1. Introduction
Forest fragmentation, the process through which previously intact forests are broken apart into smaller and more-isolated fragments [
1] is considered a major conservation issue worldwide [
2,
3,
4]. While the effects of forest fragmentation can positively or negatively affect different species [
1], there is widespread agreement that fragmentation of forests negatively affects biodiversity and ecosystem services, mostly through habitat loss [
1,
5]. One important consequence of habitat fragmentation is the increase in the ratio of edge to core habitat, promoting “edge effects”, which include both behavioral responses to edges by species and the change in environmental characteristics due to the transition between different habitat types [
1,
6]. Such transition creates different abiotic conditions (e.g., increased light, wind, and temperature; reduced moisture) [
3,
5] that, together with changes in landscape connectivity and behavioral responses in wildlife, influence species composition [
2,
7].
Boreal forests are no exception to the worldwide trend of forest fragmentation, being subject to anthropogenic disturbances associated with resource extraction activities [
8]. Although boreal forests are depauperate in biodiversity compared to other forest biomes [
7], these forests represent one third of the worldwide forest cover and carbon stocks [
9]. They also sustain a number of charismatic and/or threatened species, such as woodland caribou (
Rangifer tarandus caribou) and grizzly bear (
Ursus arctos), while providing natural resources of primary interest to human societies [
9]. Natural resource extraction of Canada’s boreal forest includes forestry and energy extraction, with Alberta’s forests being one of the most developed in Canada [
8]. One major source of local forest fragmentation is the extraction of bitumen from the oil sands. With the majority of bitumen being deeper than 70 m, its extraction in these areas is from the use of underground wells [
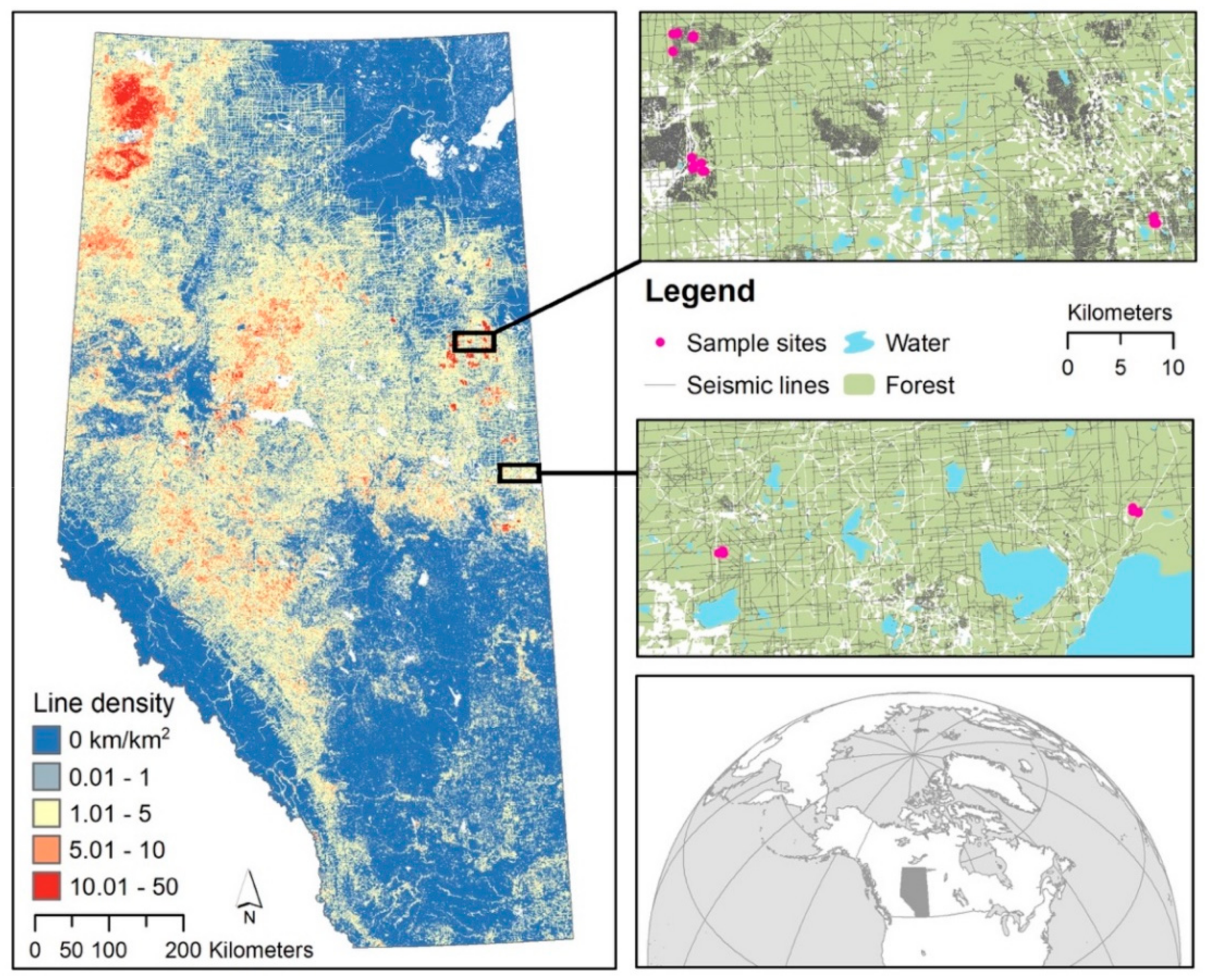
8]. Here most of the forests remain, but within that forest there are numerous smaller disturbances created by well pads, roads, pipelines, and exploratory seismic lines. Of these disturbances, seismic lines are the most pervasive in terms of extent and amount of forest edge. These narrow (<10-m wide) linear forest clearings are created to map the extent and depth of oil reserves via seismic assessments [
8,
10,
11]. Generally, there are two kinds of seismic lines: traditional (2D) and “low-impact” (3D). Traditional lines are typically 5–10 m in width, many kilometers long, and widely spaced (200–500 m), while low-impact lines (<5-m wide, typically 1.5–3 m wide) introduced since the 2000s to minimize footprint are used to more precisely determine the depth of the oil sands reserve in areas of greater interest [
12]. Although these lines are narrow, they occur at densities reaching as high as 40 km/km
2 and thus contribute most to the creation of forest edges (
Figure 1).
A key conservation issue is the need to regenerate these disturbances to forest conditions in order to minimize the effects of this footprint [
10]. For instance, for wind dispersed plant species this form of linear fragmentation increases seed dispersal [
2,
13], which is closely associated with higher wind speeds [
14,
15,
16]. As well as altering wind, these lines also alter light conditions that may affect tree regeneration and the movement of invasive species [
10,
17]. High levels of fragmentation may therefore result in a propagation of invasive species into forests. Although the relationship between corridors and fragmentation and the spread of invasive species is currently unclear [
18,
19,
20], recent experiments showed that seed dispersal is positively affected by seismic line corridors [
16], raising concern about the effects of these linear features in spreading invasive species [
21]. While propagules may make it to the site, light is a limiting factor for many plant species and thus a contributing factor to the recovery process. Seismic line disturbances change the forest structure and increase the amount of early seral forest conditions, thus altering assemblage composition and movement of vertebrate and invertebrate species [
22,
23,
24,
25,
26]. For instance, seismic lines can facilitate predator movements [
27,
28], such as increases in gray wolf (
Canis lupus) movements which increase encounter rates and potential predation of woodland caribou [
28].
Therefore, it is important to understand conditions that promote tree regeneration on seismic lines. Natural tree regeneration on many of these lines is slow, with many of the oldest lines showing little regrowth of woody vegetation decades later [
10,
12]. This lack of natural recovery can occur due to conversion of some lines to temporary roads and Off-Highway Vehicle (OHV) trails [
10,
12], but there is also evidence that width and orientation of seismic lines influences regeneration by affecting abiotic factors (e.g., narrower lines and lines oriented east-to-west have improved regrowth of trees [
10]). Light and wind may play important roles in moderating responses to forest “dissection” (sensu Jaeger [
6]), as light availability can affect plant species assemblages [
8], while wind dynamics can alter plant community composition [
7,
26]. Estimating recovery trajectories will ultimately necessitate a better understanding of changes in the abiotic environment.
This study aims to determine the extent of abiotic environment changes within fragmented boreal forests from in situ oil sands developments in Alberta, Canada. Specifically, we evaluate whether and how sunlight intensity and wind speed differ on seismic lines compared with undisturbed forest controls, and in large reference openings (exploratory well pads). We further investigated how (i) line orientation, (ii) line width, and (iii) adjacent stand characteristics (approximated by canopy height) affect local variation in light and wind. We expected an increase in light intensity and wind speed when comparing seismic lines with reference undisturbed forests (controls). When assessing fragmented forest and seismic line characteristics, we hypothesized that (i) light intensity and wind speed will increase with magnitude (width) of disturbance; (ii) average and maximum wind speeds would be faster on east-west lines because the prevailing winds in the area are westerly; sunlight intensity will on average be higher on east-west orientated lines because they are more orientated with the movement of the sun during summer; (iii) taller forests will buffer changes in light intensity and wind speed in comparison to shorter forests; and (iv) increases in line width will increase light intensity and wind speed. Although these trends are expected, the magnitude of these differences by disturbance type (gap width of line) and their interaction are largely unknown, including whether low impact seismic lines reduce differences in abiotic conditions to that of forests compared to conventional seismic lines and that of forests.
4. Discussion
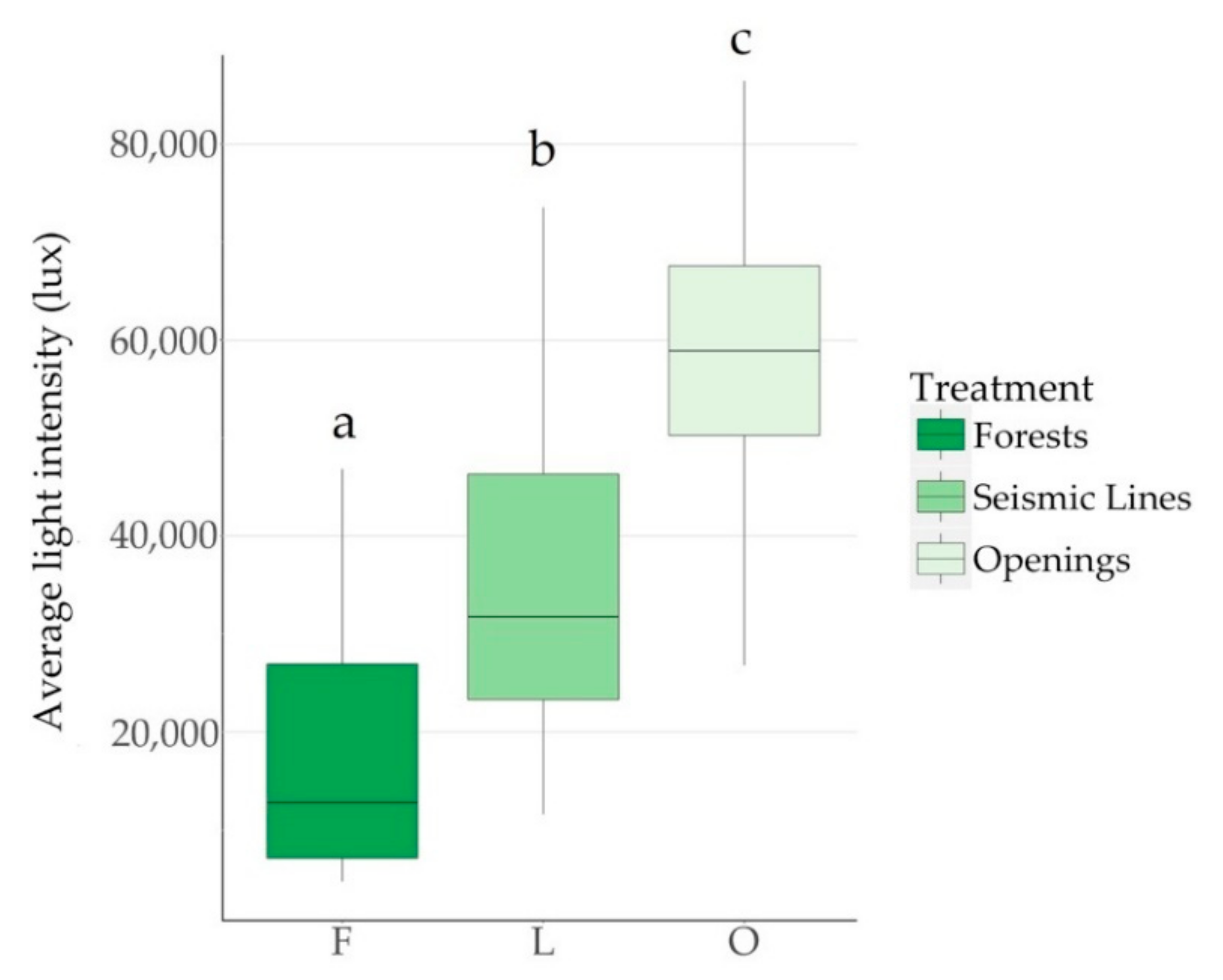
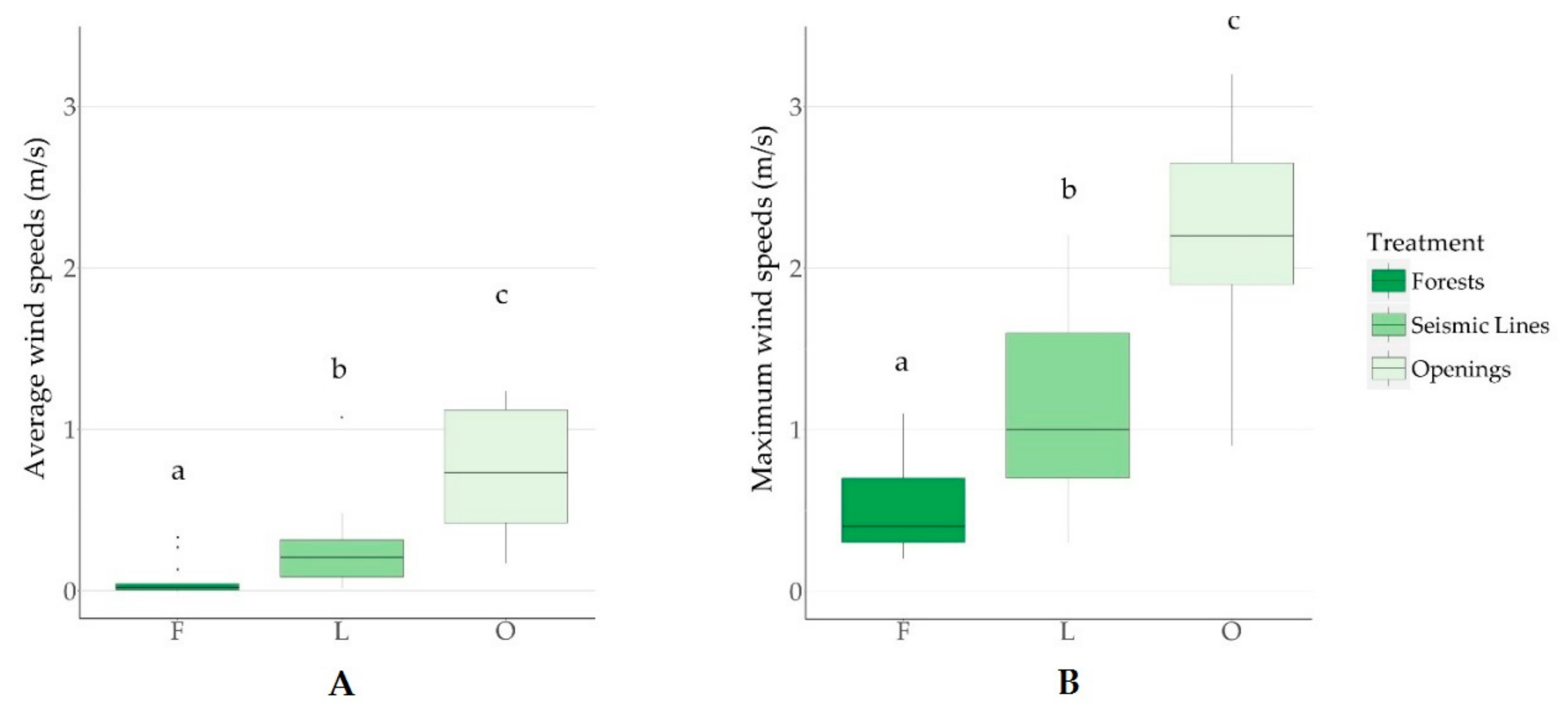
Forest disturbances associated with in situ oil sands exploration altered local environmental patterns of wind and light. Both seismic lines and well pads had a strong positive effect on light intensity, average wind speed, and maximum wind speed. On average the forest “dissection” [
6] (fragmentation) associated with these seismic line corridors doubled light intensity and maximum wind speed, while average wind speed was three times higher. Concurrently, exploratory well pads had approximately three times more light, 16 times higher average wind speed, and four times faster maximum wind than forests. The size of the well pad openings did not affect wind and light patterns, suggesting that a 60 × 60 m gap (smallest reference opening used in the study) is larger than a hypothetical “gap threshold”, after which the increase in light intensity and wind speed plateaus. Therefore, our results suggest that forest fragmentation associated with in situ oil sands exploration affects abiotic conditions (i.e., light intensity and wind speed) across a wide area of boreal forests, with even narrow seismic lines altering light and wind patterns. When focusing on the different characteristics of seismic line corridors, we found that light conditions within lines was best explained by a model including the interaction of gap width, canopy height, and line orientation. Lines oriented along an east-to-west axis had 22% greater average light intensity than lines oriented along a north-to-south axis. This concurs with our expectation as east-west lines are parallel to the path of the sun, such that the sun is less obstructed by canopy than in north-south lines. While gap width alone did not have a significant effect on light intensity, when interacting with canopy height the effect was significant. Thus, when canopy heights are tall and highly obstructive to sunlight, the width of the linear feature becomes a more important factor in explaining sunlight conditions along lines.
The result that seismic lines in the east-west direction are exposed to more light may contribute to explaining why van Rensen et al. (2015) [
10] identified increases in tree regeneration on east-west lines. The implications of these patterns in light intensity may be important not only for plants, but also for ectotherm organisms (e.g., insects). For instance, butterfly assemblages and behaviors have been shown to respond to these differences in anthropogenic disturbances [
22,
23] with similar responses also expected for other insects (e.g., Schultz 1998 [
32]). While the main response of these taxa to seismic lines is likely due to changes in habitat structure and composition, investigating the thermal ecology of species may reveal changes in species behavior and fitness, as well as more broadly in ecosystem processes and services (e.g., pollination [
33]).
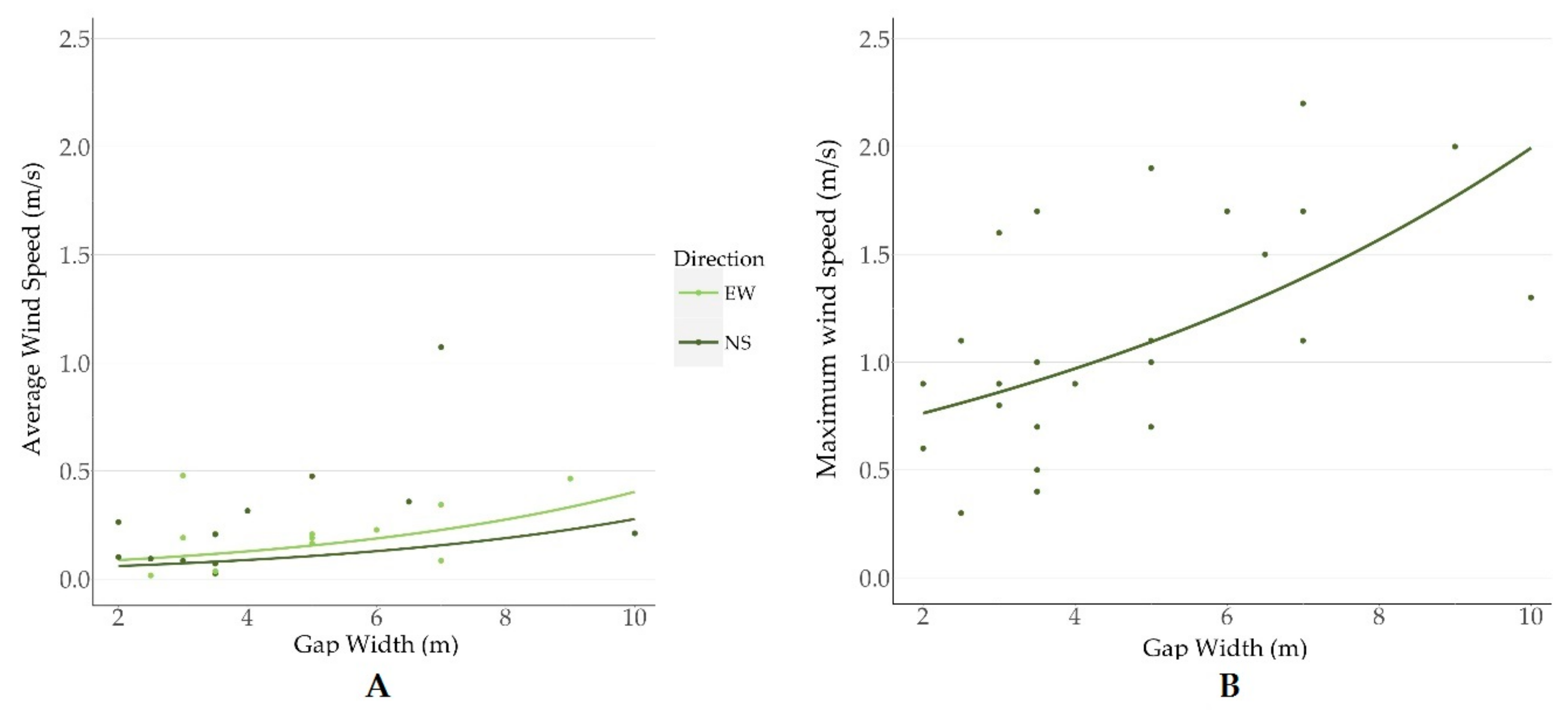
To understand the complex nature of wind dynamics, we separated wind into maximum and average values over 24-h periods. Specific wind characteristics influence various processes: average wind speed could gradually reduce soil moisture [
34], while maximum wind speed may affect dispersal of plant and animal species, as well as animal behavior and species interactions [
19,
35]. Notably, extreme maximum wind speeds can reduce canopy cover and increase tree mortality [
36], particularly in fragmented forests with more edges [
37]. Gap width was important in explaining wind conditions regardless of forest structure (i.e., canopy height). This suggests that, despite a minor structural difference, even seismic lines cleared across short forests elicit changes in wind dynamics analogous to those produced by seismic lines in taller forests. Specifically, the most supported model explaining average wind speed included line orientation and gap width as explanatory variables. We found that lines oriented along a north-to-south axis had 1.5 greater average wind speeds than those oriented east-to-west. This result was unexpected given the prevailing westerly winds in the region. Given the narrow time frame under which data were collected and the stochastic nature of wind dynamics, which vary between days and during the season, this result can be attributed to short-term wind patterns at the time of sampling. Consistently, gap width also had a positive effect on the maximum wind speed in these dissected forests. Maximum wind speeds were best explained by gap width alone, also suggesting that wind direction varied during our sampling sessions. The most supported model predicted that wide gaps (10 m) had 2.7 times faster maximum wind speeds than narrow gaps (2 m). Overall, these results corroborate those of Roberts et al. (2018) who found that wind speeds were 7 times higher on seismic lines than in forests, which led to seed dispersal that was 4 times farther in these linear features than in forests [
16].
Our finding that average wind speed was affected by direction of the linear disturbance was also consistent with Damschen [
19], who found that wind direction interacts with the orientation of structures originated by forest fragmentation. Therefore, seismic lines are acting as landscape features not only by directing animals that can respond to a change in forest structure with changes in behavioral (movement) patterns [
22,
27,
28], but also for species (animals and plants) that are dispersed by wind [
16]. As average wind speed was higher in seismic lines, these features may also be affecting other abiotic conditions including the moisture regime of these forests [
35]. This interaction of behavioral and environmental changes may trigger unexpected and complex patterns of responses in boreal species assemblages, with potential interactive effects of changes in behavior [
25] (including movement [
27]), habitat suitability [
22,
38], and species interactions across plant, invertebrates and vertebrates [
26].
Limitations restricted the scope of the study. The stochastic nature of weather in the region added to variation in the data, since the time frame over which the data were collected was relatively short and during a period of naturally fluctuating weather (i.e., summer). In particular, the random effect of day-to-day variations accounted for significant variation in average wind speeds. Future confirmation studies should focus on collecting data over a longer time frame and for more sites. A wider selection of gap types could also increase the understanding of these covariations.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




