.; Prado, J.G.; Kang, Y.H.; Chen, F.; Riddell, L.A.; Luzzi, G.; Goulder, P.; Klenerman, P. HIV-1 infection is characterized by profound depletion of CD161+ Th17 cells and gradual decline in regulatory T cells. Aids 2010, 24, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, Y.; Tuen, M.; Nadas, A.; Hioe, C.E. In vitro restoration of Th17 response during HIV infection with an antiretroviral drug and Th17 differentiation cytokines. AIDS Res. Hum. Retrovir. 2012, 28, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Rowan, A.G.; Fletcher, J.M.; Ryan, E.J.; Moran, B.; Hegarty, J.E.; O’Farrelly, C.; Mills, K.H. Hepatitis C virus-specific Th17 cells are suppressed by virus-induced TGF-beta. J. Immunol. 2008, 181, 4485–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Q.; Wang, Y.K.; Zhao, Q.; Wang, C.Z.; Hu, Y.Z.; Wu, B.Y. Th17 cells are increased with severity of liver inflammation in patients with chronic hepatitis C. J. Gastroenterol. Hepatol. 2012, 27, 273–278. [Google Scholar] [CrossRef]
- Fathy, A.; Ahmed, A.S.; Metwally, L.; Hassan, A. T helper type 1/T helper type 17-related cytokines in chronic hepatitis C patients before and after interferon and ribavirin therapy. Med. Princ. Pract. 2011, 20, 345–349. [Google Scholar] [CrossRef]
- Ge, J.; Wang, K.; Meng, Q.H.; Qi, Z.X.; Meng, F.L.; Fan, Y.C. Implication of Th17 and Th1 cells in patients with chronic active hepatitis B. J. Clin. Immunol. 2010, 30, 60–67. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhang, Z.; Lin, F.; Zou, Z.S.; Xu, R.N.; Jin, L.; Fu, J.L.; Shi, F.; Shi, M.; Wang, H.F.; et al. Interleukin-17-producing CD4(+) T cells increase with severity of liver damage in patients with chronic hepatitis B. Hepatology 2010, 51, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Panorama Epidemiológico Dengue 2014. Available online: http://www.epidemiologia.salud.gob.mx/doctos/panodengue/PANORAMAS_2014/Pano_dengue_sem_53_2014.pdf) (accessed on 10 December 2019).
- WHO. Dengue Hemorrhagic Fever: Diagnosis, Treatment, Prevention and Control; WHO: Geneva, Switzerland, 1997. [Google Scholar]
- Vaughn, D.W.; Nisalak, A.; Solomon, T.; Kalayanarooj, S.; Nguyen, M.D.; Kneen, R.; Cuzzubbo, A.; Devine, P.L. Rapid serologic diagnosis of dengue virus infection using a commercial capture ELISA that distinguishes primary and secondary infections. Am. J. Trop. Med. Hyg. 1999, 60, 693–698. [Google Scholar] [CrossRef]
- Picker, L.J.; Singh, M.K.; Zdraveski, Z.; Treer, J.R.; Waldrop, S.L.; Bergstresser, P.R.; Maino, V.C. Direct demonstration of cytokine synthesis heterogeneity among human memory/effector T cells by flow cytometry. Blood 1995, 86, 1408–1419. [Google Scholar] [CrossRef]
- Chatila, T.; Silverman, L.; Miller, R.; Geha, R. Mechanisms of T cell activation by the calcium ionophore ionomycin. J. Immunol. 1989, 143, 1283–1289. [Google Scholar] [PubMed]
- Karlsson, F.; Hassan-Zahraee, M. Quantification of Th1 and Th17 Cells with Intracellular Staining Following PMA/Ionomycin Stimulation. Curr. Protoc. Cytom. 2015, 71, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Brunialti, M.K.; Santos, M.C.; Rigato, O.; Machado, F.R.; Silva, E.; Salomao, R. Increased percentages of T helper cells producing IL-17 and monocytes expressing markers of alternative activation in patients with sepsis. PLoS ONE 2012, 7, e37393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Scaleia, R.; Barba, M.; Di Nardo, G.; Bonamico, M.; Oliva, S.; Nenna, R.; Valitutti, F.; Mennini, M.; Barbato, M.; Montuori, M.; et al. Size and dynamics of mucosal and peripheral IL-17A+ T-cell pools in pediatric age, and their disturbance in celiac disease. Mucosal Immunol. 2012, 5, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivanco-Cid, H.; Maldonado-Renteria, M.J.; Sanchez-Vargas, L.A.; Izaguirre-Hernandez, I.Y.; Hernandez-Flores, K.G.; Remes-Ruiz, R. Dynamics of interleukin-21 production during the clinical course of primary and secondary dengue virus infections. Immunol. Lett. 2014, 161, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Duran, A.; Valero, N.; Mosquera, J.; Pons, H.; Torres, M.; Alcocer, S.; Castillo, J.L. Dengue nonstructural protein-1 status is not associated to circulating levels of interleukin-17, C-reactive protein and complement in children with acute dengue. J. Clin. Virol. 2013, 56, 199–206. [Google Scholar] [CrossRef]
- Jain, A.; Pandey, N.; Garg, R.K.; Kumar, R. IL-17 level in patients with Dengue virus infection & its association with severity of illness. J. Clin. Immunol. 2013, 33, 613–618. [Google Scholar] [CrossRef]
- Furuta, T.; Murao, L.A.; Lan, N.T.; Huy, N.T.; Huong, V.T.; Thuy, T.T.; Tham, V.D.; Nga, C.T.; Ha, T.T.; Ohmoto, Y.; et al. Association of mast cell-derived VEGF and proteases in Dengue shock syndrome. PLoS Negl. Trop. Dis. 2012, 6, e1505. [Google Scholar] [CrossRef] [Green Version]
- Guabiraba, R.; Besnard, A.G.; Marques, R.E.; Maillet, I.; Fagundes, C.T.; Conceicao, T.M.; Rust, N.M.; Charreau, S.; Paris, I.; Lecron, J.C.; et al. IL-22 modulates IL-17A production and controls inflammation and tissue damage in experimental dengue infection. Eur. J. Immunol. 2013, 43, 1529–1544. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Adachi, M.; Oda, N.; Kokubu, F.; Huang, S.K. IL-17 cytokine family. J. Allergy Clin. Immunol. 2004, 114, 1265–1273. [Google Scholar] [CrossRef]
- Kolls, J.K.; Linden, A. Interleukin-17 family members and inflammation. Immunity 2004, 21, 467–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, C.T.; Hatton, R.D.; Mangan, P.R.; Harrington, L.E. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu. Rev. Immunol. 2007, 25, 821–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishigame, H.; Kakuta, S.; Nagai, T.; Kadoki, M.; Nambu, A.; Komiyama, Y.; Fujikado, N.; Tanahashi, Y.; Akitsu, A.; Kotaki, H.; et al. Differential roles of interleukin-17A and -17F in host defense against mucoepithelial bacterial infection and allergic responses. Immunity 2009, 30, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappu, R.; Ramirez-Carrozzi, V.; Sambandam, A. The interleukin-17 cytokine family: Critical players in host defence and inflammatory diseases. Immunology 2011, 134, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, L.; Vergis, A.L.; Ye, H.; Bajwa, A.; Narayan, V.; Strieter, R.M.; Rosin, D.L.; Okusa, M.D. IL-17 produced by neutrophils regulates IFN-gamma-mediated neutrophil migration in mouse kidney ischemia-reperfusion injury. J. Clin. Investig. 2010, 120, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockhart, E.; Green, A.M.; Flynn, J.L. IL-17 production is dominated by gammadelta T cells rather than CD4 T cells during Mycobacterium tuberculosis infection. J. Immunol. 2006, 177, 4662–4669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, M.L.; Keller, A.C.; Paget, C.; Fujio, M.; Trottein, F.; Savage, P.B.; Wong, C.H.; Schneider, E.; Dy, M.; Leite-de-Moraes, M.C. Identification of an IL-17-producing NK1.1(neg) iNKT cell population involved in airway neutrophilia. J. Exp. Med. 2007, 204, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Vanlaere, I.; de Rycke, R.; Cauwels, A.; Joosten, L.A.; Lubberts, E.; van den Berg, W.B.; Libert, C. IL-17 produced by Paneth cells drives TNF-induced shock. J. Exp. Med. 2008, 205, 1755–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, K.; Gozzi, L.; Iannone, A.; Lo Tartaro, D.; Mattioli, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Jovanovic, D.V.; Di Battista, J.A.; Martel-Pelletier, J.; Jolicoeur, F.C.; He, Y.; Zhang, M.; Mineau, F.; Pelletier, J.P. IL-17 stimulates the production and expression of proinflammatory cytokines, IL-beta and TNF-alpha, by human macrophages. J. Immunol. 1998, 160, 3513–3521. [Google Scholar]
- Erbel, C.; Akhavanpoor, M.; Okuyucu, D.; Wangler, S.; Dietz, A.; Zhao, L.; Stellos, K.; Little, K.M.; Lasitschka, F.; Doesch, A.; et al. IL-17A influences essential functions of the monocyte/macrophage lineage and is involved in advanced murine and human atherosclerosis. J. Immunol. 2014, 193, 4344–4355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.Q.; Suarez-Farinas, M.; Nograles, K.E.; Mimoso, C.A.; Shrom, D.; Dow, E.R.; Heffernan, M.P.; Hoffman, R.W.; Krueger, J.G. IL-17 induces inflammation-associated gene products in blood monocytes, and treatment with ixekizumab reduces their expression in psoriasis patient blood. J. Investig. Dermatol. 2014, 134, 2990–2993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, S.; Gurney, A.L. IL-17: Prototype member of an emerging cytokine family. J. Leukoc. Biol. 2002, 71, 1–8. [Google Scholar] [PubMed]
- Sebkova, L.; Pellicano, A.; Monteleone, G.; Grazioli, B.; Guarnieri, G.; Imeneo, M.; Pallone, F.; Luzza, F. Extracellular signal-regulated protein kinase mediates interleukin 17 (IL-17)-induced IL-8 secretion in Helicobacter pylori-infected human gastric epithelial cells. Infect. Immun. 2004, 72, 5019–5026. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, W.A.; Vanaudenaerde, B.M.; Dupont, L.J.; Van Raemdonck, D.E.; Demedts, M.G.; Verleden, G.M. Interleukin-17--induced interleukin-8 release in human airway smooth muscle cells: Role for mitogen-activated kinases and nuclear factor-kappaB. J. Heart Lung Transplant. 2005, 24, 875–881. [Google Scholar] [CrossRef]
- Hwang, S.Y.; Kim, J.Y.; Kim, K.W.; Park, M.K.; Moon, Y.; Kim, W.U.; Kim, H.Y. IL-17 induces production of IL-6 and IL-8 in rheumatoid arthritis synovial fibroblasts via NF-kappaB- and PI3-kinase/Akt-dependent pathways. Arthritis Res. Ther. 2004, 6, R120–R128. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Hu, X.; Liu, C.; Qv, X.; Xu, C. Interleukin (IL)-17 promotes macrophages to produce IL-8, IL-6 and tumour necrosis factor-alpha in aplastic anaemia. Br. J. Haematol. 2008, 142, 109–114. [Google Scholar] [CrossRef]
- Chen, J.; Liao, M.Y.; Gao, X.L.; Zhong, Q.; Tang, T.T.; Yu, X.; Liao, Y.H.; Cheng, X. IL-17A induces pro-inflammatory cytokines production in macrophages via MAPKinases, NF-kappaB and AP-1. Cell. Physiol. Biochem. 2013, 32, 1265–1274. [Google Scholar] [CrossRef]
- Juffrie, M.; van Der Meer, G.M.; Hack, C.E.; Haasnoot, K.; Sutaryo; Veerman, A.J.; Thijs, L.G. Inflammatory mediators in dengue virus infection in children: Interleukin-8 and its relationship to neutrophil degranulation. Infect. Immun. 2000, 68, 702–707. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; Yang, J.; Yang, X.; Wei, Y.; Zhu, L.; Gao, D.; Li, M. IL-17A induces endothelial inflammation in systemic sclerosis via the ERK signaling pathway. PLoS ONE 2013, 8, e85032. [Google Scholar] [CrossRef]
- Djamiatun, K.; van der Ven, A.J.; de Groot, P.G.; Faradz, S.M.; Hapsari, D.; Dolmans, W.M.; Sebastian, S.; Fijnheer, R.; de Mast, Q. Severe dengue is associated with consumption of von Willebrand factor and its cleaving enzyme ADAMTS-13. PLoS Negl. Trop. Dis. 2012, 6, e1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Wang, Q.; Guo, C.; Wang, X.; Cao, X.; Shi, Y.; Gao, F.; Ma, C.; Zhang, L. IL-17 induces apoptosis of vascular endothelial cells: A potential mechanism for human acute coronary syndrome. Clin. Immunol. 2011, 141, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Vasquez Ochoa, M.; Garcia Cordero, J.; Gutierrez Castaneda, B.; Santos Argumedo, L.; Villegas Sepulveda, N.; Cedillo Barron, L. A clinical isolate of dengue virus and its proteins induce apoptosis in HMEC-1 cells: A possible implication in pathogenesis. Arch. Virol. 2009, 154, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Xu, J.; Huang, J. FasL/Fas pathway is involved in dengue virus induced apoptosis of the vascular endothelial cells. J. Med. Virol. 2010, 82, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Tanaka, H.; Kawato, T.; Kitami, S.; Nakai, K.; Motohashi, M.; Suzuki, N.; Wang, C.L.; Ochiai, K.; Isokawa, K.; et al. Interleukin-17A induces cathepsin K and MMP-9 expression in osteoclasts via celecoxib-blocked prostaglandin E2 in osteoblasts. Biochimie 2011, 93, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Starodubtseva, N.L.; Sobolev, V.V.; Soboleva, A.G.; Nikolaev, A.A.; Bruskin, S.A. [Expression of genes for metalloproteinases (MMP-1, MMP-2, MMP-9, and MMP-12) associated with psoriasis]. Genetika 2011, 47, 1254–1261. [Google Scholar] [CrossRef]
- Kubelka, C.F.; Azeredo, E.L.; Gandini, M.; Oliveira-Pinto, L.M.; Barbosa, L.S.; Damasco, P.V.; Avila, C.A.; Motta-Castro, A.R.; Cunha, R.V.; Cruz, O.G. Metalloproteinases are produced during dengue fever and MMP9 is associated with severity. J. Infect. 2010, 61, 501–505. [Google Scholar] [CrossRef]
- Marder, W.; Khalatbari, S.; Myles, J.D.; Hench, R.; Yalavarthi, S.; Lustig, S.; Brook, R.; Kaplan, M.J. Interleukin 17 as a novel predictor of vascular function in rheumatoid arthritis. Ann. Rheum. Dis. 2011, 70, 1550–1555. [Google Scholar] [CrossRef] [Green Version]
- Golden, J.B.; McCormick, T.S.; Ward, N.L. IL-17 in psoriasis: Implications for therapy and cardiovascular co-morbidities. Cytokine 2013, 62, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Kang, H.S.; Kim, B.S. Th17 cells enhance viral persistence and inhibit T cell cytotoxicity in a model of chronic virus infection. J. Exp. Med. 2009, 206, 313–328. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, Y.; Tomimori, Y.; Yumoto, K.; Hasegawa, S.; Ando, T.; Tagaya, Y.; Crotty, S.; Kawakami, T. Inhibition of NK cell activity by IL-17 allows vaccinia virus to induce severe skin lesions in a mouse model of eczema vaccinatum. J. Exp. Med. 2009, 206, 1219–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, S.C.; Nickerson-Nutter, C.; Pittman, D.D.; Carrier, Y.; Goodwin, D.G.; Shields, K.M.; Lambert, A.J.; Schelling, S.H.; Medley, Q.G.; Ma, H.L.; et al. IL-22 induces an acute-phase response. J. Immunol. 2010, 185, 5531–5538. [Google Scholar] [CrossRef] [Green Version]
- Radaeva, S.; Sun, R.; Pan, H.N.; Hong, F.; Gao, B. Interleukin 22 (IL-22) plays a protective role in T cell-mediated murine hepatitis: IL-22 is a survival factor for hepatocytes via STAT3 activation. Hepatology 2004, 39, 1332–1342. [Google Scholar] [CrossRef]
- Zenewicz, L.A.; Yancopoulos, G.D.; Valenzuela, D.M.; Murphy, A.J.; Karow, M.; Flavell, R.A. Interleukin-22 but not interleukin-17 provides protection to hepatocytes during acute liver inflammation. Immunity 2007, 27, 647–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, S.; Dambacher, J.; Beigel, F.; Zitzmann, K.; Heeg, M.H.; Weiss, T.S.; Prufer, T.; Olszak, T.; Steib, C.J.; Storr, M.; et al. IL-22-mediated liver cell regeneration is abrogated by SOCS-1/3 overexpression in vitro. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1019–G1028. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Hu, B.; Colletti, L.M. IL-22 is involved in liver regeneration after hepatectomy. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G74–G80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaeffer, E.; Flacher, V.; Papageorgiou, V.; Decossas, M.; Fauny, J.D.; Kramer, M.; Mueller, C.G. Dermal CD14(+) Dendritic Cell and Macrophage Infection by Dengue Virus Is Stimulated by Interleukin-4. J. Investig. Dermatol. 2015, 135, 1743–1751. [Google Scholar] [CrossRef] [Green Version]
- Roses, R.E.; Xu, S.; Xu, M.; Koldovsky, U.; Koski, G.; Czerniecki, B.J. Differential production of IL-23 and IL-12 by myeloid-derived dendritic cells in response to TLR agonists. J. Immunol. 2008, 181, 5120–5127. [Google Scholar] [CrossRef] [Green Version]
- Andersson, A.; Kokkola, R.; Wefer, J.; Erlandsson-Harris, H.; Harris, R.A. Differential macrophage expression of IL-12 and IL-23 upon innate immune activation defines rat autoimmune susceptibility. J. Leukoc. Biol. 2004, 76, 1118–1124. [Google Scholar] [CrossRef]
- Yannam, G.R.; Gutti, T.; Poluektova, L.Y. IL-23 in infections, inflammation, autoimmunity and cancer: Possible role in HIV-1 and AIDS. J. Neuroimmune Pharmacol. 2012, 7, 95–112. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | DF n = 109 (71.7%) | DHF n = 43 (28.3%) | p Value |
|---|---|---|---|
| Male sex n= (%) | 38 (34.9) | 20 (46.5) | 0.1986 |
| Days of illness median (range) | 5 (1–28) | 7 (2–12) | <0.0001 |
| Age (years) median (range) | 26 (12–66) | 26 (12–63) | 0.8585 |
| Hematocrit % mean (SD) | 39.2 (3.9) | 41.3 (4.8) | 0.0147 |
| Platelet (103/mm3) median (range) | 151.5 (23–465) | 66 (16–246) | <0.0001 |
| Lymphocytes (103/mm3) median (range) | 0.9 (0.2–4.8) | 1.7 (0.4–6.1) | <0.0025 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Vargas, L.A.; Hernández-Flores, K.G.; Thomas-Dupont, P.; Izaguirre-Hernández, I.Y.; Sánchez-Marce, E.E.; Remes-Ruiz, R.; Fonseca-Coronado, S.; Hernández-Romano, P.A.; Flores-Collins, M.E.; Vivanco-Cid, H. Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections. Viruses 2020, 12, 1435. https://doi.org/10.3390/v12121435
Sánchez-Vargas LA, Hernández-Flores KG, Thomas-Dupont P, Izaguirre-Hernández IY, Sánchez-Marce EE, Remes-Ruiz R, Fonseca-Coronado S, Hernández-Romano PA, Flores-Collins ME, Vivanco-Cid H. Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections. Viruses. 2020; 12(12):1435. https://doi.org/10.3390/v12121435
Chicago/Turabian StyleSánchez-Vargas, Luis Alberto, Karina Guadalupe Hernández-Flores, Pablo Thomas-Dupont, Irma Yadira Izaguirre-Hernández, Elvis Efraín Sánchez-Marce, Ricardo Remes-Ruiz, Salvador Fonseca-Coronado, Pablo Augurio Hernández-Romano, María Estrella Flores-Collins, and Héctor Vivanco-Cid. 2020. "Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections" Viruses 12, no. 12: 1435. https://doi.org/10.3390/v12121435
APA StyleSánchez-Vargas, L. A., Hernández-Flores, K. G., Thomas-Dupont, P., Izaguirre-Hernández, I. Y., Sánchez-Marce, E. E., Remes-Ruiz, R., Fonseca-Coronado, S., Hernández-Romano, P. A., Flores-Collins, M. E., & Vivanco-Cid, H. (2020). Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections. Viruses, 12(12), 1435. https://doi.org/10.3390/v12121435