Prophylactic and Therapeutic Vaccination against Hepatitis C Virus (HCV): Developments and Future Perspectives

Abstract
:1. Introduction
2. Genetic Diversity of HCV
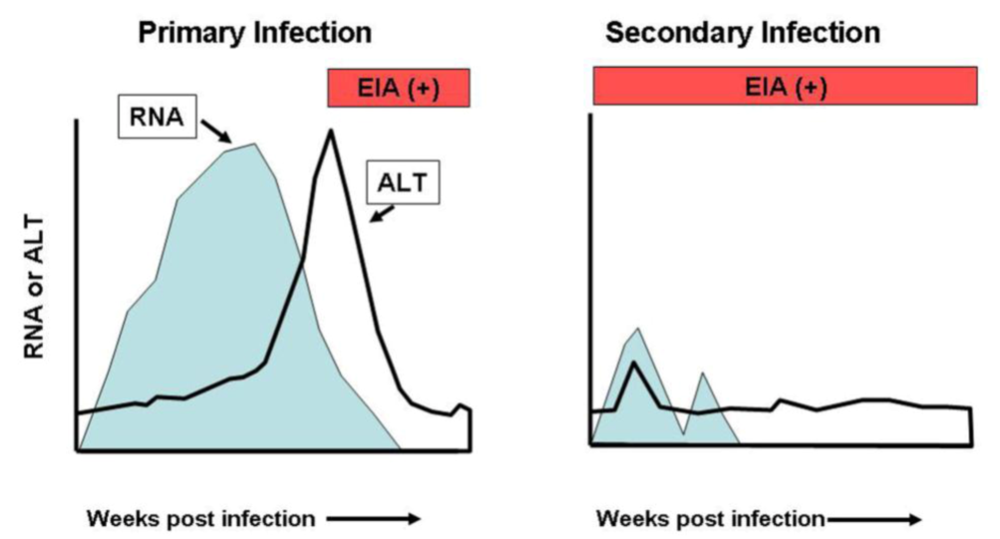
3. HCV Kinetics During Primary and Secondary Infections
4. Immune Correlates of Viral Clearance
5. Prophylactic Vaccines

{kind=link}
| Vaccine(Genotype sequence used)(Number of animals vaccinated) | Immunogenicity | Challenge Inoculum*Dose (CID50§) | Outcome | Ref. |
|---|---|---|---|---|
| Recombinant gpE1/gpE2 in Oil/water adjuvants(Genotype 1a)(N=21) | Induced antibodies to E1E2 | Genotype 1aHomologous10 to 100CID50 | Protects against infection or chronic infection5 protected from infection14 resolved infection2 developed persistent infection | [72,73] |
| DNA vaccine expressing E2 protein(Genotype 1a)(N=2) | Induced antibody and T cell responses to E2. | Genotype 1aHomologous100CID50 | Modifies infection, protects from chronic infection2 resolved infection | [74] |
| Recombinant gpE1/gpE2 in oil/water adjuvants(Genotype 1a)(N=1) | Induced antibodies and cellular responses to E1E2 | Genotype 1aHomologous100CID50 | Delayed/modified infection1 developed persistent infection | [76] |
| DNA prime and protein boost (using C, gpE1, gpE2 and NS3)(Genotypes 1a (core, NS3) and 1b (core, E1E2, NS3))(N=2) | Induced specific T-cell responses and antibody to E1 and E2. | Genotype 1bHomologous25CID50 | Modifies infection, protects from chronic infection1 resolved infection1 developed persistent infection | [77] |
| DNA prime and Recombinant Adenovirus expressing core, E1,E2 and NS3 to NS5B(Genotype 1b)(N=6) | Induced specific T-cell responses and anti-E2 antibody (neutralizing) | Genotype 1bHomologous100CID50 | Modifies infection, protects from chronic infection1 protected from infection1 resolved infection4 developed persistent infection | [78] |
| DNA prime and Recombinant VV expressing NS3,NS5A,NS5B(Genotype 1a)(N=1) | Induced specific T-cell responses | Genotype 1aHomologous100CID50 | Modifies infection1 developed persistent infection | [25] |
| DNA prime and Recombinant Adenovirus expressing NS3 to NS5B(Genotype 1b)(N=5) | Induced specific T-cell responses | Genotype 1aHeterologous100CID50 | Modifies infection, protects from chronic infection4 resolved infection1 developed persistent infection | [26] |
| DNA prime and rMVA boost (using C, gpE1, gpE2 and NS3)(Genotype 1b)(N=4) | Induced specific T-cell responses and antibody to E1 and E2. | Genotype 1bHomologous25CID50 | Modifies infection, protects from chronic infection1 resolved infection3 developed persistent infection | [79] |
| Recombinant VLPs containing C, E1 and E2(Genotype 1b)(N=4) | Induced specific T-cell responses. No detectable anti-E1E2 response. | Genotype 1bHomologous100CID50 | Modifies infection, protects from chronic infection4 resolved infection | [75] |
| Recombinant VV core, E1, E2, p7, NS2 and NS3(Genotype 1b)(N=4) | Induced specific T-cell responses. Weak anti-E1E2 response. | Genotype 1bHomologous2.5 and 24CID50† | Modifies infection, protects from chronic infection4 resolved infection | [70] |
| DNA prime and Recombinant Adenovirus expressing NS3,NS5A,NS5B(Genotype 1a)(N=2) | Induced specific T-cell responses | Genotype 1aHomologous100CID50 | Modifies infection or protects from chronic infection1 resolved infection1 developed persistent infection | [71] |
| Vaccine(Genotype sequence used) | Patient HCV Genotype | Immunogenicity | Outcome | Ref. |
|---|---|---|---|---|
| Alum-adjuvanted recombinant gpE1 protein(Genotype 1b) | *Genotype 1a Genotype 1b | Antibody and T-cell responses in some healthy volunteers.Boosts humoral and cellular responses in a subset of patients | May ameliorate hepatitis | [109,115,116] |
| Peptide vaccine targeting E1, E2, NS3 and NS5A.(Genotype 1b) | Genotype 1b | Boosted peptide-specific T-cell and antibody responses. | Decreased ALT levels and HCV RNA levels in a subset of patients. | [106] |
| DNA vaccine (CIGB-230) expressing core, E1, E2 plus recombinant core protein(Genotype 1b) | Genotype 1b | Boosted neutralizing antibody responses and T-cell responses in patients. | May ameliorate hepatitis | [110] |
| Synthetic peptide vaccine (core, NS3, NS4) IC41 with or without poly-L-arginine.Genotype 1 | Genotype 1Genotype 3Genotype 4 | Boosted T-cell responses in patients. | Transient decline (>1-log) in HCV serum RNA titers in a subset of patients. | [107] |
| V-5 Immunitor – heat-inactivated HCV antigens from HCV-infected donors administered orally in tablet form. (Genotype not defined) | Genotype not defined | Not measured. | Decreased liver enzyme levels. May improve HCV-associated clinical symptoms. | [111] |
| ICOMATRIX-adjuvanted recombinant core protein(Genotype 1a) | Healthy patients used. | Induced HCV-specific immune responses and CD+ T-cell responses in a subset of patients. | Clinical trial in healthy adults, no HCV infected patients tested. | [117,118,119] |
| MVA expressing NS3, NS4 and NS5B proteins (Transgene TG4040). (Genotype 1b) | Genotype not defined | Boosted HCV-specific immune responses | Transient decrease in viral load in a subset of patients. | [108,120,121] |
6. Therapeutic Vaccines
7. Conclusions
Acknowledgments
References
- Bartenschlager, R.; Frese, M.; Pietschmann, T. Novel insights into hepatitis C virus replication and persistence. Adv. Virus Res. 2004, 63, 71–180. [Google Scholar] [PubMed]
- Moradpour, D.; Penin, F.; Rice, C.M. Replication of hepatitis C virus. Nat. Rev. Microbiol. 2007, 5, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Alter, M.J.; Kruszon-Moran, D.; Nainan, O.V.; McQuillan, G.M.; Gao, F.; Moyer, L.A.; Kaslow, R.A.; Margolis, H.S. The prevalence of hepatitis C virus infection in the United States, 1988 through 1994. N. Engl. J. Med. 1999, 341, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.J.; Rehermann, B.; Seeff, L.B.; Hoofnagle, J.H. Pathogenesis, natural history, treatment, and prevention of hepatitis C. Ann. Intern. Med. 2000, 132, 296–305. [Google Scholar] [PubMed]
- Hoofnagle, J.H. Course and outcome of hepatitis C. Hepatology 2002, 36, S21–S29. [Google Scholar] [CrossRef] [PubMed]
- Manns, M.P.; McHutchison, J.G.; Gordon, S.C.; Rustgi, V.K.; Shiffman, M.; Reindollar, R.; Goodman, Z.D.; Koury, K.; Ling, M.; Albrecht, J.K. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: a randomised trial. Lancet 2001, 358, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Fried, M.W.; Shiffman, M.L.; Reddy, K.R.; Smith, C.; Marinos, G.; Goncales, F.L.; Haussinger, D.; Diago, M.; Carosi, G.; Dhumeaux, D.; Craxi, A.; Lin, A.; Hoffman, J.; Yu, J. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N. Engl. J. Med. 2002, 347, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, P.; Bukh, J.; Combet, C.; Deleage, G.; Enomoto, N.; Feinstone, S.; Halfon, P.; Inchauspe, G.; Kuiken, C.; Maertens, G.; Mizokami, M.; Murphy, D.G.; Okamoto, H.; Pawlotsky, J.M.; Penin, F.; Sablon, E.; Shin, I.; Stuyver, L.J.; Thiel, H.J.; Viazov, S.; Weiner, A.J.; Widell, A. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology 2005, 42, 962–973. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, H.; Hijikata, M.; Tanji, Y.; Kimura, K.; Shimotohno, K. Analysis of N-terminal processing of hepatitis C virus nonstructural protein 2. J. Virol. 1994, 68, 2731–2734. [Google Scholar] [PubMed]
- Weiner, A.J.; Brauer, M.J.; Rosenblatt, J.; Richman, K.H.; Tung, J.; Crawford, K.; Bonino, F.; Saracco, G.; Choo, Q.L.; Houghton, M. Variable and hypervariable domains are found in the regions of HCV corresponding to the flavivirus envelope and NS1 proteins and the pestivirus envelope glycoproteins. Virology 1991, 180, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, H.; Kato, N.; Ootsuyama, Y.; Nakazawa, T.; Yamauchi, K.; Shimotohno, K. Genetic alterations of the putative envelope proteins encoding region of the hepatitis C virus in the progression to relapsed phase from acute hepatitis: humoral immune response to hypervariable region 1. Int. J. Cancer 1994, 57, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Major, M.E.; Feinstone, S.M. The molecular virology of hepatitis C. Hepatology 1997, 25, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, S.; Okamoto, H.; Sakamoto, M.; Kojima, M.; Tsuda, F.; Tanaka, T.; Munekata, E.; Muchmore, E.E.; Peterson, D.A.; Mishiro, S. A structurally flexible and antigenically variable N-terminal domain of the hepatitis C virus E2/NS1 protein: implication for an escape from antibody. Virology 1993, 195, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Forns, X.; Thimme, R.; Govindarajan, S.; Emerson, S.U.; Purcell, R.H.; Chisari, F.V.; Bukh, J. Hepatitis C virus lacking the hypervariable region 1 of the second envelope protein is infectious and causes acute resolving or persistent infection in chimpanzees. Proc. Natl. Acad. Sci. USA 2000, 97, 13318–13323. [Google Scholar] [CrossRef]
- Farci, P.; Shimoda, A.; Wong, D.; Cabezon, T.; De Gioannis, D.; Strazzera, A.; Shimizu, Y.; Shapiro, M.; Alter, H.J.; Purcell, R.H. Prevention of hepatitis C virus infection in chimpanzees by hyperimmune serum against the hypervariable region 1 of the envelope 2 protein. Proc. Natl. Acad. Sci. USA 1996, 93, 15394–15399. [Google Scholar] [CrossRef]
- Weiner, A.J.; Geysen, H.M.; Christopherson, C.; Hall, J.E.; Mason, T.J.; Saracco, G.; Bonino, F.; Crawford, K.; Marion, C.D.; Crawford, K.A.; Brunetto, M.; Barr, P.J.; Miyamura, T.; McHutchinson, J.; Houghton, M. Evidence for immune selection of hepatitis C virus (HCV) putative envelope glycoprotein variants: Potential role in chronic HCV infections. Proc. Natl. Acad. Sci. USA 1992, 89, 3468–3472. [Google Scholar] [CrossRef]
- Kato, N.; Sekiya, H.; Ootsuyama, Y.; Nakazawa, T.; Hijikata, M.; Ohkoshi, S.; Shimotohno, K. Humoral immune response to hypervariable region 1 of the putative envelope glycoprotein (gp70) of hepatitis C virus. J. Virol. 1993, 67, 3923–3930. [Google Scholar] [PubMed]
- Shimizu, Y.K.; Hijikata, M.; Iwamoto, A.; Alter, H.J.; Purcell, R.H.; Yoshikura, H. Neutralizing antibodies against hepatitis C virus and the emergence of neutralization escape mutant viruses. J. Virol. 1994, 68, 1494–1500. [Google Scholar] [PubMed]
- Stumpf, M.P.; Pybus, O.G. Genetic diversity and models of viral evolution for the hepatitis C virus. FEMS Microbiol. Lett. 2002, 214, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.U.; Lam, N.P.; Dahari, H.; Gretch, D.R.; Wiley, T.E.; Layden, T.J.; Perelson, A.S. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-α therapy. Science 1998, 282, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Erickson, A.L.; Kimura, Y.; Igarashi, S.; Eichelberger, J.; Houghton, M.; Sidney, J.; McKinney, D.; Sette, A.; Hughes, A.L.; Walker, C.M. The outcome of hepatitis C virus infection is predicted by escape mutations in epitopes targeted by cytotoxic T lymphocytes. Immunity 2001, 15, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Bowen, D.G.; Walker, C.M. Mutational escape from CD8+ T cell immunity: HCV evolution, from chimpanzees to man. J. Exp. Med. 2005, 201, 1709–1714. [Google Scholar] [CrossRef] [PubMed]
- Uebelhoer, L.; Han, J.H.; Callendret, B.; Mateu, G.; Shoukry, N.H.; Hanson, H.L.; Rice, C.M.; Walker, C.M.; Grakoui, A. Stable cytotoxic T cell escape mutation in hepatitis C virus is linked to maintenance of viral fitness. PLoS. Pathog. 2008, 4, 1–15. [Google Scholar] [CrossRef]
- von Hahn, T.; Yoon, J.C.; Alter, H.; Rice, C.M.; Rehermann, B.; Balfe, P.; McKeating, J.A. Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection in vivo. Gastroenterology 2007, 132, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.; Mihalik, K.; Tilton, J.C.; Williams, O.; Merchlinsky, M.; Connors, M.; Feinstone, S.M.; Major, M.E. CD4+ immune escape and subsequent T-cell failure following chimpanzee immunization against hepatitis C virus. Hepatology 2006, 44, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Folgori, A.; Capone, S.; Ruggeri, L.; Meola, A.; Sporeno, E.; Ercole, B.B.; Pezzanera, M.; Tafi, R.; Arcuri, M.; Fattori, E.; Lahm, A.; Luzzago, A.; Vitelli, A.; Colloca, S.; Cortese, R.; Nicosia, A. A T-cell HCV vaccine eliciting effective immunity against heterologous virus challenge in chimpanzees. Nat. Med. 2006, 12, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Logvinoff, C.; Major, M.E.; Oldach, D.; Heyward, S.; Talal, A.; Balfe, P.; Feinstone, S.M.; Alter, H.; Rice, C.M.; McKeating, J.A. Neutralizing antibody response during acute and chronic hepatitis C virus infection. Proc. Natl. Acad. Sci. USA 2004, 101, 10149–10154. [Google Scholar] [CrossRef]
- Owsianka, A.; Tarr, A.W.; Juttla, V.S.; Lavillette, D.; Bartosch, B.; Cosset, F.L.; Ball, J.K.; Patel, A.H. Monoclonal antibody AP33 defines a broadly neutralizing epitope on the hepatitis C virus E2 envelope glycoprotein. J. Virol. 2005, 79, 11095–11104. [Google Scholar] [CrossRef] [PubMed]
- Tarr, A.W.; Owsianka, A.M.; Timms, J.M.; McClure, C.P.; Brown, R.J.; Hickling, T.P.; Pietschmann, T.; Bartenschlager, R.; Patel, A.H.; Ball, J.K. Characterization of the hepatitis C virus E2 epitope defined by the broadly neutralizing monoclonal antibody AP33. Hepatology 2006, 43, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Law, M.; Maruyama, T.; Lewis, J.; Giang, E.; Tarr, A.W.; Stamataki, Z.; Gastaminza, P.; Chisari, F.V.; Jones, I.M.; Fox, R.I.; Ball, J.K.; McKeating, J.A.; Kneteman, N.M.; Burton, D.R. Broadly neutralizing antibodies protect against hepatitis C virus quasispecies challenge. Nat. Med. 2008, 14, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Dahari, H.; Major, M.; Zhang, X.; Mihalik, K.; Rice, C.M.; Perelson, A.S.; Feinstone, S.M.; Neumann, A.U. Mathematical modeling of primary hepatitis C infection: noncytolytic clearance and early blockage of virion production. Gastroenterology 2005, 128, 1056–1066. [Google Scholar] [CrossRef] [PubMed]
- Major, M.E.; Dahari, H.; Mihalik, K.; Puig, M.; Rice, C.M.; Neumann, A.U.; Feinstone, S.M. Hepatitis C virus kinetics and host responses associated with disease and outcome of infection in chimpanzees. Hepatology 2004, 39, 1709–1720. [Google Scholar] [CrossRef] [PubMed]
- Prince, A.M.; Pawlotsky, J.M.; Soulier, A.; Tobler, L.; Brotman, B.; Pfahler, W.; Lee, D.H.; Li, L.; Shata, M.T. Hepatitis C virus replication kinetics in chimpanzees with self-limited and chronic infections. J. Viral Hepat. 2004, 11, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Shoukry, N.H.; Grakoui, A.; Houghton, M.; Chien, D.Y.; Ghrayeb, J.; Reimann, K.A.; Walker, C.M. Memory CD8+ T cells are required for protection from persistent hepatitis C virus infection. J. Exp. Med. 2003, 197, 1645–1655. [Google Scholar] [CrossRef] [PubMed]
- Thimme, R.; Bukh, J.; Spangenberg, H.C.; Wieland, S.; Pemberton, J.; Steiger, C.; Govindarajan, S.; Purcell, R.H.; Chisari, F.V. Viral and immunological determinants of hepatitis C virus clearance, persistence, and disease. Proc. Natl. Acad. Sci. USA 2002, 99, 15661–15668. [Google Scholar] [CrossRef]
- Farci, P.; Alter, H.J.; Govindarajan, S.; Wong, D.C.; Engle, R.; Lesniewski, R.; Mushahwar, I.K.; Desai, S.M.; Miller, R.H.; Ogata, N.; Purcell, R.H. Lack of protective immunity against reinfection with hepatitis C virus. Science 1992, 258, 140. [Google Scholar]
- Prince, A.M.; Brotman, B.; Huima, T.; Pascual, D.; Jaffery, M.; Inchauspe, G. Immunity in hepatitis C infection. J. Infect. Dis. 1992, 165, 438–443. [Google Scholar] [PubMed]
- Bassett, S.E.; Guerra, B.; Brasky, K.; Miskovsky, E.; Houghton, M.; Klimpel, G.R.; Lanford, R.E. Protective immune response to hepatitis C virus in chimpanzees rechallenged following clearance of primary infection. Hepatology 2001, 33, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Major, M.E.; Mihalik, K.; Puig, M.; Rehermann, B.; Nascimbeni, M.; Rice, C.M.; Feinstone, S.M. Previously infected and recovered chimpanzees exhibit rapid responses that control hepatitis C virus replication upon rechallenge. J. Virol. 2002, 76, 6586–6595. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.H.; Cox, A.; Hoover, D.R.; Wang, X.H.; Mao, Q.; Ray, S.; Strathdee, S.A.; Vlahov, D.; Thomas, D.L. Protection against persistence of hepatitis C. Lancet 2002, 359, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Bukh, J.; Thimme, R.; Meunier, J.C.; Faulk, K.; Spangenberg, H.C.; Chang, K.M.; Satterfield, W.; Chisari, F.V.; Purcell, R.H. Previously infected chimpanzees are not consistently protected against reinfection or persistent infection after reexposure to the identical hepatitis C virus strain. J. Virol. 2008, 82, 8183–8195. [Google Scholar] [CrossRef] [PubMed]
- Lanford, R.E.; Guerra, B.; Chavez, D.; Bigger, C.; Brasky, K.M.; Wang, X.H.; Ray, S.C.; Thomas, D.L. Cross-genotype immunity to hepatitis C virus. J. Virol. 2004, 78, 1575–1581. [Google Scholar] [CrossRef] [PubMed]
- Weiner, A.J.; Paliard, X.; Selby, M.J.; Medina-Selby, A.; Coit, D.; Nguyen, S.; Kansopon, J.; Arian, C.L.; Ng, P.; Tucker, J.; Lee, C.T.; Polakos, N.K.; Han, J.; Wong, S.; Lu, H.H.; Rosenberg, S.; Brasky, K.M.; Chien, D.; Kuo, G.; Houghton, M. Intrahepatic genetic inoculation of hepatitis C virus RNA confers cross-protective immunity. J. Virol. 2001, 75, 7142–7148. [Google Scholar] [CrossRef] [PubMed]
- Diepolder, H.M.; Zachoval, R.; Hoffmann, R.M.; Wierenga, E.A.; Santantonio, T.; Jung, M.C.; Eichenlaub, D.; Pape, G.R. Possible mechanism involving T-lymphocyte response to non-structural protein 3 in viral clearance in acute hepatitis C virus infection. Lancet 1995, 346, 1006–1007. [Google Scholar] [CrossRef] [PubMed]
- Diepolder, H.M.; Gerlach, J.T.; Zachoval, R.; Hoffmann, R.M.; Jung, M.C.; Wierenga, E.A.; Scholtz, S.; Santantonio, T.; Houghton, M.; Southwood, S.; Sette, A.; Pape, G.R. Immunodominant CD4+ T-cell epitope within nonstructural protein 3 in acute hepatitis C virus infection. J. Virol. 1997, 71, 6011–6019. [Google Scholar] [PubMed]
- Gerlach, J.T.; Diepolder, H.M.; Jung, M.C.; Gruener, N.H.; Schraut, W.W.; Zachoval, R.; Hoffmann, R.; Schirren, C.A.; Santantonio, T.; Pape, G.R. Recurrence of hepatitis C virus after loss of virus-specific CD4(+) T- cell response in acute hepatitis C. Gastroenterology 1999, 117, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, A.M.; Miner, C.; Lewinsohn, D.M.; Sasaki, A.W.; Kaufman, E.; Rosen, H.R. Novel CD4+ and CD8+ T-cell determinants within the NS3 protein in subjects with spontaneously resolved HCV infection. Hepatology 2003, 37, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Lechner, F.; Wong, D.K.; Dunbar, P.R.; Chapman, R.; Chung, R.T.; Dohrenwend, P.; Robbins, G.; Phillips, R.; Klenerman, P.; Walker, B.D. Analysis of successful immune responses in persons infected with hepatitis C virus. J. Exp. Med. 2000, 191, 1499–1512. [Google Scholar] [CrossRef] [PubMed]
- Thimme, R.; Oldach, D.; Chang, K.M.; Steiger, C.; Ray, S.C.; Chisari, F.V. Determinants of viral clearance and persistence during acute hepatitis C virus infection. J. Exp. Med. 2001, 194, 1395–1406. [Google Scholar] [CrossRef] [PubMed]
- Bowen, D.G.; Walker, C.M. Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature 2005, 436, 946–952. [Google Scholar] [CrossRef] [PubMed]
- Takaki, A.; Wiese, M.; Maertens, G.; Depla, E.; Seifert, U.; Liebetrau, A.; Miller, J.L.; Manns, M.P.; Rehermann, B. Cellular immune responses persist and humoral responses decrease two decades after recovery from a single-source outbreak of hepatitis C. Nat. Med. 2000, 6, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Grakoui, A.; Shoukry, N.H.; Woollard, D.J.; Han, J.H.; Hanson, H.L.; Ghrayeb, J.; Murthy, K.K.; Rice, C.M.; Walker, C.M. HCV persistence and immune evasion in the absence of memory T cell help. Science 2003, 302, 659–662. [Google Scholar] [CrossRef] [PubMed]
- Nascimbeni, M.; Mizukoshi, E.; Bosmann, M.; Major, M.E.; Mihalik, K.; Rice, C.M.; Feinstone, S.M.; Rehermann, B. Kinetics of CD4+ and CD8+ memory T-cell responses during hepatitis C virus rechallenge of previously recovered chimpanzees. J. Virol. 2003, 77, 4781–4793. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, B.; Dubuisson, J.; Cosset, F.L. Infectious hepatitis C virus pseudo-particles containing functional E1-E2 envelope protein complexes. J. Exp. Med. 2003, 197, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.; Zhang, J.; Flint, M.; Logvinoff, C.; Cheng-Mayer, C.; Rice, C.M.; McKeating, J.A. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc. Natl. Acad. Sci. USA 2003, 100, 7271–7276. [Google Scholar] [CrossRef]
- Wakita, T.; Pietschmann, T.; Kato, T.; Date, T.; Miyamoto, M.; Zhao, Z.; Murthy, K.; Habermann, A.; Krausslich, H.G.; Mizokami, M.; Bartenschlager, R.; Liang, T.J. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat. Med. 2005, 11, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Lindenbach, B.D.; Evans, M.J.; Syder, A.J.; Wolk, B.; Tellinghuisen, T.L.; Liu, C.C.; Maruyama, T.; Hynes, R.O.; Burton, D.R.; McKeating, J.A.; Rice, C.M. Complete replication of hepatitis C virus in cell culture. Science 2005, 309, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Matsumura, T.; Heller, T.; SAITO, S.; Sapp, R.K.; Murthy, K.; Wakita, T.; Liang, T.J. Production of infectious hepatitis C virus of various genotypes in cell cultures. J. Virol. 2007, 81, 4405–4411. [Google Scholar] [CrossRef] [PubMed]
- Gottwein, J.M.; Scheel, T.K.; Hoegh, A.M.; Lademann, J.B.; Eugen-Olsen, J.; Lisby, G.; Bukh, J. Robust hepatitis C genotype 3a cell culture releasing adapted intergenotypic 3a/2a (S52/JFH1) viruses. Gastroenterology 2007, 133, 1614–1626. [Google Scholar] [CrossRef] [PubMed]
- Scheel, T.K.; Gottwein, J.M.; Jensen, T.B.; Prentoe, J.C.; Hoegh, A.M.; Alter, H.J.; Eugen-Olsen, J.; Bukh, J. Development of JFH1-based cell culture systems for hepatitis C virus genotype 4a and evidence for cross-genotype neutralization. Proc. Natl. Acad. Sci. USA 2008, 105, 997–1002. [Google Scholar] [CrossRef]
- Jensen, T.B.; Gottwein, J.M.; Scheel, T.K.; Hoegh, A.M.; Eugen-Olsen, J.; Bukh, J. Highly efficient JFH1-based cell-culture system for hepatitis C virus genotype 5a: failure of homologous neutralizing-antibody treatment to control infection. J. Infect. Dis. 2008, 198, 1756–1765. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, B.; Bukh, J.; Meunier, J.C.; Granier, C.; Engle, R.E.; Blackwelder, W.C.; Emerson, S.U.; Cosset, F.L.; Purcell, R.H. In vitro assay for neutralizing antibody to hepatitis C virus: evidence for broadly conserved neutralization epitopes. Proc. Natl. Acad. Sci. USA 2003, 100, 14199–14204. [Google Scholar] [CrossRef]
- Pestka, J.M.; Zeisel, M.B.; Blaser, E.; Schurmann, P.; Bartosch, B.; Cosset, F.L.; Patel, A.H.; Meisel, H.; Baumert, J.; Viazov, S.; Rispeter, K.; Blum, H.E.; Roggendorf, M.; Baumert, T.F. Rapid induction of virus-neutralizing antibodies and viral clearance in a single-source outbreak of hepatitis C. Proc. Natl. Acad. Sci. USA 2007, 104, 6025–6030. [Google Scholar] [CrossRef]
- Lavillette, D.; Morice, Y.; Germanidis, G.; Donot, P.; Soulier, A.; Pagkalos, E.; Sakellariou, G.; Intrator, L.; Bartosch, B.; Pawlotsky, J.M.; Cosset, F.L. Human serum facilitates hepatitis C virus infection, and neutralizing responses inversely correlate with viral replication kinetics at the acute phase of hepatitis C virus infection. J. Virol. 2005, 79, 6023–6034. [Google Scholar] [CrossRef] [PubMed]
- Dowd, K.A.; Netski, D.M.; Wang, X.H.; Cox, A.L.; Ray, S.C. Selection pressure from neutralizing antibodies drives sequence evolution during acute infection with hepatitis C virus. Gastroenterology 2009, 136, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Meunier, J.C.; Engle, R.E.; Faulk, K.; Zhao, M.; Bartosch, B.; Alter, H.; Emerson, S.U.; Cosset, F.L.; Purcell, R.H.; Bukh, J. Evidence for cross-genotype neutralization of hepatitis C virus pseudo-particles and enhancement of infectivity by apolipoprotein C1. Proc. Natl. Acad. Sci. USA 2005, 102, 4560–4565. [Google Scholar] [CrossRef]
- Farci, P.; Alter, H.J.; Wong, D.C.; Miller, R.H.; Govindarajan, S.; Engle, R.; Shapiro, M.; Purcell, R.H. Prevention of hepatitis C virus infection in chimpanzees after antibody-mediated in vitro neutralization. Proc. Natl. Acad. Sci. USA 1994, 91, 7792–7796. [Google Scholar] [CrossRef]
- Krawczynski, K.; Alter, M.J.; Tankersley, D.L.; Beach, M.; Robertson, B.H.; Lambert, S.; Kuo, G.; Spelbring, J.E.; Meeks, E.; Sinha, S.; Carson, D.A. Effect of immune globulin on the prevention of experimental hepatitis C virus infection. J. Infect. Dis. 1996, 173, 822–828. [Google Scholar] [PubMed]
- Yu, M.Y.; Bartosch, B.; Zhang, P.; Guo, Z.P.; Renzi, P.M.; Shen, L.M.; Granier, C.; Feinstone, S.M.; Cosset, F.L.; Purcell, R.H. Neutralizing antibodies to hepatitis C virus (HCV) in immune globulins derived from anti-HCV-positive plasma. Proc. Natl. Acad. Sci. USA 2004, 101, 7705–7710. [Google Scholar] [CrossRef]
- Youn, J.W.; Hu, Y.W.; Tricoche, N.; Pfahler, W.; Shata, M.T.; Dreux, M.; Cosset, F.L.; Folgori, A.; Lee, D.H.; Brotman, B.; Prince, A.M. Evidence for protection against chronic hepatitis C virus infection in chimpanzees by immunization with replicating recombinant vaccinia virus. J. Virol. 2008, 82, 10896–10905. [Google Scholar] [CrossRef] [PubMed]
- Zubkova, I.; Choi, Y.H.; Chang, E.; Pirollo, K.; Uren, T.; Watanabe, H.; Wells, F.; Kachko, A.; Krawczynski, K.; Major, M.E. T-cell vaccines that elicit effective immune responses against HCV in chimpanzees may create greater immune pressure for viral mutation. Vaccine 2009, 27, 2594–2602. [Google Scholar] [CrossRef] [PubMed]
- Choo, Q.L.; Kuo, G.; Ralston, R.; Weiner, A.; Chien, D.; Van Nest, G.; Han, J.; Berger, K.; Thudium, K.; Kuo, C. Vaccination of chimpanzees against infection by the hepatitis C virus. Proc. Natl. Acad. Sci. USA 1994, 91, 1294–1298. [Google Scholar] [CrossRef]
- Coates, S.; Choo, Q. L.; Kuo, G.; Crawford, K.; Dong, C.; Wininger, M.; Weiner, A. J.; Berger, K.; Wong, S.; Ralston, R.; Morandi, M.; Pileri, P.; Rosa, D.; Muchmore, E.; Mahony, J.; Brasky, K.M.; Purceli, R.H.; Abrignani, S.; Houghton, M. Protection of Chimpanzees against Heterologous 1a Viral Challenge using a gpE1/gpE2 Heterodimer Vaccine. In Proceedings of the 11th International Symposium on Viral Hepatitis and Liver Disease, Sydney, Australia, 6-10 April, 2003. Jilbert, A.R., Grgacic, E., Vickery, K., Burrell, C.J., Cossart,, Y.E., Eds.; Australian Center for Hepatitis Virology: Sydney, Australia, 2004. [Google Scholar]
- Forns, X.; Payette, P.J.; Ma, X.; Satterfield, W.; Eder, G.; Mushahwar, I.K.; Govindarajan, S.; Davis, H.L.; Emerson, S.U.; Purcell, R.H.; Bukh, J. Vaccination of chimpanzees with plasmid DNA encoding the hepatitis C virus (HCV) envelope E2 protein modified the infection after challenge with homologous monoclonal HCV. Hepatology 2000, 32, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Elmowalid, G.A.; Qiao, M.; Jeong, S.H.; Borg, B.B.; Baumert, T.F.; Sapp, R.K.; Hu, Z.; Murthy, K.; Liang, T.J. Immunization with hepatitis C virus-like particles results in control of hepatitis C virus infection in chimpanzees. Proc. Natl. Acad. Sci. U SA 2007, 104, 8427–8432. [Google Scholar] [CrossRef]
- Puig, M.; Major, M.E.; Mihalik, K.; Yu, M.Y.; Feinstone, S.M. Immunization of chimpanzees with an envelope protein-based vaccine enhances specific humoral and cellular immune responses that delay hepatitis C virus infection. Vaccine 2004, 22, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Rollier, C.; Depla, E.; Drexhage, J.A.; Verschoor, E.J.; Verstrepen, B.E.; Fatmi, A.; Brinster, C.; Fournillier, A.; Whelan, J.A.; Whelan, M.; Jacobs, D.; Maertens, G.; Inchauspe, G.; Heeney, J.L. Control of heterologous hepatitis C virus infection in chimpanzees is associated with the quality of vaccine-induced peripheral T-helper immune response. J. Virol. 2004, 78, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.W.; Park, S.H.; Lavillette, D.; Cosset, F.L.; Yang, S.H.; Lee, C.G.; Jin, H.T.; Kim, C.M.; Shata, M.T.; Lee, D.H.; Pfahler, W.; Prince, A.M.; Sung, Y.C. Sustained E2 antibody response correlates with reduced peak viremia after hepatitis C virus infection in the chimpanzee. Hepatology 2005, 42, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Rollier, C.S.; Paranhos-Baccala, G.; Verschoor, E.J.; Verstrepen, B.E.; Drexhage, J.A.; Fagrouch, Z.; Berland, J.L.; Komurian-Pradel, F.; Duverger, B.; Himoudi, N.; Staib, C.; Meyr, M.; Whelan, M.; Whelan, J.A.; Adams, V.C.; Larrea, E.; Riezu, J.I.; Lasarte, J.J.; Bartosch, B.; Cosset, F.L.; Spaan, W.J.; Diepolder, H.M.; Pape, G.R.; Sutter, G.; Inchauspe, G.; Heeney, J.L. Vaccine-induced early control of hepatitis C virus infection in chimpanzees fails to impact on hepatic PD-1 and chronicity. Hepatology 2007, 45, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.D.; Heath, J.; Collins, J.; Greene, T.; Antipa, L.; Rota, P.; Bellini, W.; McChesney, M. Experimental measles. II. Infection and immunity in the rhesus macaque. Virology 1997, 233, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Betts, R.F.; Treanor, J.J. Approaches to improved influenza vaccination. Vaccine 2000, 18, 1690–1695. [Google Scholar] [CrossRef] [PubMed]
- Onorato, I.M.; Modlin, J.F.; McBean, A.M.; Thoms, M.L.; Losonsky, G.A.; Bernier, R.H. Mucosal immunity induced by enhance-potency inactivated and oral polio vaccines. J. Infect. Dis. 1991, 163, 1–6. [Google Scholar] [PubMed]
- Ross, R.S.; Viazov, S.O.; Hoffmann, S.; Roggendorf, M. Performance characteristics of a transcription-mediated nucleic acid amplification assay for qualitative detection of hepatitis C virus RNA. J. Clin. Lab Anal. 2001, 15, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Werzberger, A.; Mensch, B.; Kuter, B.; Brown, L.; Lewis, J.; Sitrin, R.; Miller, W.; Shouval, D.; Wiens, B.; Calandra, G. A controlled trial of a formalin-inactivated hepatitis A vaccine in healthy children. N. Engl. J. Med. 1992, 327, 453–457. [Google Scholar] [PubMed]
- Innis, B.L.; Snitbhan, R.; Kunasol, P.; Laorakpongse, T.; Poopatanakool, W.; Kozik, C.A.; Suntayakorn, S.; Suknuntapong, T.; Safary, A.; Tang, D.B. Protection against hepatitis A by an inactivated vaccine. JAMA 1994, 271, 1328–1334. [Google Scholar] [PubMed]
- Zuckerman, A.J. Immunization against hepatitis B. Br. Med. Bull. 1990, 46, 383–398. [Google Scholar] [PubMed]
- Koutsky, L.A.; Ault, K.A.; Wheeler, C.M.; Brown, D.R.; Barr, E.; Alvarez, F.B.;Chiacchierini; Jansen, K.U. A controlled trial of a human papillomavirus type 16 vaccine . N. Engl. J. Med. 2002, 347, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Alter, H.J.; Seeff, L.B. Recovery, persistence, and sequelae in hepatitis C virus infection: a perspective on long-term outcome. Semin. Liver Dis. 2000, 20, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Seeff, L.B. Natural history of chronic hepatitis C. Hepatology 2002, 36, S35–S46. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhong, L.; Struble, E.B.; Watanabe, H.; Kachko, A.; Mihalik, K.; Virata-Theimer, M.L.; Alter, H.J.; Feinstone, S.; Major, M. Depletion of interfering antibodies in chronic hepatitis C patients and vaccinated chimpanzees reveals broad cross-genotype neutralizing activity. Proc. Natl. Acad. Sci. USA 2009, 106, 7537–7541. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, C.G.; Mihalik, K.; Virata-Theimer, M.L.; Yu, M.Y.; Alter, H.J.; Feinstone, S.M. Hepatitis C virus epitope-specific neutralizing antibodies in Igs prepared from human plasma. Proc. Natl. Acad. Sci. USA 2007, 104, 8449–8454. [Google Scholar] [CrossRef]
- Seder, R.A.; Ahmed, R. Similarities and differences in CD4+ and CD8+ effector and memory T cell generation. Nat. Immunol. 2003, 4, 835–842. [Google Scholar] [CrossRef]
- Harari, A.; Bart, P.A.; Stohr, W.; Tapia, G.; Garcia, M.; Medjitna-Rais, E.; Burnet, S.; Cellerai, C.; Erlwein, O.; Barber, T.; Moog, C.; Liljestrom, P.; Wagner, R.; Wolf, H.; Kraehenbuhl, J.P.; Esteban, M.; Heeney, J.; Frachette, M.J.; Tartaglia, J.; McCormack, S.; Babiker, A.; Weber, J.; Pantaleo, G. An HIV-1 clade C DNA prime, NYVAC boost vaccine regimen induces reliable, polyfunctional, and long-lasting T cell responses. J. Exp. Med. 2008, 205, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Thacker, E.E.; Timares, L.; Matthews, Q.L. Strategies to overcome host immunity to adenovirus vectors in vaccine development. Exp. Rev. Vaccines. 2009, 8, 761–777. [Google Scholar] [CrossRef]
- Abraham, J.D.; Himoudi, N.; Kien, F.; Berland, J.L.; Codran, A.; Bartosch, B.; Baumert, T.; Paranhos-Baccala, G.; Schuster, C.; Inchauspe, G.; Kieny, M.P. Comparative immunogenicity analysis of modified vaccinia Ankara vectors expressing native or modified forms of hepatitis C virus E1 and E2 glycoproteins. Vaccine 2004, 22, 3917–3928. [Google Scholar] [CrossRef] [PubMed]
- Earl, P.L.; Americo, J.L.; Wyatt, L.S.; Espenshade, O.; Bassler, J.; Gong, K.; Lin, S.; Peters, E.; Rhodes, L.; Spano, Y.E.; Silvera, P.M.; Moss, B. Rapid protection in a monkeypox model by a single injection of a replication-deficient vaccinia virus. Proc. Natl. Acad. Sci. USA 2008, 105, 10889–10894. [Google Scholar] [CrossRef]
- Brinster, C.; Chen, M.; Boucreux, D.; Paranhos-Baccala, G.; Liljestrom, P.; Lemmonier, F.; Inchauspe, G. Hepatitis C virus non-structural protein 3-specific cellular immune responses following single or combined immunization with DNA or recombinant Semliki Forest virus particles. J. Gen. Virol. 2002, 83, 369–381. [Google Scholar] [PubMed]
- Lin, Y.; Kwon, T.; Polo, J.; Zhu, Y.F.; Coates, S.; Crawford, K.; Dong, C.; Wininger, M.; Hall, J.; Selby, M.; Coit, D.; Medina-Selby, A.; McCoin, C.; Ng, P.; Drane, D.; Chien, D.; Han, J.; Vajdy, M.; Houghton, M. Induction of broad CD4+ and CD8+ T-cell responses and cross-neutralizing antibodies against hepatitis C virus by vaccination with Th1-adjuvanted polypeptides followed by defective alphaviral particles expressing envelope glycoproteins gpE1 and gpE2 and nonstructural proteins 3, 4, and 5. J. Virol. 2008, 82, 7492–7503. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Levis, R.; Shen, P.; Schlesinger, S.; Rice, C.M.; Huang, H.V. Sindbis virus: an efficient, broad host range vector for gene expression in animal cells. Science 1989, 243, 1188–1191. [Google Scholar] [PubMed]
- Leitner, W.W.; Hwang, L.N.; deVeer, M.J.; Zhou, A.; Silverman, R.H.; Williams, B.R.; Dubensky, T.W.; Ying, H.; Restifo, N.P. Alphavirus-based DNA vaccine breaks immunological tolerance by activating innate antiviral pathways. Nat. Med. 2003, 9, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Perri, S.; Greer, C.E.; Thudium, K.; Doe, B.; Legg, H.; Liu, H.; Romero, R.E.; Tang, Z.; Bin, Q.; Dubensky, T.W.; Vajdy, M.; Otten, G.R.; Polo, J.M. An alphavirus replicon particle chimera derived from venezuelan equine encephalitis and sindbis viruses is a potent gene-based vaccine delivery vector. J. Virol. 2003, 77, 10394–10403. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Simon, B.; Lone, Y.C.; Chatel, L.; Barry, R.; Inchauspe, G.; Fournillier, A. A vector-based minigene vaccine approach results in strong induction of T-cell responses specific of hepatitis C virus. Vaccine 2008, 26, 2471–2481. [Google Scholar] [CrossRef] [PubMed]
- Jaeckel, E.; Cornberg, M.; Wedemeyer, H.; Santantonio, T.; Mayer, J.; Zankel, M.; Pastore, G.; Dietrich, M.; Trautwein, C.; Manns, M.P. Treatment of acute hepatitis C with interferon alfa-2b. N. Engl. J. Med. 2001, 345, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, V.; Korner, F.; Koch, J.; Herian, U.; Theilmann, L.; Bartenschlager, R. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science 1999, 285, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Blight, K.J.; Kolykhalov, A.A.; Rice, C.M. Efficient initiation of HCV RNA replication in cell culture. Science 2000, 290, 1972–1974. [Google Scholar] [CrossRef] [PubMed]
- Yutani, S.; Yamada, A.; Yoshida, K.; Takao, Y.; Tamura, M.; Komatsu, N.; Ide, T.; Tanaka, M.; Sata, M.; Itoh, K. Phase I clinical study of a personalized peptide vaccination for patients infected with hepatitis C virus (HCV) 1b who failed to respond to interferon-based therapy. Vaccine 2007, 25, 7429–7435. [Google Scholar] [CrossRef] [PubMed]
- Klade, C.S.; Wedemeyer, H.; Berg, T.; Hinrichsen, H.; Cholewinska, G.; Zeuzem, S.; Blum, H.; Buschle, M.; Jelovcan, S.; Buerger, V.; Tauber, E.; Frisch, J.; Manns, M.P. Therapeutic vaccination of chronic hepatitis C nonresponder patients with the peptide vaccine IC41. Gastroenterology 2008, 134, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Fournillier, A.; Gerossier, E.; Evlache, A.; Martin, P.; Bain, C.; Schmidt, D.; Silvestre N; Julien, M.; Habersetzer, F.; Zarski, J.; Leroy, V.; Maynard-Muet, M.; Trepo, C.; Agathon, D.; Bonnefoy, J.; Burtin, B.; Honnet, G.; Inchauspe, G. Therapeutic vaccination against HCV: pre-clinical and clinical evaluation of a novel MVA-based vaccine.5 October. In 15th International Symposium on Hepatitis C and Related Viruses: San Antonio, Texas, USA, 2008; . [Google Scholar]
- Nevens, F.; Roskams, T.; Van Vlierberghe, H.; Horsmans, Y.; Sprengers, D.; Elewaut, A.; Desmet, V.; Leroux-Roels, G.; Quinaux, E.; Depla, E.; Dincq, S.; Vander, S.C.; Maertens, G.; Hulstaert, F. A pilot study of therapeutic vaccination with envelope protein E1 in 35 patients with chronic hepatitis C. Hepatology 2003, 38, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Lajonchere, L.; Shoukry, N.H.; Gra, B.; Amador-Canizares, Y.; Helle, F.; Bedard, N.; Guerra, I.; Drouin, C.; Dubuisson, J.; Gonzalez-Horta, E.E.; Martinez, G.; Marante, J.; Cinza, Z.; Castellanos, M.; Duenas-Carrera, S. Immunogenicity of CIGB-230, a therapeutic DNA vaccine preparation, in HCV-chronically infected individuals in a Phase I clinical trial. J. Viral Hepat. 2009, 16, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Batdelger, D.; Dandii, D.; Jirathitikal, V.; Bourinbaiar, A.S. Open-label trial of therapeutic immunization with oral V-5 Immunitor (V5) vaccine in patients with chronic hepatitis C. Vaccine 2008, 26, 2733–2737. [Google Scholar] [CrossRef] [PubMed]
- Wedemeyer, H.; Schuller, E.; Schlaphoff, V.; Stauber, R.E.; Wiegand, J.; Schiefke, I.; Firbas, C.; Jilma, B.; Thursz, M.; Zeuzem, S.; Hofmann, W.P.; Hinrichsen, H.; Tauber, E.; Manns, M.P.; Klade, C.S. Therapeutic vaccine IC41 as late add-on to standard treatment in patients with chronic hepatitis C. in Press. Vaccine 2009. [Google Scholar]
- Chua, B.Y.; Eriksson, E.M.; Brown, L.E.; Zeng, W.; Gowans, E.J.; Torresi, J.; Jackson, D.C. A self-adjuvanting lipopeptide-based vaccine candidate for the treatment of hepatitis C virus infection. Vaccine 2008, 26, 4866–4875. [Google Scholar] [CrossRef] [PubMed]
- Thammanichanond, D.; Moneer, S.; Yotnda, P.; Aitken, C.; Earnest-Silveira, L.; Jackson, D.; Hellard, M.; McCluskey, J.; Torresi, J.; Bharadwaj, M. Fiber-modified recombinant adenoviral constructs encoding hepatitis C virus proteins induce potent HCV-specific T cell response. Clin. Immunol. 2008, 128, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Leroux-Roels, G.; Depla, E.; Hulstaert, F.; Tobback, L.; Dincq, S.; Desmet, J.; Desombere, I.; Maertens, G. A candidate vaccine based on the hepatitis C E1 protein: tolerability and immunogenicity in healthy volunteers. Vaccine 2004, 22, 3080–3086. [Google Scholar] [CrossRef] [PubMed]
- Leroux-Roels, G.; Batens, A.H.; Desombere, I.; Van Den, S.B.; Vander, S.C.; Maertens, G.; Hulstaert, F. Immunogenicity and tolerability of intradermal administration of an HCV E1-based vaccine candidate in healthy volunteers and patients with resolved or ongoing chronic HCV infection. Hum. Vaccin. 2005, 1, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Polakos, N.K.; Drane, D.; Cox, J.; Ng, P.; Selby, M.J.; Chien, D.; O'Hagan, D.T.; Houghton, M.; Paliard, X. Characterization of hepatitis C virus core-specific immune responses primed in rhesus macaques by a nonclassical ISCOM vaccine. J. Immunol. 2001, 166, 3589–3598. [Google Scholar] [PubMed]
- Pearse, M.J.; Drane, D. ISCOMATRIX adjuvant for antigen delivery. Adv. Drug Deliv. Rev. 2005, 57, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Drane, D.; Maraskovsky, E.; Gibson, R.; Mitchell, S.; Barnden, M.; Moskwa, A.; Shaw, D.; Gervase, B.; Coates, S.; Houghton, M.; Basser, R. Priming of CD4(+) and CD8(+) T cell responses using a HCV core ISCOMATRIX(TM) vaccine: A phase I study in healthy volunteers. Hum. Vaccin. 2009, 5, 151–157. [Google Scholar] [PubMed]
- Fournillier, A.; Gerossier, E.; Evlashev, A.; Schmitt, D.; Simon, B.; Chatel, L.; Martin, P.; Silvestre, N.; Balloul, J.M.; Barry, R.; Inchauspe, G. An accelerated vaccine schedule with a poly-antigenic hepatitis C virus MVA-based candidate vaccine induces potent, long lasting and in vivo cross-reactive T cell responses. Vaccine 2007, 25, 7339–7353. [Google Scholar] [CrossRef] [PubMed]
- Inchauspe, G.; Michel, M.L. Vaccines and immunotherapies against hepatitis B and hepatitis C viruses. J. Viral Hepat. 2007, 14, 97–103. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/)
Share and Cite
Major, M.E. Prophylactic and Therapeutic Vaccination against Hepatitis C Virus (HCV): Developments and Future Perspectives. Viruses 2009, 1, 144-165. https://doi.org/10.3390/v1020144
Major ME. Prophylactic and Therapeutic Vaccination against Hepatitis C Virus (HCV): Developments and Future Perspectives. Viruses. 2009; 1(2):144-165. https://doi.org/10.3390/v1020144
Chicago/Turabian StyleMajor, Marian E. 2009. "Prophylactic and Therapeutic Vaccination against Hepatitis C Virus (HCV): Developments and Future Perspectives" Viruses 1, no. 2: 144-165. https://doi.org/10.3390/v1020144
APA StyleMajor, M. E. (2009). Prophylactic and Therapeutic Vaccination against Hepatitis C Virus (HCV): Developments and Future Perspectives. Viruses, 1(2), 144-165. https://doi.org/10.3390/v1020144