The Old and the New: Prospects for Non-Integrating Lentiviral Vector Technology


{kind=link}
Abstract
1. Lentiviral Vectors
2. Insertional Mutagenesis Safety Concerns and the Rise of Non-Integrating LVs (NILVs)
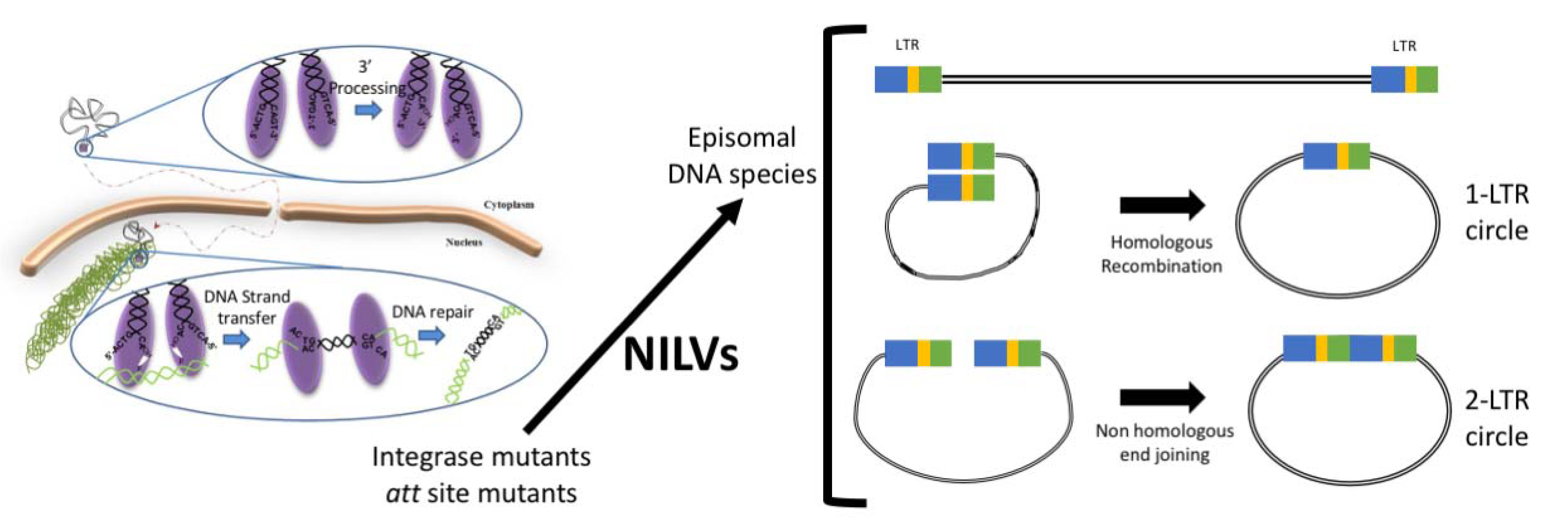
3. Non-Integrating Lentiviral Vector Development
4. Expression Levels of NILVs and Other Vector Designs
5. Old and Current Applications of NILVs
6. Prospects for NILV Technology
7. Conclusions
Funding
Conflicts of Interest
References
- Naldini, L.; Blomer, U.; Gallay, P.; Ory, D.; Mulligan, R.; Gage, F.H.; Verma, I.M.; Trono, D. In Vivo Gene Delivery and Stable Transduction of Nondividing Cells by a Lentiviral Vector. Science 1996, 272, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Zufferey, R.; Nagy, D.; Mandel, R.J.; Naldini, L.; Trono, D. Multiply Attenuated Lentiviral Vector Achieves Efficient Gene Delivery in Vivo. Nat. Biotechnol. 1997, 15, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A Third-Generation Lentivirus Vector with a Conditional Packaging System. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [CrossRef]
- Burns, J.C.; Friedmann, T.; Driever, W.; Burrascano, M.; Yee, J.K. Vesicular Stomatitis Virus G Glycoprotein Pseudotyped Retroviral Vectors: Concentration to very High Titer and Efficient Gene Transfer into Mammalian and Nonmammalian Cells. Proc. Natl. Acad. Sci. USA 1993, 90, 8033–8037. [Google Scholar] [CrossRef]
- Gutierrez-Guerrero, A.; Cosset, F.L.; Verhoeyen, E. Lentiviral Vector Pseudotypes: Precious Tools to Improve Gene Modification of Hematopoietic Cells for Research and Gene Therapy. Viruses 2020, 12, 1016. [Google Scholar] [CrossRef] [PubMed]
- Malim, M.H.; Hauber, J.; Le, S.Y.; Maizel, J.V.; Cullen, B.R. The HIV-1 Rev Trans-Activator Acts through a Structured Target Sequence to Activate Nuclear Export of Unspliced Viral mRNA. Nature 1989, 338, 254–257. [Google Scholar] [CrossRef]
- Zufferey, R.; Dull, T.; Mandel, R.J.; Bukovsky, A.; Quiroz, D.; Naldini, L.; Trono, D. Self-Inactivating Lentivirus Vector for Safe and Efficient in Vivo Gene Delivery. J. Virol. 1998, 72, 9873–9880. [Google Scholar] [CrossRef]
- Zennou, V.; Petit, C.; Guetard, D.; Nerhbass, U.; Montagnier, L.; Charneau, P. HIV-1 Genome Nuclear Import is Mediated by a Central DNA Flap. Cell 2000, 101, 173–185. [Google Scholar] [CrossRef]
- Follenzi, A.; Ailles, L.E.; Bakovic, S.; Geuna, M.; Naldini, L. Gene Transfer by Lentiviral Vectors is Limited by Nuclear Translocation and Rescued by HIV-1 Pol Sequences. Nat. Genet. 2000, 25, 217–222. [Google Scholar] [CrossRef]
- Zufferey, R.; Donello, J.E.; Trono, D.; Hope, T.J. Woodchuck Hepatitis Virus Posttranscriptional Regulatory Element Enhances Expression of Transgenes Delivered by Retroviral Vectors. J. Virol. 1999, 73, 2886–2892. [Google Scholar] [CrossRef]
- Gaspar, H.B.; Parsley, K.L.; Howe, S.; King, D.; Gilmour, K.C.; Sinclair, J.; Brouns, G.; Schmidt, M.; Von Kalle, C.; Barington, T.; et al. Gene Therapy of X-Linked Severe Combined Immunodeficiency by use of a Pseudotyped Gammaretroviral Vector. Lancet 2004, 364, 2181–2187. [Google Scholar] [CrossRef]
- Cavazzana-Calvo, M.; Hacein-Bey, S.; de Saint Basile, G.; Gross, F.; Yvon, E.; Nusbaum, P.; Selz, F.; Hue, C.; Certain, S.; Casanova, J.L.; et al. Gene Therapy of Human Severe Combined Immunodeficiency (SCID)-X1 Disease. Science 2000, 288, 669–672. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey-Abina, S.; Garrigue, A.; Wang, G.P.; Soulier, J.; Lim, A.; Morillon, E.; Clappier, E.; Caccavelli, L.; Delabesse, E.; Beldjord, K.; et al. Insertional Oncogenesis in 4 Patients After Retrovirus-Mediated Gene Therapy of SCID-X. J. Clin. Investig. 2008, 118, 3132–3142. [Google Scholar] [CrossRef] [PubMed]
- Howe, S.J.; Mansour, M.R.; Schwarzwaelder, K.; Bartholomae, C.; Hubank, M.; Kempski, H.; Brugman, M.H.; Pike-Overzet, K.; Chatters, S.J.; de Ridder, D.; et al. Insertional Mutagenesis Combined with Acquired Somatic Mutations Causes Leukemogenesis Following Gene Therapy of SCID-X1 Patients. J. Clin. Investig. 2008, 118, 3143–3150. [Google Scholar] [CrossRef] [PubMed]
- Hacein-Bey-Abina, S.; Von Kalle, C.; Schmidt, M.; McCormack, M.P.; Wulffraat, N.; Leboulch, P.; Lim, A.; Osborne, C.S.; Pawliuk, R.; Morillon, E.; et al. LMO2-Associated Clonal T Cell Proliferation in Two Patients After Gene Therapy for SCID-X1. Science 2003, 302, 415–419. [Google Scholar] [CrossRef]
- Braun, C.J.; Boztug, K.; Paruzynski, A.; Witzel, M.; Schwarzer, A.; Rothe, M.; Modlich, U.; Beier, R.; Gohring, G.; Steinemann, D.; et al. Gene Therapy for Wiskott-Aldrich Syndrome--Long-Term Efficacy and Genotoxicity. Sci. Transl. Med. 2014, 6, 227ra33. [Google Scholar] [CrossRef]
- Ott, M.G.; Schmidt, M.; Schwarzwaelder, K.; Stein, S.; Siler, U.; Koehl, U.; Glimm, H.; Kuhlcke, K.; Schilz, A.; Kunkel, H.; et al. Correction of X-Linked Chronic Granulomatous Disease by Gene Therapy, Augmented by Insertional Activation of MDS1-EVI1, PRDM16 Or SETBP1. Nat. Med. 2006, 12, 401–409. [Google Scholar] [CrossRef]
- Stein, S.; Ott, M.G.; Schultze-Strasser, S.; Jauch, A.; Burwinkel, B.; Kinner, A.; Schmidt, M.; Kramer, A.; Schwable, J.; Glimm, H.; et al. Genomic Instability and Myelodysplasia with Monosomy 7 Consequent to EVI1 Activation After Gene Therapy for Chronic Granulomatous Disease. Nat. Med. 2010, 16, 198–204. [Google Scholar] [CrossRef]
- Cavazzana-Calvo, M.; Payen, E.; Negre, O.; Wang, G.; Hehir, K.; Fusil, F.; Down, J.; Denaro, M.; Brady, T.; Westerman, K.; et al. Transfusion Independence and HMGA2 Activation After Gene Therapy of Human Beta-Thalassaemia. Nature 2010, 467, 318–322. [Google Scholar] [CrossRef]
- Modlich, U.; Bohne, J.; Schmidt, M.; von Kalle, C.; Knoss, S.; Schambach, A.; Baum, C. Cell-Culture Assays Reveal the Importance of Retroviral Vector Design for Insertional Genotoxicity. Blood 2006, 108, 2545–2553. [Google Scholar] [CrossRef]
- Bokhoven, M.; Stephen, S.L.; Knight, S.; Gevers, E.F.; Robinson, I.C.; Takeuchi, Y.; Collins, M.K. Insertional Gene Activation by Lentiviral and Gammaretroviral Vectors. J. Virol. 2009, 83, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.S.; McClure, M.A.; Feng, D.F.; Gray, J.; Doolittle, R.F. Computer Analysis of Retroviral Pol Genes: Assignment of Enzymatic Functions to Specific Sequences and Homologies with Nonviral Enzymes. Proc. Natl. Acad. Sci. USA 1986, 83, 7648–7652. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Jenkins, T.M.; Craigie, R. Zinc Folds the N-Terminal Domain of HIV-1 Integrase, Promotes Multimerization, and Enhances Catalytic Activity. Proc. Natl. Acad. Sci. USA 1996, 93, 13659–13664. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.; Craigie, R. Identification of Conserved Amino Acid Residues Critical for Human Immunodeficiency Virus Type 1 Integrase Function in Vitro. J. Virol. 1992, 66, 6361–6369. [Google Scholar] [CrossRef]
- Mumm, S.R.; Grandgenett, D.P. Defining Nucleic Acid-Binding Properties of Avian Retrovirus Integrase by Deletion Analysis. J. Virol. 1991, 65, 1160–1167. [Google Scholar] [CrossRef]
- Maertens, G.; Cherepanov, P.; Pluymers, W.; Busschots, K.; De Clercq, E.; Debyser, Z.; Engelborghs, Y. LEDGF/p75 is Essential for Nuclear and Chromosomal Targeting of HIV-1 Integrase in Human Cells. J. Biol. Chem. 2003, 278, 33528–33539. [Google Scholar] [CrossRef]
- Ciuffi, A.; Llano, M.; Poeschla, E.; Hoffmann, C.; Leipzig, J.; Shinn, P.; Ecker, J.R.; Bushman, F. A Role for LEDGF/p75 in Targeting HIV DNA Integration. Nat. Med. 2005, 11, 1287–1289. [Google Scholar] [CrossRef]
- Katzman, M.; Katz, R.A.; Skalka, A.M.; Leis, J. The Avian Retroviral Integration Protein Cleaves the Terminal Sequences of Linear Viral DNA at the in Vivo Sites of Integration. J. Virol. 1989, 63, 5319–5327. [Google Scholar] [CrossRef]
- Bushman, F.D.; Craigie, R. Activities of Human Immunodeficiency Virus (HIV) Integration Protein in Vitro: Specific Cleavage and Integration of HIV DNA. Proc. Natl. Acad. Sci. USA 1991, 88, 1339–1343. [Google Scholar] [CrossRef]
- Varmus, H.E.; Guntaka, R.V.; Fan, W.J.; Heasley, S.; Bishop, J.M. Synthesis of Viral DNA in the Cytoplasm of Duck Embryo Fibroblasts and in Enucleated Cells After Infection by Avian Sarcoma Virus. Proc. Natl. Acad. Sci. USA 1974, 71, 3874–3878. [Google Scholar] [CrossRef]
- Gianni, A.M.; Smotkin, D.; Weinberg, R.A. Murine Leukemia Virus: Detection of Unintegrated Double-Stranded DNA Forms of the Provirus. Proc. Natl. Acad. Sci. USA 1975, 72, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Ju, G.; Skalka, A.M. Nucleotide Sequence Analysis of the Long Terminal Repeat (LTR) of Avian Retroviruses: Structural Similarities with Transposable Elements. Cell 1980, 22, 379–386. [Google Scholar] [CrossRef]
- Shoemaker, C.; Goff, S.; Gilboa, E.; Paskind, M.; Mitra, S.W.; Baltimore, D. Structure of a Cloned Circular Moloney Murine Leukemia Virus DNA Molecule Containing an Inverted Segment: Implications for Retrovirus Integration. Proc. Natl. Acad. Sci. USA 1980, 77, 3932–3936. [Google Scholar] [CrossRef]
- Wu, Y.; Marsh, J.W. Early Transcription from Nonintegrated DNA in Human Immunodeficiency Virus Infection. J. Virol. 2003, 77, 10376–10382. [Google Scholar] [CrossRef] [PubMed]
- Engelman, A.; Englund, G.; Orenstein, J.M.; Martin, M.A.; Craigie, R. Multiple Effects of Mutations in Human Immunodeficiency Virus Type 1 Integrase on Viral Replication. J. Virol. 1995, 69, 2729–2736. [Google Scholar] [CrossRef] [PubMed]
- Poon, B.; Chen, I.S. Human Immunodeficiency Virus Type 1 (HIV-1) Vpr Enhances Expression from Unintegrated HIV-1 DNA. J. Virol. 2003, 77, 3962–3972. [Google Scholar] [CrossRef] [PubMed]
- Yanez-Munoz, R.J.; Balaggan, K.S.; MacNeil, A.; Howe, S.J.; Schmidt, M.; Smith, A.J.; Buch, P.; MacLaren, R.E.; Anderson, P.N.; Barker, S.E.; et al. Effective Gene Therapy with Nonintegrating Lentiviral Vectors. Nat. Med. 2006, 12, 348–353. [Google Scholar] [CrossRef]
- Leavitt, A.D.; Robles, G.; Alesandro, N.; Varmus, H.E. Human Immunodeficiency Virus Type 1 Integrase Mutants Retain in Vitro Integrase Activity Yet Fail to Integrate Viral DNA Efficiently during Infection. J. Virol. 1996, 70, 721–728. [Google Scholar] [CrossRef]
- Cornu, T.I.; Cathomen, T. Targeted Genome Modifications using Integrase-Deficient Lentiviral Vectors. Mol. Ther. 2007, 15, 2107–2113. [Google Scholar] [CrossRef]
- Apolonia, L.; Waddington, S.N.; Fernandes, C.; Ward, N.J.; Bouma, G.; Blundell, M.P.; Thrasher, A.J.; Collins, M.K.; Philpott, N.J. Stable Gene Transfer to Muscle using Non-Integrating Lentiviral Vectors. Mol. Ther. 2007, 15, 1947–1954. [Google Scholar] [CrossRef]
- Emiliani, S.; Mousnier, A.; Busschots, K.; Maroun, M.; Van Maele, B.; Tempe, D.; Vandekerckhove, L.; Moisant, F.; Ben-Slama, L.; Witvrouw, M.; et al. Integrase Mutants Defective for Interaction with LEDGF/p75 are Impaired in Chromosome Tethering and HIV-1 Replication. J. Biol. Chem. 2005, 280, 25517–25523. [Google Scholar] [CrossRef] [PubMed]
- Nightingale, S.J.; Hollis, R.P.; Pepper, K.A.; Petersen, D.; Yu, X.J.; Yang, C.; Bahner, I.; Kohn, D.B. Transient Gene Expression by Nonintegrating Lentiviral Vectors. Mol. Ther. 2006, 13, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.M.; Joseph, G.L.; Jasti, A.C.; Sastry-Dent, L.; Witting, S.; Cornetta, K. Differences in Vector-Genome Processing and Illegitimate Integration of Non-Integrating Lentiviral Vectors. Gene Ther. 2017, 24, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Kantor, B.; Bayer, M.; Ma, H.; Samulski, J.; Li, C.; McCown, T.; Kafri, T. Notable Reduction in Illegitimate Integration Mediated by a PPT-Deleted, Nonintegrating Lentiviral Vector. Mol. Ther. 2011, 19, 547–556. [Google Scholar] [CrossRef]
- Hu, P.; Li, Y.; Sands, M.S.; McCown, T.; Kafri, T. Generation of a Stable Packaging Cell Line Producing High-Titer PPT-Deleted Integration-Deficient Lentiviral Vectors. Mol. Ther. Methods Clin. Dev. 2015, 2, 15025. [Google Scholar] [CrossRef]
- Hu, P.; Bi, Y.; Ma, H.; Suwanmanee, T.; Zeithaml, B.; Fry, N.J.; Kohn, D.B.; Kafri, T. Superior Lentiviral Vectors Designed for BSL-0 Environment Abolish Vector Mobilization. Gene Ther. 2018, 25, 454–472. [Google Scholar] [CrossRef] [PubMed]
- Kantor, B.; Ma, H.; Webster-Cyriaque, J.; Monahan, P.E.; Kafri, T. Epigenetic Activation of Unintegrated HIV-1 Genomes by Gut-Associated Short Chain Fatty Acids and its Implications for HIV Infection. Proc. Natl. Acad. Sci. USA 2009, 106, 18786–18791. [Google Scholar] [CrossRef]
- Pelascini, L.P.; Janssen, J.M.; Goncalves, M.A. Histone Deacetylase Inhibition Activates Transgene Expression from Integration-Defective Lentiviral Vectors in Dividing and Non-Dividing Cells. Hum. Gene Ther. 2013, 24, 78–96. [Google Scholar] [CrossRef]
- Bayer, M.; Kantor, B.; Cockrell, A.; Ma, H.; Zeithaml, B.; Li, X.; McCown, T.; Kafri, T. A Large U3 Deletion Causes Increased in Vivo Expression from a Nonintegrating Lentiviral Vector. Mol. Ther. 2008, 16, 1968–1976. [Google Scholar] [CrossRef]
- Negri, D.R.; Rossi, A.; Blasi, M.; Michelini, Z.; Leone, P.; Chiantore, M.V.; Baroncelli, S.; Perretta, G.; Cimarelli, A.; Klotman, M.E.; et al. Simian Immunodeficiency Virus-Vpx for Improving Integrase Defective Lentiviral Vector-Based Vaccines. Retrovirology 2012, 9, 69. [Google Scholar] [CrossRef]
- Berger, G.; Goujon, C.; Darlix, J.L.; Cimarelli, A. SIVMAC Vpx Improves the Transduction of Dendritic Cells with Nonintegrative HIV-1-Derived Vectors. Gene Ther. 2009, 16, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Sather, B.D.; Wang, X.; Adair, J.; Khan, I.; Singh, S.; Lang, S.; Adams, A.; Curinga, G.; Kiem, H.P.; et al. Rapamycin Relieves Lentiviral Vector Transduction Resistance in Human and Mouse Hematopoietic Stem Cells. Blood 2014, 124, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Ozog, S.; Timberlake, N.D.; Hermann, K.; Garijo, O.; Haworth, K.G.; Shi, G.; Glinkerman, C.M.; Schefter, L.E.; D’Souza, S.; Simpson, E.; et al. Resveratrol Trimer Enhances Gene Delivery to Hematopoietic Stem Cells by Reducing Antiviral Restriction at Endosomes. Blood 2019, 134, 1298–1311. [Google Scholar] [CrossRef]
- Ngom, M.; Imren, S.; Maetzig, T.; Adair, J.E.; Knapp, D.J.H.F.; Chagraoui, J.; Fares, I.; Bordeleau, M.E.; Sauvageau, G.; Leboulch, P.; et al. UM171 Enhances Lentiviral Gene Transfer and Recovery of Primitive Human Hematopoietic Cells. Mol. Ther. Methods Clin. Dev. 2018, 10, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, C.; Thorne, L.G.; Unali, G.; Schiroli, G.; Giordano, A.M.S.; Piras, F.; Cuccovillo, I.; Petit, S.J.; Ahsan, F.; Noursadeghi, M.; et al. Cyclosporine H Overcomes Innate Immune Restrictions to Improve Lentiviral Transduction and Gene Editing in Human Hematopoietic Stem Cells. Cell. Stem Cell. 2018, 23, 820–832.e9. [Google Scholar] [CrossRef] [PubMed]
- Heffner, G.C.; Bonner, M.; Christiansen, L.; Pierciey, F.J.; Campbell, D.; Smurnyy, Y.; Zhang, W.; Hamel, A.; Shaw, S.; Lewis, G.; et al. Prostaglandin E2 Increases Lentiviral Vector Transduction Efficiency of Adult Human Hematopoietic Stem and Progenitor Cells. Mol. Ther. 2018, 26, 320–328. [Google Scholar] [CrossRef]
- Santoni de Sio, F.R.; Gritti, A.; Cascio, P.; Neri, M.; Sampaolesi, M.; Galli, C.; Luban, J.; Naldini, L. Lentiviral Vector Gene Transfer is Limited by the Proteasome at Postentry Steps in various Types of Stem Cells. Stem Cells 2008, 26, 2142–2152. [Google Scholar] [CrossRef]
- Santoni de Sio, F.R.; Cascio, P.; Zingale, A.; Gasparini, M.; Naldini, L. Proteasome Activity Restricts Lentiviral Gene Transfer into Hematopoietic Stem Cells and is Down-Regulated by Cytokines that Enhance Transduction. Blood 2006, 107, 4257–4265. [Google Scholar] [CrossRef]
- Vargas, J.; Gusella, G.L.; Najfeld, V.; Klotman, M.E.; Cara, A. Novel Integrase-Defective Lentiviral Episomal Vectors for Gene Transfer. Hum. Gene Ther. 2004, 15, 361–372. [Google Scholar] [CrossRef]
- Flynn, R.P.; Zacharias, J.; Zhou, X.; Cannon, M.L.; Philpott, N.J. Non-Integrating Lentiviral Vectors for Specific Killing of Epstein-Barr Virus Nuclear Antigen 1-Positive B Cell Lymphoma Cells. J. Gene Med. 2011, 13, 487–496. [Google Scholar] [CrossRef]
- Xu, Z.; Chen, F.; Zhang, L.; Lu, J.; Xu, P.; Liu, G.; Xie, X.; Mu, W.; Wang, Y.; Liu, D. Non-Integrating Lentiviral Vectors Based on the Minimal S/MAR Sequence Retain Transgene Expression in Dividing Cells. Sci. China Life Sci. 2016, 59, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Qi, X.; Zhang, R.; Wu, Z.Y.; Yan, C.E.; Li, J.; Liu, Q.Y.; Qi, J. Episomal Lentiviral Vectors Confer Erythropoietin Expression in Dividing Cells. Plasmid 2017, 90, 15–19. [Google Scholar] [CrossRef]
- Jin, C.; Fotaki, G.; Ramachandran, M.; Nilsson, B.; Essand, M.; Yu, D. Safe Engineering of CAR T Cells for Adoptive Cell Therapy of Cancer using Long-Term Episomal Gene Transfer. EMBO Mol. Med. 2016, 8, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Verghese, S.C.; Goloviznina, N.A.; Skinner, A.M.; Lipps, H.J.; Kurre, P. S/MAR Sequence Confers Long-Term Mitotic Stability on Non-Integrating Lentiviral Vector Episomes without Selection. Nucleic Acids Res. 2014, 42, e53. [Google Scholar] [CrossRef] [PubMed]
- Kymalainen, H.; Appelt, J.U.; Giordano, F.A.; Davies, A.F.; Ogilvie, C.M.; Ahmed, S.G.; Laufs, S.; Schmidt, M.; Bode, J.; Yanez-Munoz, R.J.; et al. Long-Term Episomal Transgene Expression from Mitotically Stable Integration-Deficient Lentiviral Vectors. Hum. Gene Ther. 2014, 25, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Vink, C.A.; Gaspar, H.B.; Gabriel, R.; Schmidt, M.; McIvor, R.S.; Thrasher, A.J.; Qasim, W. Sleeping Beauty Transposition from Nonintegrating Lentivirus. Mol. Ther. 2009, 17, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Moldt, B.; Staunstrup, N.H.; Jakobsen, M.; Yanez-Munoz, R.J.; Mikkelsen, J.G. Genomic Insertion of Lentiviral DNA Circles Directed by the Yeast Flp Recombinase. BMC Biotechnol. 2008, 8, 60. [Google Scholar] [CrossRef]
- Philippe, S.; Sarkis, C.; Barkats, M.; Mammeri, H.; Ladroue, C.; Petit, C.; Mallet, J.; Serguera, C. Lentiviral Vectors with a Defective Integrase Allow Efficient and Sustained Transgene Expression in Vitro and in Vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 17684–17689. [Google Scholar] [CrossRef]
- Peluffo, H.; Foster, E.; Ahmed, S.G.; Lago, N.; Hutson, T.H.; Moon, L.; Yip, P.; Wanisch, K.; Caraballo-Miralles, V.; Olmos, G.; et al. Efficient Gene Expression from Integration-Deficient Lentiviral Vectors in the Spinal Cord. Gene Ther. 2013, 20, 645–657. [Google Scholar] [CrossRef]
- Ahmed, S.G.; Waddington, S.N.; Boza-Moran, M.G.; Yanez-Munoz, R.J. High-Efficiency Transduction of Spinal Cord Motor Neurons by Intrauterine Delivery of Integration-Deficient Lentiviral Vectors. J. Control. Release 2018, 273, 99–107. [Google Scholar] [CrossRef]
- Rahim, A.A.; Wong, A.M.; Howe, S.J.; Buckley, S.M.; Acosta-Saltos, A.D.; Elston, K.E.; Ward, N.J.; Philpott, N.J.; Cooper, J.D.; Anderson, P.N.; et al. Efficient Gene Delivery to the Adult and Fetal CNS using Pseudotyped Non-Integrating Lentiviral Vectors. Gene Ther. 2009, 16, 509–520. [Google Scholar] [CrossRef]
- Lu-Nguyen, N.B.; Broadstock, M.; Schliesser, M.G.; Bartholomae, C.C.; von Kalle, C.; Schmidt, M.; Yanez-Munoz, R.J. Transgenic Expression of Human Glial Cell Line-Derived Neurotrophic Factor from Integration-Deficient Lentiviral Vectors is Neuroprotective in a Rodent Model of Parkinson’s Disease. Hum. Gene Ther. 2014, 25, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Suwanmanee, T.; Hu, G.; Gui, T.; Bartholomae, C.C.; Kutschera, I.; von Kalle, C.; Schmidt, M.; Monahan, P.E.; Kafri, T. Integration-Deficient Lentiviral Vectors Expressing Codon-Optimized R338L Human FIX Restore Normal Hemostasis in Hemophilia B Mice. Mol. Ther. 2014, 22, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Ocampo, D.; Cawen, F.A.; Alamo-Pindado, L.A.; Negro-Demontel, M.L.; Peluffo, H. Safe and Neuroprotective Vectors for Long-Term Traumatic Brain Injury Gene Therapy. Gene Ther. 2020, 27, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Karwacz, K.; Mukherjee, S.; Apolonia, L.; Blundell, M.P.; Bouma, G.; Escors, D.; Collins, M.K.; Thrasher, A.J. Nonintegrating Lentivector Vaccines Stimulate Prolonged T-Cell and Antibody Responses and are Effective in Tumor Therapy. J. Virol. 2009, 83, 3094–3103. [Google Scholar] [CrossRef] [PubMed]
- Daenthanasanmak, A.; Salguero, G.; Borchers, S.; Figueiredo, C.; Jacobs, R.; Sundarasetty, B.S.; Schneider, A.; Schambach, A.; Eiz-Vesper, B.; Blasczyk, R.; et al. Integrase-Defective Lentiviral Vectors Encoding Cytokines Induce Differentiation of Human Dendritic Cells and Stimulate Multivalent Immune Responses in Vitro and in Vivo. Vaccine 2012, 30, 5118–5131. [Google Scholar] [CrossRef] [PubMed]
- Negri, D.R.; Michelini, Z.; Baroncelli, S.; Spada, M.; Vendetti, S.; Buffa, V.; Bona, R.; Leone, P.; Klotman, M.E.; Cara, A. Successful Immunization with a Single Injection of Non-Integrating Lentiviral Vector. Mol. Ther. 2007, 15, 1716–1723. [Google Scholar] [CrossRef]
- Hu, B.; Dai, B.; Wang, P. Vaccines Delivered by Integration-Deficient Lentiviral Vectors Targeting Dendritic Cells Induces Strong Antigen-Specific Immunity. Vaccine 2010, 28, 6675–6683. [Google Scholar] [CrossRef][Green Version]
- Negri, D.R.; Michelini, Z.; Baroncelli, S.; Spada, M.; Vendetti, S.; Bona, R.; Leone, P.; Klotman, M.E.; Cara, A. Nonintegrating Lentiviral Vector-Based Vaccine Efficiently Induces Functional and Persistent CD8+ T Cell Responses in Mice. J. Biomed. Biotechnol. 2010, 2010, 534501. [Google Scholar] [CrossRef]
- Blasi, M.; Negri, D.; LaBranche, C.; Alam, S.M.; Baker, E.J.; Brunner, E.C.; Gladden, M.A.; Michelini, Z.; Vandergrift, N.A.; Wiehe, K.J.; et al. IDLV-HIV-1 Env Vaccination in Non-Human Primates Induces Affinity Maturation of Antigen-Specific Memory B Cells. Commun. Biol. 2018, 1, 134–136, eCollection 2018. [Google Scholar] [CrossRef]
- Gallinaro, A.; Borghi, M.; Pirillo, M.F.; Cecchetti, S.; Bona, R.; Canitano, A.; Michelini, Z.; Di Virgilio, A.; Olvera, A.; Brander, C.; et al. Development and Preclinical Evaluation of an Integrase Defective Lentiviral Vector Vaccine Expressing the HIVACAT T Cell Immunogen in Mice. Mol. Ther. Methods Clin. Dev. 2020, 17, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Wee, E.G.; Ondondo, B.; Berglund, P.; Archer, J.; McMichael, A.J.; Baltimore, D.; Ter Meulen, J.H.; Hanke, T. HIV-1 Conserved Mosaics Delivered by Regimens with Integration-Deficient DC-Targeting Lentiviral Vector Induce Robust T Cells. Mol. Ther. 2017, 25, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Fontana, J.M.; Christos, P.J.; Michelini, Z.; Negri, D.; Cara, A.; Salvatore, M. Mucosal Immunization with Integrase-Defective Lentiviral Vectors Protects Against Influenza Virus Challenge in Mice. PLoS ONE 2014, 9, e97270. [Google Scholar] [CrossRef] [PubMed]
- Negri, D.R.; Bona, R.; Michelini, Z.; Leone, P.; Macchia, I.; Klotman, M.E.; Salvatore, M.; Cara, A. Transduction of Human Antigen-Presenting Cells with Integrase-Defective Lentiviral Vector Enables Functional Expansion of Primed Antigen-Specific CD8(+) T Cells. Hum. Gene Ther. 2010, 21, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Gallinaro, A.; Borghi, M.; Bona, R.; Grasso, F.; Calzoletti, L.; Palladino, L.; Cecchetti, S.; Vescio, M.F.; Macchia, D.; Morante, V.; et al. Integrase Defective Lentiviral Vector as a Vaccine Platform for Delivering Influenza Antigens. Front. Immunol. 2018, 9, 171. [Google Scholar] [CrossRef]
- Ku, M.W.; Anna, F.; Souque, P.; Petres, S.; Prot, M.; Simon-Loriere, E.; Charneau, P.; Bourgine, M. A Single Dose of NILV-Based Vaccine Provides Rapid and Durable Protection Against Zika Virus. Mol. Ther. 2020, 28, 1772–1782. [Google Scholar] [CrossRef]
- Grasso, F.; Negri, D.R.; Mochi, S.; Rossi, A.; Cesolini, A.; Giovannelli, A.; Chiantore, M.V.; Leone, P.; Giorgi, C.; Cara, A. Successful Therapeutic Vaccination with Integrase Defective Lentiviral Vector Expressing Nononcogenic Human Papillomavirus E7 Protein. Int. J. Cancer 2013, 132, 335–344. [Google Scholar] [CrossRef]
- Odegard, J.M.; Kelley-Clarke, B.; Tareen, S.U.; Campbell, D.J.; Flynn, P.A.; Nicolai, C.J.; Slough, M.M.; Vin, C.D.; McGowan, P.J.; Nelson, L.T.; et al. Virological and Preclinical Characterization of a Dendritic Cell Targeting, Integration-Deficient Lentiviral Vector for Cancer Immunotherapy. J. Immunother. 2015, 38, 41–53. [Google Scholar] [CrossRef]
- Coutant, F.; Frenkiel, M.P.; Despres, P.; Charneau, P. Protective Antiviral Immunity Conferred by a Nonintegrative Lentiviral Vector-Based Vaccine. PLoS ONE 2008, 3, e3973. [Google Scholar] [CrossRef]
- Deng, Y.; Guan, J.; Wen, B.; Zhu, N.; Chen, H.; Song, J.; Yang, Y.; Wang, Y.; Tan, W. Induction of Broadly Neutralising HCV Antibodies in Mice by Integration-Deficient Lentiviral Vector-Based Pseudotyped Particles. PLoS ONE 2013, 8, e62684. [Google Scholar] [CrossRef]
- Coutant, F.; Sanchez David, R.Y.; Felix, T.; Boulay, A.; Caleechurn, L.; Souque, P.; Thouvenot, C.; Bourgouin, C.; Beignon, A.S.; Charneau, P. A Nonintegrative Lentiviral Vector-Based Vaccine Provides Long-Term Sterile Protection Against Malaria. PLoS ONE 2012, 7, e48644. [Google Scholar] [CrossRef]
- Tareen, S.U.; Kelley-Clarke, B.; Nicolai, C.J.; Cassiano, L.A.; Nelson, L.T.; Slough, M.M.; Vin, C.D.; Odegard, J.M.; Sloan, D.D.; Van Hoeven, N.; et al. Design of a Novel Integration-Deficient Lentivector Technology that Incorporates Genetic and Posttranslational Elements to Target Human Dendritic Cells. Mol. Ther. 2014, 22, 575–587. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pollack, S.M.; Lu, H.; Gnjatic, S.; Somaiah, N.; O’Malley, R.B.; Jones, R.L.; Hsu, F.J.; Ter Meulen, J. First-in-Human Treatment with a Dendritic Cell-Targeting Lentiviral Vector-Expressing NY-ESO-1, LV305, Induces Deep, Durable Response in Refractory Metastatic Synovial Sarcoma Patient. J. Immunother. 2017, 40, 302–306. [Google Scholar] [CrossRef] [PubMed]
- Pollack, S.M. The Potential of the CMB305 Vaccine Regimen to Target NY-ESO-1 and Improve Outcomes for Synovial Sarcoma and Myxoid/Round Cell Liposarcoma Patients. Expert Rev. Vaccines 2018, 17, 107–114. [Google Scholar] [CrossRef]
- Somaiah, N.; Block, M.S.; Kim, J.W.; Shapiro, G.I.; Do, K.T.; Hwu, P.; Eder, J.P.; Jones, R.L.; Lu, H.; Ter Meulen, J.H.; et al. First-in-Class, First-in-Human Study Evaluating LV305, a Dendritic-Cell Tropic Lentiviral Vector, in Sarcoma and Other Solid Tumors Expressing NY-ESO-1. Clin. Cancer Res. 2019, 25, 5808–5817. [Google Scholar] [CrossRef]
- Albershardt, T.C.; Campbell, D.J.; Parsons, A.J.; Slough, M.M.; Ter Meulen, J.; Berglund, P. LV305, a Dendritic Cell-Targeting Integration-Deficient ZVex(TM)-Based Lentiviral Vector Encoding NY-ESO-1, Induces Potent Anti-Tumor Immune Response. Mol. Ther. Oncolytics 2016, 3, 16010. [Google Scholar] [CrossRef]
- Lin, B.; Gao, A.; Zhang, R.; Ma, H.; Shen, H.; Hu, Q.; Zhang, H.; Zhao, M.; Lan, X.; Liu, K. Use of a Novel Integrase-Deficient Lentivirus for Targeted Anti-Cancer Therapy with Survivin Promoter-Driven Diphtheria Toxin A. Medicine 2015, 94, e1301. [Google Scholar] [CrossRef]
- Okada, Y.; Ueshin, Y.; Hasuwa, H.; Takumi, K.; Okabe, M.; Ikawa, M. Targeted Gene Modification in Mouse ES Cells using Integrase-Defective Lentiviral Vectors. Genesis 2009, 47, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, A.; Cesana, D.; Genovese, P.; Di Stefano, B.; Provasi, E.; Colombo, D.F.; Neri, M.; Magnani, Z.; Cantore, A.; Lo Riso, P.; et al. Site-Specific Integration and Tailoring of Cassette Design for Sustainable Gene Transfer. Nat. Methods 2011, 8, 861–869. [Google Scholar] [CrossRef]
- Izmiryan, A.; Basmaciogullari, S.; Henry, A.; Paques, F.; Danos, O. Efficient Gene Targeting Mediated by a Lentiviral Vector-Associated Meganuclease. Nucleic Acids Res. 2011, 39, 7610–7619. [Google Scholar] [CrossRef]
- Joglekar, A.V.; Hollis, R.P.; Kuftinec, G.; Senadheera, S.; Chan, R.; Kohn, D.B. Integrase-Defective Lentiviral Vectors as a Delivery Platform for Targeted Modification of Adenosine Deaminase Locus. Mol. Ther. 2013, 21, 1705–1717. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Razak, H.H.; Rocca, C.J.; Howe, S.J.; Alonso-Ferrero, M.E.; Wang, J.; Gabriel, R.; Bartholomae, C.C.; Gan, C.H.V.; Garin, M.I.; Roberts, A.; et al. Molecular Evidence of Genome Editing in a Mouse Model of Immunodeficiency. Sci. Rep. 2018, 8, 8214–8219. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, A.; Genovese, P.; Beausejour, C.M.; Colleoni, S.; Lee, Y.L.; Kim, K.A.; Ando, D.; Urnov, F.D.; Galli, C.; Gregory, P.D.; et al. Gene Editing in Human Stem Cells using Zinc Finger Nucleases and Integrase-Defective Lentiviral Vector Delivery. Nat. Biotechnol. 2007, 25, 1298–1306. [Google Scholar] [CrossRef] [PubMed]
- Shariati, L.; Modarressi, M.H.; Tabatabaiefar, M.A.; Kouhpayeh, S.; Hejazi, Z.; Shahbazi, M.; Sabzehei, F.; Salehi, M.; Khanahmad, H. Engineered Zinc-Finger Nuclease to Generate Site-Directed Modification in the KLF1 Gene for Fetal Hemoglobin Induction. J. Cell. Biochem. 2018. [Google Scholar] [CrossRef]
- Popplewell, L.; Koo, T.; Leclerc, X.; Duclert, A.; Mamchaoui, K.; Gouble, A.; Mouly, V.; Voit, T.; Paques, F.; Cedrone, F.; et al. Gene Correction of a Duchenne Muscular Dystrophy Mutation by Meganuclease-Enhanced Exon Knock-In. Hum. Gene Ther. 2013, 24, 692–701. [Google Scholar] [CrossRef]
- Mali, P.; Ye, Z.; Hommond, H.H.; Yu, X.; Lin, J.; Chen, G.; Zou, J.; Cheng, L. Improved Efficiency and Pace of Generating Induced Pluripotent Stem Cells from Human Adult and Fetal Fibroblasts. Stem Cells 2008, 26, 1998–2005. [Google Scholar] [CrossRef]
- Yang, G.; Si-Tayeb, K.; Corbineau, S.; Vernet, R.; Gayon, R.; Dianat, N.; Martinet, C.; Clay, D.; Goulinet-Mainot, S.; Tachdjian, G.; et al. Integration-Deficient Lentivectors: An Effective Strategy to Purify and Differentiate Human Embryonic Stem Cell-Derived Hepatic Progenitors. BMC Biol. 2013, 11, 86. [Google Scholar] [CrossRef]
- Hamilton, A.M.; Foster, P.J.; Ronald, J.A. Evaluating Nonintegrating Lentiviruses as Safe Vectors for Noninvasive Reporter-Based Molecular Imaging of Multipotent Mesenchymal Stem Cells. Hum. Gene Ther. 2018, 29, 1213–1225. [Google Scholar] [CrossRef]
- Ortinski, P.I.; O’Donovan, B.; Dong, X.; Kantor, B. Integrase-Deficient Lentiviral Vector as an all-in-One Platform for Highly Efficient CRISPR/Cas9-Mediated Gene Editing. Mol. Ther. Methods Clin. Dev. 2017, 5, 153–164. [Google Scholar] [CrossRef]
- Surun, D.; Schwable, J.; Tomasovic, A.; Ehling, R.; Stein, S.; Kurrle, N.; von Melchner, H.; Schnutgen, F. High Efficiency Gene Correction in Hematopoietic Cells by Donor-Template-Free CRISPR/Cas9 Genome Editing. Mol. Ther. Nucleic Acids 2018, 10, 1–8. [Google Scholar] [CrossRef]
- Izmiryan, A.; Ganier, C.; Bovolenta, M.; Schmitt, A.; Mavilio, F.; Hovnanian, A. Ex Vivo COL7A1 Correction for Recessive Dystrophic Epidermolysis Bullosa using CRISPR/Cas9 and Homology-Directed Repair. Mol. Ther. Nucleic Acids 2018, 12, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Schokrpur, S.; Archang, M.; Hermann, K.; Sharrow, A.C.; Khanna, P.; Novak, J.; Signoretti, S.; Bhatt, R.S.; Knudsen, B.S.; et al. A Non-Integrating Lentiviral Approach Overcomes Cas9-Induced Immune Rejection to Establish an Immunocompetent Metastatic Renal Cancer Model. Mol. Ther. Methods Clin. Dev. 2018, 9, 203–210. [Google Scholar] [CrossRef] [PubMed]

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apolonia, L. The Old and the New: Prospects for Non-Integrating Lentiviral Vector Technology. Viruses 2020, 12, 1103. https://doi.org/10.3390/v12101103
Apolonia L. The Old and the New: Prospects for Non-Integrating Lentiviral Vector Technology. Viruses. 2020; 12(10):1103. https://doi.org/10.3390/v12101103
Chicago/Turabian StyleApolonia, Luis. 2020. "The Old and the New: Prospects for Non-Integrating Lentiviral Vector Technology" Viruses 12, no. 10: 1103. https://doi.org/10.3390/v12101103
APA StyleApolonia, L. (2020). The Old and the New: Prospects for Non-Integrating Lentiviral Vector Technology. Viruses, 12(10), 1103. https://doi.org/10.3390/v12101103