Electron Microscopy in Discovery of Novel and Emerging Viruses from the Collection of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA)

 and
and 
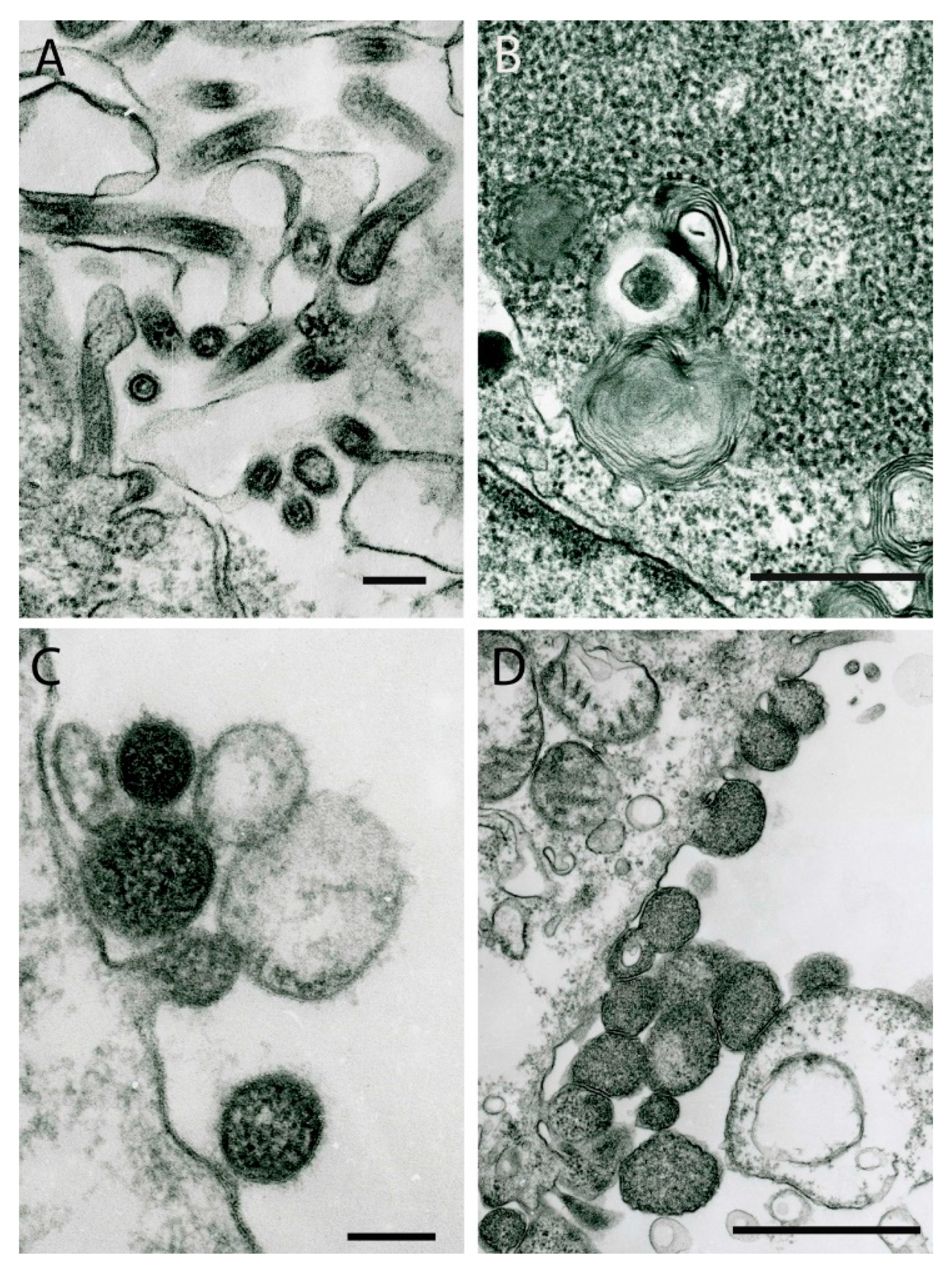{kind=link}
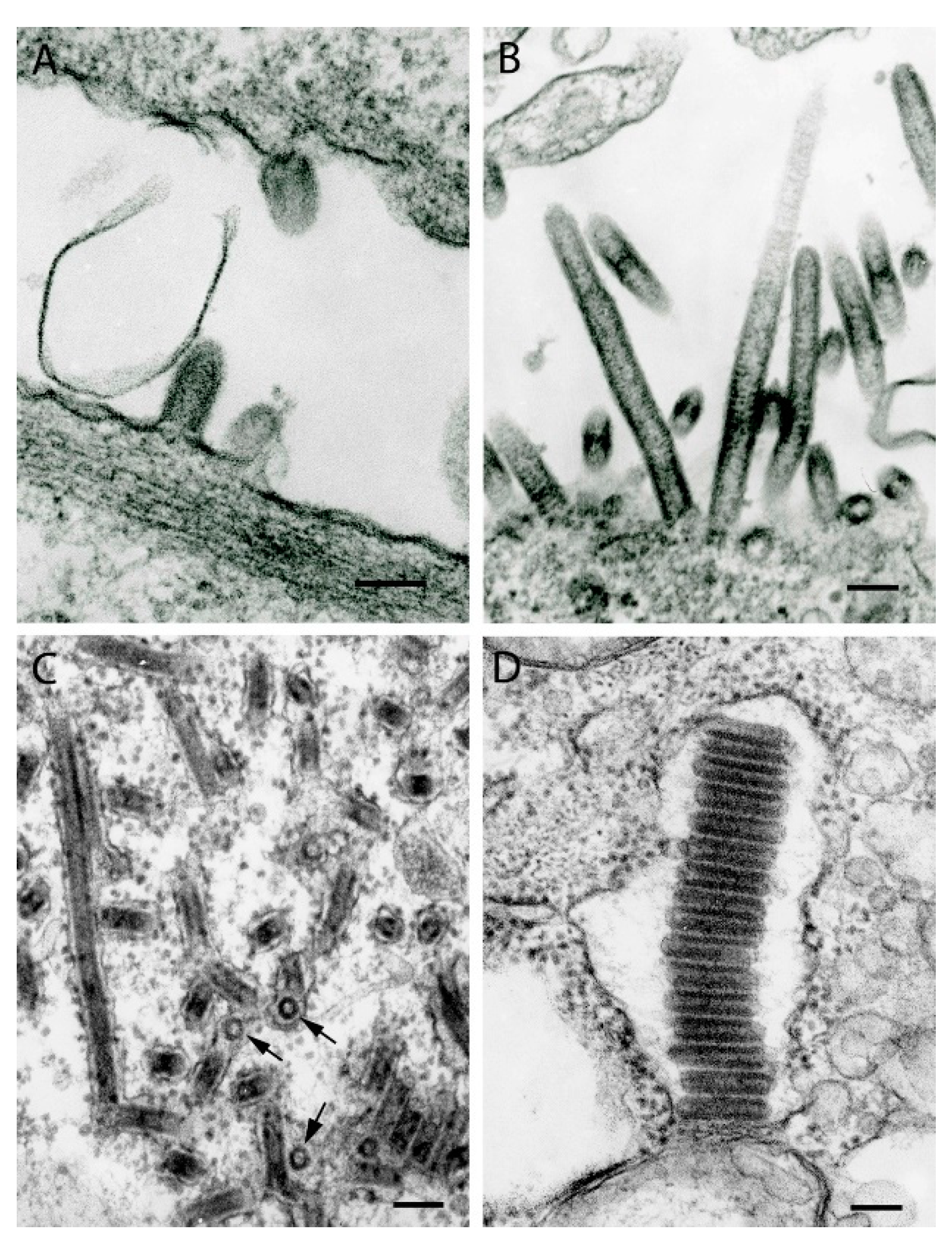{kind=link}
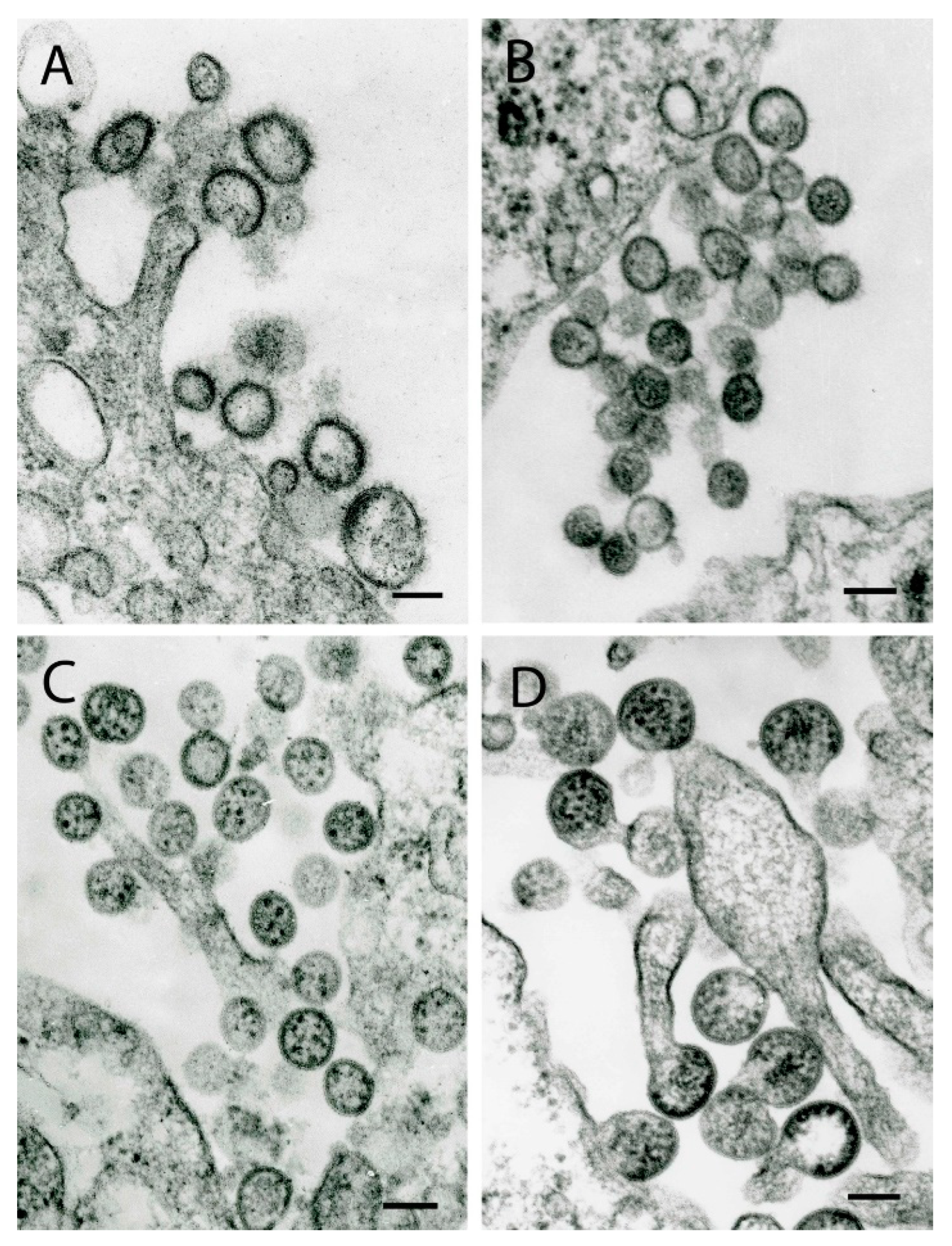{kind=link}
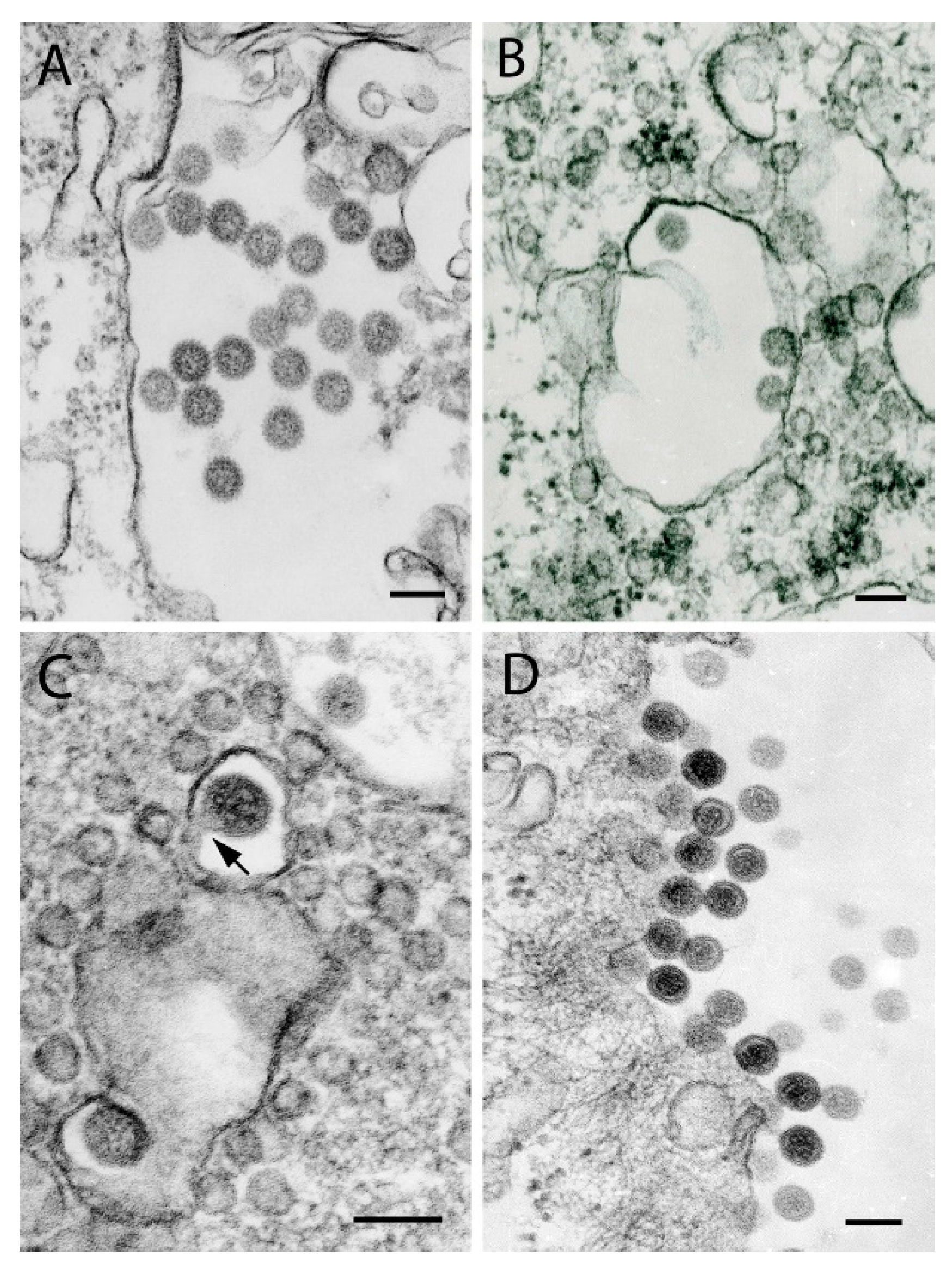{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
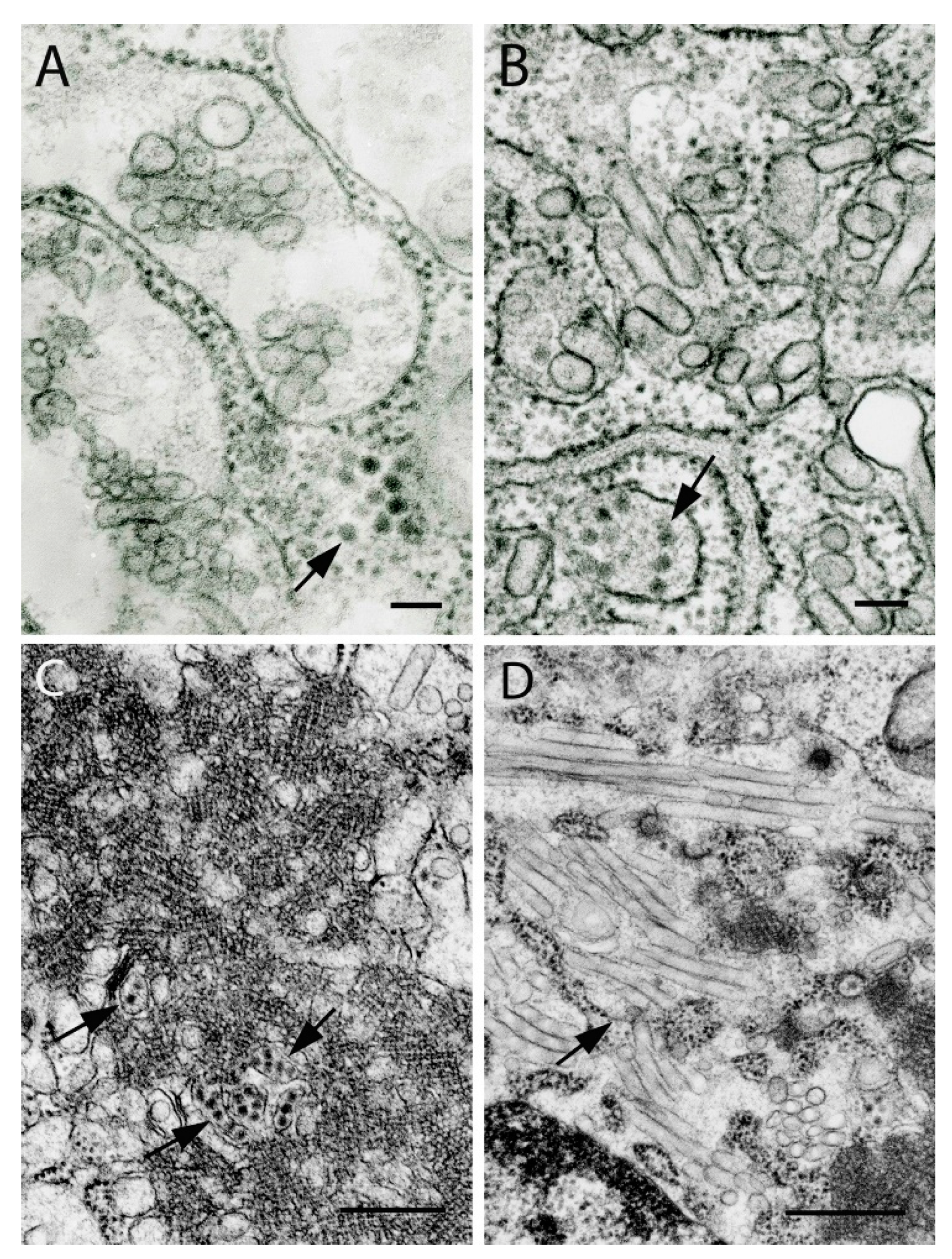
1. Paramyxoviruses
2. Rhabdoviruses
3. Nyamiviruses
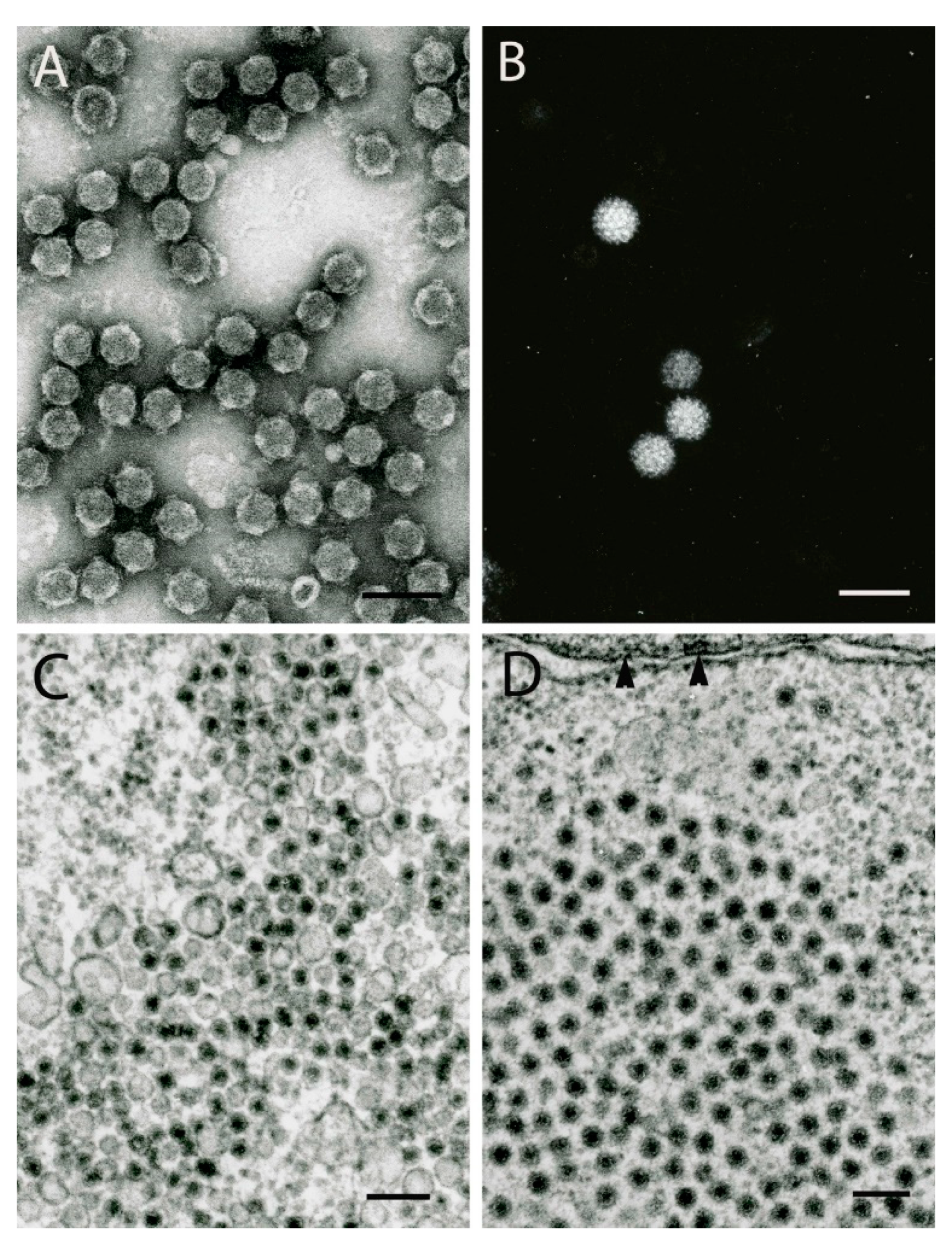
4. Orthomyxoviruses
5. Phleboviruses
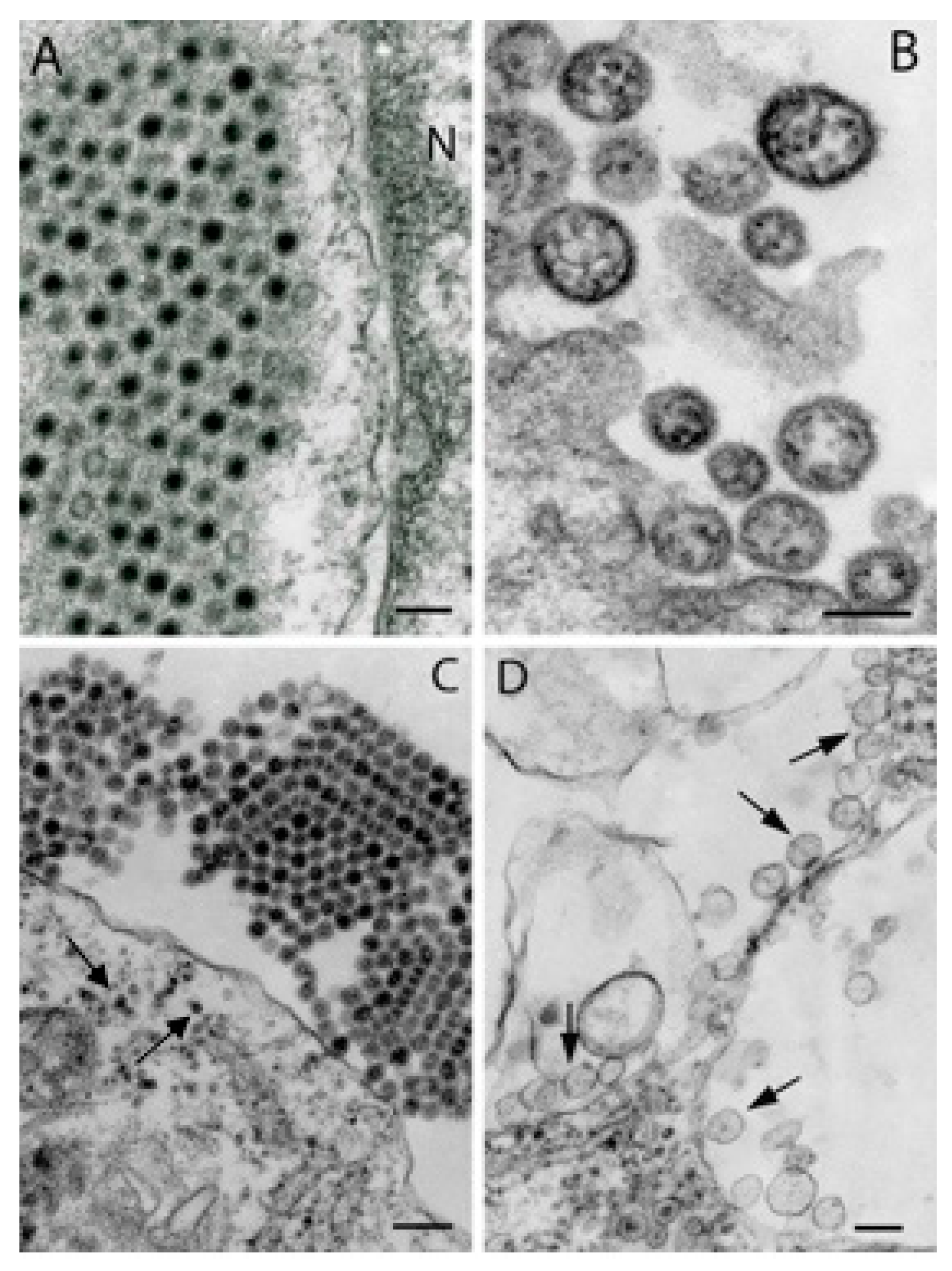
6. Flaviviruses
7. Reoviruses
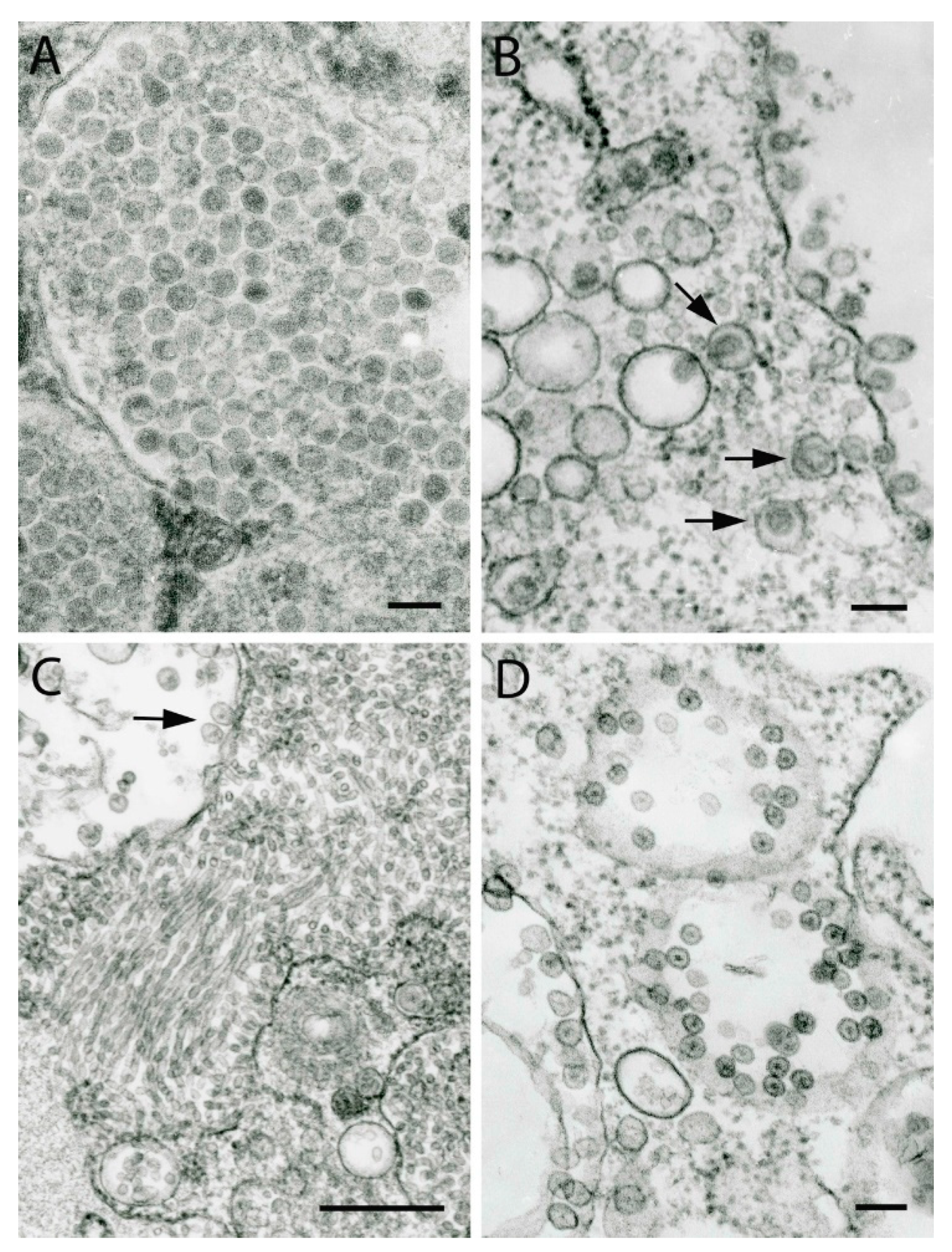
8. Arenaviruses
9. Alphaviruses
10. Mesoniviruses
11. Negeviruses
12. Mixed Infections
13. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, F.A. The Foundations of Virology - Discoverers and Discoveries, Inventors and Inventions, Developers and Technologies; Infinity Publishing: West Conshohocken, PA, USA, 2012. [Google Scholar]
- Miller, S.E. Electron microscopy of viral infections. In Laboratory Diagnosis of Viral Infections, 3rd ed.; Lennette, E.H., Smith, T.F., Eds.; Marcel Dekker: New York, NY, USA, 1999; pp. 45–70. [Google Scholar]
- Miller, S.E. Diagnosis of viral infections by electron microscopy. In Diagnostic Procedures for Viral, Rickettsial and Chlamydial Infections, 7th ed.; Lennette, E.H., Lennette, D.A., Lennette, E.T., Eds.; American Public Health Association: Washington, DC, USA, 1995; pp. 37–78. [Google Scholar]
- Hsiung, G.D.; Fong, C.K.Y.; Landry, M.L. Hsiung’s Diagnostic Virology as Illustrated by Light and Electron Microscopy, 4th ed.; Yale University Press: New Haven, CT, USA, 1994; p. 404. [Google Scholar]
- Adams, J.R.; Bonami, J.R. Atlas of Invertebrate Viruses; CRC Press: Boca Raton, FL, USA, 1991. [Google Scholar]
- Palmer, E.L.; Martin, M.L. Electron Microscopy in Viral Diagnosis; CRC Press: Boca Raton, FL, USA, 1988. [Google Scholar]
- Palmer, E.L.; Martin, M.L. An Atlas of Mammalian Viruses; CRC Press: Boca Raton, FL, USA, 1982. [Google Scholar]
- Doane, F.W.; Anderson, N. Electron Microscopy in Diagnostic Virology: A Practical Guide and Atlas; Cambridge University Press: New York, NY, USA, 1987. [Google Scholar]
- Maramorosch, K. The Atlas of Insect and Plant Viruses: Including Mycoplasmaviruses and Viroids; Academic Press: New York, NY, USA, 1978; Volume 8. [Google Scholar]
- Dalton, A.J.; Haguenau, F. Ultrastructure of Animal Viruses and Bacteriophages: An Atlas; Academic Press: New York, NY, USA, 1973; Volume 5. [Google Scholar]
- Avakian, A.A.; Bykovsky, A.F. Atlas of Anatomy and Ontogenesis of Human and Animal Viruses; Meditsina: Moscow, Russia, 1970. (In Russian) [Google Scholar]
- Madeley, C.R.; Field, A.M. Virus Morphology, 2nd ed.; Churchill Livingstone: Edinburgh, UK, 1988. [Google Scholar]
- Goldsmith, C.S.; Miller, S.E. Modern uses of electron microscopy for detection of viruses. Clin. Microbiol. Rev. 2009, 22, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, C.S.; Ksiazek, T.G.; Rollin, P.E.; Comer, J.A.; Nicholson, W.L.; Peret, T.C.; Erdman, D.D.; Bellini, W.J.; Harcourt, B.H.; Rota, P.A.; et al. Cell culture and electron microscopy for identifying viruses in diseases of unknown cause. Emerg. Infect. Dis. 2013, 19, 886–891. [Google Scholar] [CrossRef]
- Goldsmith, C.S. Morphologic differentiation of viruses beyond the family level. Viruses 2014, 6, 4902–4913. [Google Scholar] [CrossRef]
- Richert-Poggeler, K.R.; Franzke, K.; Hipp, K.; Kleespies, R.G. Electron microscopy methods for virus diagnosis and high resolution analysis of viruses. Front. Microbiol. 2018, 9, 3255. [Google Scholar] [CrossRef]
- Mann, B.R.; McMullen, A.R.; Swetnam, D.M.; Salvato, V.; Reyna, M.; Guzman, H.; Bueno, R., Jr.; Dennett, J.A.; Tesh, R.B.; Barrett, A.D. Continued evolution of west nile virus, houston, texas, USA, 2002–2012. Emerg. Infect. Dis. 2013, 19, 1418–1427. [Google Scholar] [CrossRef]
- Dennett, J.A.; Bala, A.; Wuithiranyagool, T.; Randle, Y.; Sargent, C.B.; Guzman, H.; Siirin, M.; Hassan, H.K.; Reyna-Nava, M.; Unnasch, T.R.; et al. Associations between two mosquito populations and west nile virus in harris county, texas, 2003–2006. J. Am. Mosq. Control Assoc. 2007, 23, 264–275. [Google Scholar] [CrossRef]
- Meyer, T.E.; Bull, L.M.; Cain Holmes, K.; Pascua, R.F.; Travassos da Rosa, A.; Gutierrez, C.R.; Corbin, T.; Woodward, J.L.; Taylor, J.P.; Tesh, R.B.; et al. West nile virus infection among the homeless, houston, texas. Emerg. Infect. Dis. 2007, 13, 1500–1503. [Google Scholar] [CrossRef]
- Matsuno, K.; Weisend, C.; Kajihara, M.; Matysiak, C.; Williamson, B.N.; Simuunza, M.; Mweene, A.S.; Takada, A.; Tesh, R.B.; Ebihara, H. Comprehensive molecular detection of tick-borne phleboviruses leads to the retrospective identification of taxonomically unassigned bunyaviruses and the discovery of a novel member of the genus phlebovirus. J. Virol. 2015, 89, 594–604. [Google Scholar] [CrossRef]
- Matsuno, K.; Weisend, C.; Travassos da Rosa, A.P.; Anzick, S.L.; Dahlstrom, E.; Porcella, S.F.; Dorward, D.W.; Yu, X.J.; Tesh, R.B.; Ebihara, H. Characterization of the bhanja serogroup viruses (bunyaviridae): A novel species of the genus phlebovirus and its relationship with other emerging tick-borne phleboviruses. J. Virol. 2013, 87, 3719–3728. [Google Scholar] [CrossRef] [PubMed]
- Briese, T.; Chowdhary, R.; Travassos da Rosa, A.; Hutchison, S.K.; Popov, V.; Street, C.; Tesh, R.B.; Lipkin, W.I. Upolu virus and aransas bay virus, two presumptive bunyaviruses, are novel members of the family orthomyxoviridae. J. Virol. 2014, 88, 5298–5309. [Google Scholar] [CrossRef]
- Contreras-Gutierrez, M.A.; Nunes, M.R.T.; Guzman, H.; Uribe, S.; Suaza Vasco, J.D.; Cardoso, J.F.; Popov, V.L.; Widen, S.G.; Wood, T.G.; Vasilakis, N.; et al. Sinu virus, a novel and divergent orthomyxovirus related to members of the genus thogotovirus isolated from mosquitoes in colombia. Virology 2017, 501, 166–175. [Google Scholar] [CrossRef]
- Lima, J.A.; Nunes Neto, J.P.; Castro, K.S.; Travassos da Rosa, A.P.A.; Tesh, R.; Nunes, M.R.T.; Popov, V.L.; Vasilakis, N.; Guzman, H.; Widen, S.; et al. Characterization of triniti virus supports its reclassification in the family peribunyaviridae. J. Gen. Virol. 2019, 100, 137–144. [Google Scholar] [CrossRef]
- Chiang, J.O.; Marciel de Souza, W.; Teixeira Nunes, M.R.; Acrani, G.O.; Paes de Andrade Travassos da Rosa, A.; Mesquita de Freitas, N.; Patroca da Silva, S.; Dorta da Silva, P.H.; Watanabe de Sousa, A.; Rodrigues, S.G.; et al. Characterization of the gamboa virus serogroup (orthobunyavirus genus, peribunyaviridae family). Am. J. Trop. Med. Hyg. 2018, 98, 1502–1511. [Google Scholar] [CrossRef]
- Hontz, R.D.; Guevara, C.; Halsey, E.S.; Silvas, J.; Santiago, F.W.; Widen, S.G.; Wood, T.G.; Casanova, W.; Vasilakis, N.; Watts, D.M.; et al. Itaya virus, a novel orthobunyavirus associated with human febrile illness, peru. Emerg. Infect. Dis. 2015, 21, 781–788. [Google Scholar] [CrossRef]
- Rogers, M.B.; Gulino, K.M.; Tesh, R.B.; Cui, L.; Fitch, A.; Unnasch, T.R.; Popov, V.L.; Travassos da Rosa, A.P.A.; Guzman, H.; Carrera, J.P.; et al. Characterization of five unclassified orthobunyaviruses (bunyaviridae) from africa and the americas. J. Gen. Virol. 2017, 98, 2258–2266. [Google Scholar] [CrossRef]
- Rogers, M.B.; Cui, L.; Fitch, A.; Popov, V.; Travassos da Rosa, A.P.; Vasilakis, N.; Tesh, R.B.; Ghedin, E. Whole genome analysis of sierra nevada virus, a novel mononegavirus in the family nyamiviridae. Am. J. Trop. Med. Hyg. 2014, 91, 159–164. [Google Scholar] [CrossRef]
- Auguste, A.J.; Kaelber, J.T.; Fokam, E.B.; Guzman, H.; Carrington, C.V.; Erasmus, J.H.; Kamgang, B.; Popov, V.L.; Jakana, J.; Liu, X.; et al. A newly isolated reovirus has the simplest genomic and structural organization of any reovirus. J. Virol. 2015, 89, 676–687. [Google Scholar] [CrossRef]
- Contreras-Gutierrez, M.A.; Guzman, H.; Cardoso, J.F.; Popov, V.L.; Nunes, M.R.T.; Uribe, S.; Widen, S.G.; Wood, T.G.; Vasilakis, N.; Tesh, R.B. Genome sequence of chiqui virus, a novel reovirus isolated from mosquitoes collected in colombia. Microbiol. Resour. Announc. 2018, 7. [Google Scholar] [CrossRef]
- Forrester, N.L.; Widen, S.G.; Wood, T.G.; Travassos da Rosa, A.P.; Ksiazek, T.G.; Vasilakis, N.; Tesh, R.B. Identification of a new newcastle disease virus isolate from indonesia represents an ancestral lineage of class ii genotype xiii. Virus Genes 2013, 47, 168–172. [Google Scholar] [CrossRef]
- Nasar, F.; Palacios, G.; Gorchakov, R.V.; Guzman, H.; Da Rosa, A.P.; Savji, N.; Popov, V.L.; Sherman, M.B.; Lipkin, W.I.; Tesh, R.B.; et al. Eilat virus, a unique alphavirus with host range restricted to insects by rna replication. Proc. Natl. Acad. Sci. USA 2012, 109, 14622–14627. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.D.; Vasilakis, N.; Tian, J.H.; Li, C.X.; Chen, L.J.; Eastwood, G.; Diao, X.N.; Chen, M.H.; Chen, X.; et al. Divergent viruses discovered in arthropods and vertebrates revise the evolutionary history of the flaviviridae and related viruses. J. Virol. 2016, 90, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Vasilakis, N.; Guzman, H.; Widen, S.G.; Wood, T.G.; Popov, V.L.; Thangamani, S.; Tesh, R.B. Insect-specific viruses detected in laboratory mosquito colonies and their potential implications for experiments evaluating arbovirus vector competence. Am. J. Trop. Med. Hyg. 2015, 92, 422–428. [Google Scholar] [CrossRef]
- Carrera, J.P.; Guzman, H.; Beltran, D.; Diaz, Y.; Lopez-Verges, S.; Torres-Cosme, R.; Popov, V.; Widen, S.G.; Wood, T.G.; Weaver, S.C.; et al. Mercadeo virus: A novel mosquito-specific flavivirus from panama. Am. J. Trop. Med. Hyg. 2015, 93, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Guzman, H.; Contreras-Gutierrez, M.A.; Travassos da Rosa, A.P.A.; Nunes, M.R.T.; Cardoso, J.F.; Popov, V.L.; Young, K.I.; Savit, C.; Wood, T.G.; Widen, S.G.; et al. Characterization of three new insect-specific flaviviruses: Their relationship to the mosquito-borne flavivirus pathogens. Am. J. Trop. Med. Hyg. 2018, 98, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Popov, V.; Guzman, H.; Phan, T.G.; Vasilakis, N.; Tesh, R.; Delwart, E. Genomes of viral isolates derived from different mosquitos species. Virus Res. 2017, 242, 49–57. [Google Scholar] [CrossRef]
- Blasdell, K.R.; Guzman, H.; Widen, S.G.; Firth, C.; Wood, T.G.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N.; Walker, P.J. Ledantevirus: A proposed new genus in the rhabdoviridae has a strong ecological association with bats. Am. J. Trop. Med. Hyg. 2015, 92, 405–410. [Google Scholar] [CrossRef]
- Blasdell, K.R.; Widen, S.G.; Diviney, S.M.; Firth, C.; Wood, T.G.; Guzman, H.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N.; Walker, P.J. Koolpinyah and yata viruses: Two newly recognised ephemeroviruses from tropical regions of australia and africa. Vet. Microbiol. 2014, 174, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Contreras, M.A.; Eastwood, G.; Guzman, H.; Popov, V.; Savit, C.; Uribe, S.; Kramer, L.D.; Wood, T.G.; Widen, S.G.; Fish, D.; et al. Almendravirus: A proposed new genus of rhabdoviruses isolated from mosquitoes in tropical regions of the americas. Am. J. Trop. Med. Hyg. 2017, 96, 100–109. [Google Scholar] [CrossRef]
- Ghedin, E.; Rogers, M.B.; Widen, S.G.; Guzman, H.; Travassos da Rosa, A.P.; Wood, T.G.; Fitch, A.; Popov, V.; Holmes, E.C.; Walker, P.J.; et al. Kolente virus, a rhabdovirus species isolated from ticks and bats in the republic of guinea. J. Gen. Virol. 2013, 94, 2609–2615. [Google Scholar] [CrossRef]
- Palacios, G.; Forrester, N.L.; Savji, N.; Travassos da Rosa, A.P.; Guzman, H.; Detoy, K.; Popov, V.L.; Walker, P.J.; Lipkin, W.I.; Vasilakis, N.; et al. Characterization of farmington virus, a novel virus from birds that is distantly related to members of the family rhabdoviridae. Virol. J. 2013, 10, e219. [Google Scholar] [CrossRef]
- Vasilakis, N.; Castro-Llanos, F.; Widen, S.G.; Aguilar, P.V.; Guzman, H.; Guevara, C.; Fernandez, R.; Auguste, A.J.; Wood, T.G.; Popov, V.; et al. Arboretum and puerto almendras viruses: Two novel rhabdoviruses isolated from mosquitoes in peru. J. Gen. Virol. 2014, 95, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Widen, S.; Mayer, S.V.; Seymour, R.; Wood, T.G.; Popov, V.; Guzman, H.; Travassos da Rosa, A.P.; Ghedin, E.; Holmes, E.C.; et al. Niakha virus: A novel member of the family rhabdoviridae isolated from phlebotomine sandflies in senegal. Virology 2013, 444, 80–89. [Google Scholar] [CrossRef]
- Vasilakis, N.; Widen, S.; Travassos da Rosa, A.P.; Wood, T.G.; Walker, P.J.; Holmes, E.C.; Tesh, R.B. Malpais spring virus is a new species in the genus vesiculovirus. Virol. J. 2013, 10, e69. [Google Scholar] [CrossRef]
- Vasilakis, N.; Tesh, R.B.; Widen, S.G.; Mirchandani, D.; Walker, P.J. Genomic characterisation of cuiaba and charleville viruses: Arboviruses (family rhabdoviridae, genus sripuvirus) infecting reptiles and amphibians. Virus Genes 2019, 55, 87–94. [Google Scholar] [CrossRef]
- Walker, P.J.; Firth, C.; Widen, S.G.; Blasdell, K.R.; Guzman, H.; Wood, T.G.; Paradkar, P.N.; Holmes, E.C.; Tesh, R.B.; Vasilakis, N. Evolution of genome size and complexity in the rhabdoviridae. PLoS Pathog. 2015, 11, e1004664. [Google Scholar] [CrossRef]
- Vasilakis, N.; Guzman, H.; Firth, C.; Forrester, N.L.; Widen, S.G.; Wood, T.G.; Rossi, S.L.; Ghedin, E.; Popov, V.; Blasdell, K.R.; et al. Mesoniviruses are mosquito-specific viruses with extensive geographic distribution and host range. Virol. J. 2014, 11, e97. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Forrester, N.L.; Palacios, G.; Nasar, F.; Savji, N.; Rossi, S.L.; Guzman, H.; Wood, T.G.; Popov, V.; Gorchakov, R.; et al. Negevirus: A proposed new taxon of insect-specific viruses with wide geographic distribution. J. Virol. 2013, 87, 2475–2488. [Google Scholar] [CrossRef]
- Nunes, M.R.T.; Contreras-Gutierrez, M.A.; Guzman, H.; Martins, L.C.; Barbirato, M.F.; Savit, C.; Balta, V.; Uribe, S.; Vivero, R.; Suaza, J.D.; et al. Genetic characterization, molecular epidemiology, and phylogenetic relationships of insect-specific viruses in the taxon negevirus. Virology 2017, 504, 152–167. [Google Scholar] [CrossRef]
- Presti, R.M.; Zhao, G.; Beatty, W.L.; Mihindukulasuriya, K.A.; da Rosa, A.P.; Popov, V.L.; Tesh, R.B.; Virgin, H.W.; Wang, D. Quaranfil, johnston atoll, and lake chad viruses are novel members of the family orthomyxoviridae. J. Virol. 2009, 83, 11599–11606. [Google Scholar] [CrossRef]
- Da Silva, E.V.; Da Rosa, A.P.; Nunes, M.R.; Diniz, J.A.; Tesh, R.B.; Cruz, A.C.; Vieira, C.M.; Vasconcelos, P.F. Araguari virus, a new member of the family orthomyxoviridae: Serologic, ultrastructural, and molecular characterization. Am. J. Trop. Med. Hyg. 2005, 73, 1050–1058. [Google Scholar] [CrossRef]
- Bussetti, A.V.; Palacios, G.; Travassos da Rosa, A.; Savji, N.; Jain, K.; Guzman, H.; Hutchison, S.; Popov, V.L.; Tesh, R.B.; Lipkin, W.I. Genomic and antigenic characterization of jos virus. J. Gen. Virol. 2012, 93, 293–298. [Google Scholar] [CrossRef]
- Kosoy, O.I.; Lambert, A.J.; Hawkinson, D.J.; Pastula, D.M.; Goldsmith, C.S.; Hunt, D.C.; Staples, J.E. Novel thogotovirus associated with febrile illness and death, united states, 2014. Emerg. Infect. Dis. 2015, 21, 760–764. [Google Scholar] [CrossRef]
- Allison, A.B.; Ballard, J.R.; Tesh, R.B.; Brown, J.D.; Ruder, M.G.; Keel, M.K.; Munk, B.A.; Mickley, R.M.; Gibbs, S.E.; Travassos da Rosa, A.P.; et al. Cyclic avian mass mortality in the northeastern united states is associated with a novel orthomyxovirus. J. Virol. 2015, 89, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Lvov, D.K.; Al’khovskii, S.V.; Shchelkanov, M.; Shchetinin, A.M.; Deriabin, P.G.; Aristova, V.A.; Gitel’man, A.K.; Samokhvalov, E.I.; Botikov, A.G. Taxonomic status of the tyulek virus (tlkv) (orthomyxoviridae, quaranjavirus, quaranfil group) isolated from the ticks argas vulgaris filippova, 1961 (argasidae) from the birds burrow nest biotopes in the kyrgyzstan. Vopr. Virusol. 2014, 59, 28–32. [Google Scholar]
- Clerx, J.P.; Fuller, F.; Bishop, D.H. Tick-borne viruses structurally similar to orthomyxoviruses. Virology 1983, 127, 205–219. [Google Scholar] [CrossRef]
- Zeller, H.G.; Karabatsos, N.; Calisher, C.H.; Digoutte, J.P.; Murphy, F.A.; Shope, R.E. Electron microscopy and antigenic studies of uncharacterized viruses. I. Evidence suggesting the placement of viruses in families arenaviridae, paramyxoviridae, or poxviridae. Arch. Virol. 1989, 108, 191–209. [Google Scholar] [CrossRef] [PubMed]
- Overby, A.K.; Popov, V.; Neve, E.P.; Pettersson, R.F. Generation and analysis of infectious virus-like particles of uukuniemi virus (bunyaviridae): A useful system for studying bunyaviral packaging and budding. J. Virol. 2006, 80, 10428–10435. [Google Scholar] [CrossRef]
- Westaway, E.G.; Mackenzie, J.M.; Kenney, M.T.; Jones, M.K.; Khromykh, A.A. Ultrastructure of kunjin virus-infected cells: Colocalization of ns1 and ns3 with double-stranded rna, and of ns2b with ns3, in virus-induced membrane structures. J. Virol. 1997, 71, 6650–6661. [Google Scholar]
- Mackenzie, J. Wrapping things up about virus RNA replication. Traffic 2005, 6, 967–977. [Google Scholar] [CrossRef]
- Miller, S.; Krijnse-Locker, J. Modification of intracellular membrane structures for virus replication. Nat. Rev. Microbiol. 2008, 6, 363–374. [Google Scholar] [CrossRef]
- Attoui, H.; Mohd Jaafar, F.; Biagini, P.; Cantaloube, J.F.; de Micco, P.; Murphy, F.A.; de Lamballerie, X. Genus coltivirus (family reoviridae): Genomic and morphologic characterization of old world and new world viruses. Arch. Virol. 2002, 147, 533–561. [Google Scholar] [CrossRef]
- Frolova, E.I.; Gorchakov, R.; Pereboeva, L.; Atasheva, S.; Frolov, I. Functional sindbis virus replicative complexes are formed at the plasma membrane. J. Virol. 2010, 84, 11679–11695. [Google Scholar] [CrossRef] [PubMed]
- Froshauer, S.; Kartenbeck, J.; Helenius, A. Alphavirus RNA replicase is located on the cytoplasmic surface of endosomes and lysosomes. J. Cell Biol. 1988, 107, 2075–2086. [Google Scholar] [CrossRef] [PubMed]
- Boehme, K.W.; Popov, V.L.; Heidner, H.W. The host range phenotype displayed by a sindbis virus glycoprotein variant results from virion aggregation and retention on the surface of mosquito cells. J. Virol. 2000, 74, 11398–11406. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Grimley, P.M.; Berezesky, I.K.; Friedman, R.M. Cytoplasmic structures associated with an arbovirus infection: Loci of viral ribonucleic acid synthesis. J. Virol. 1968, 2, 1326–1338. [Google Scholar] [PubMed]
- Nasar, F.; Gorchakov, R.V.; Tesh, R.B.; Weaver, S.C. Eilat virus host range restriction is present at multiple levels of the virus life cycle. J. Virol. 2015, 89, 1404–1418. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Seymour, R.L.; Kaelber, J.T.; Kim, D.Y.; Leal, G.; Sherman, M.B.; Frolov, I.; Chiu, W.; Weaver, S.C.; Nasar, F. Novel insect-specific eilat virus-based chimeric vaccine candidates provide durable, mono- and multivalent, single-dose protection against lethal alphavirus challenge. J. Virol. 2018, 92. [Google Scholar]
- Erasmus, J.H.; Auguste, A.J.; Kaelber, J.T.; Luo, H.; Rossi, S.L.; Fenton, K.; Leal, G.; Kim, D.Y.; Chiu, W.; Wang, T.; et al. A chikungunya fever vaccine utilizing an insect-specific virus platform. Nat. Med. 2017, 23, 192–199. [Google Scholar] [CrossRef]
- Zirkel, F.; Kurth, A.; Quan, P.L.; Briese, T.; Ellerbrok, H.; Pauli, G.; Leendertz, F.H.; Lipkin, W.I.; Ziebuhr, J.; Drosten, C.; et al. An insect nidovirus emerging from a primary tropical rainforest. MBio 2011, 2, e00077-11. [Google Scholar] [CrossRef]
- Zirkel, F.; Roth, H.; Kurth, A.; Drosten, C.; Ziebuhr, J.; Junglen, S. Identification and characterization of genetically divergent members of the newly established family mesoniviridae. J. Virol. 2013, 87, e6346–e6358. [Google Scholar] [CrossRef]
- Kallies, R.; Kopp, A.; Zirkel, F.; Estrada, A.; Gillespie, T.R.; Drosten, C.; Junglen, S. Genetic characterization of goutanap virus, a novel virus related to negeviruses, cileviruses and higreviruses. Viruses 2014, 6, 4346–4357. [Google Scholar] [CrossRef] [PubMed]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popov, V.L.; Tesh, R.B.; Weaver, S.C.; Vasilakis, N. Electron Microscopy in Discovery of Novel and Emerging Viruses from the Collection of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA). Viruses 2019, 11, 477. https://doi.org/10.3390/v11050477
Popov VL, Tesh RB, Weaver SC, Vasilakis N. Electron Microscopy in Discovery of Novel and Emerging Viruses from the Collection of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA). Viruses. 2019; 11(5):477. https://doi.org/10.3390/v11050477
Chicago/Turabian StylePopov, Vsevolod L., Robert B. Tesh, Scott C. Weaver, and Nikos Vasilakis. 2019. "Electron Microscopy in Discovery of Novel and Emerging Viruses from the Collection of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA)" Viruses 11, no. 5: 477. https://doi.org/10.3390/v11050477
APA StylePopov, V. L., Tesh, R. B., Weaver, S. C., & Vasilakis, N. (2019). Electron Microscopy in Discovery of Novel and Emerging Viruses from the Collection of the World Reference Center for Emerging Viruses and Arboviruses (WRCEVA). Viruses, 11(5), 477. https://doi.org/10.3390/v11050477