Genome-Wide Identification and Characterization of MADS-box Family Genes Related to Floral Organ Development and Stress Resistance in Hevea brasiliensis Müll. Arg.

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. RNA Isolation and First-Strand cDNA Synthesis
2.3. Identification and Isolation of Mad-box Genes in H. brasiliensis
2.4. Protein Properties and Gene Structure Analysis of HbMad-box Genes
2.5. Multiple Sequence Alignments and Phylogenetic Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.7. Statistical Analysis
3. Results
3.1. Subsection Identification and Characterization of HbMad-box Genes in H. brasiliensis
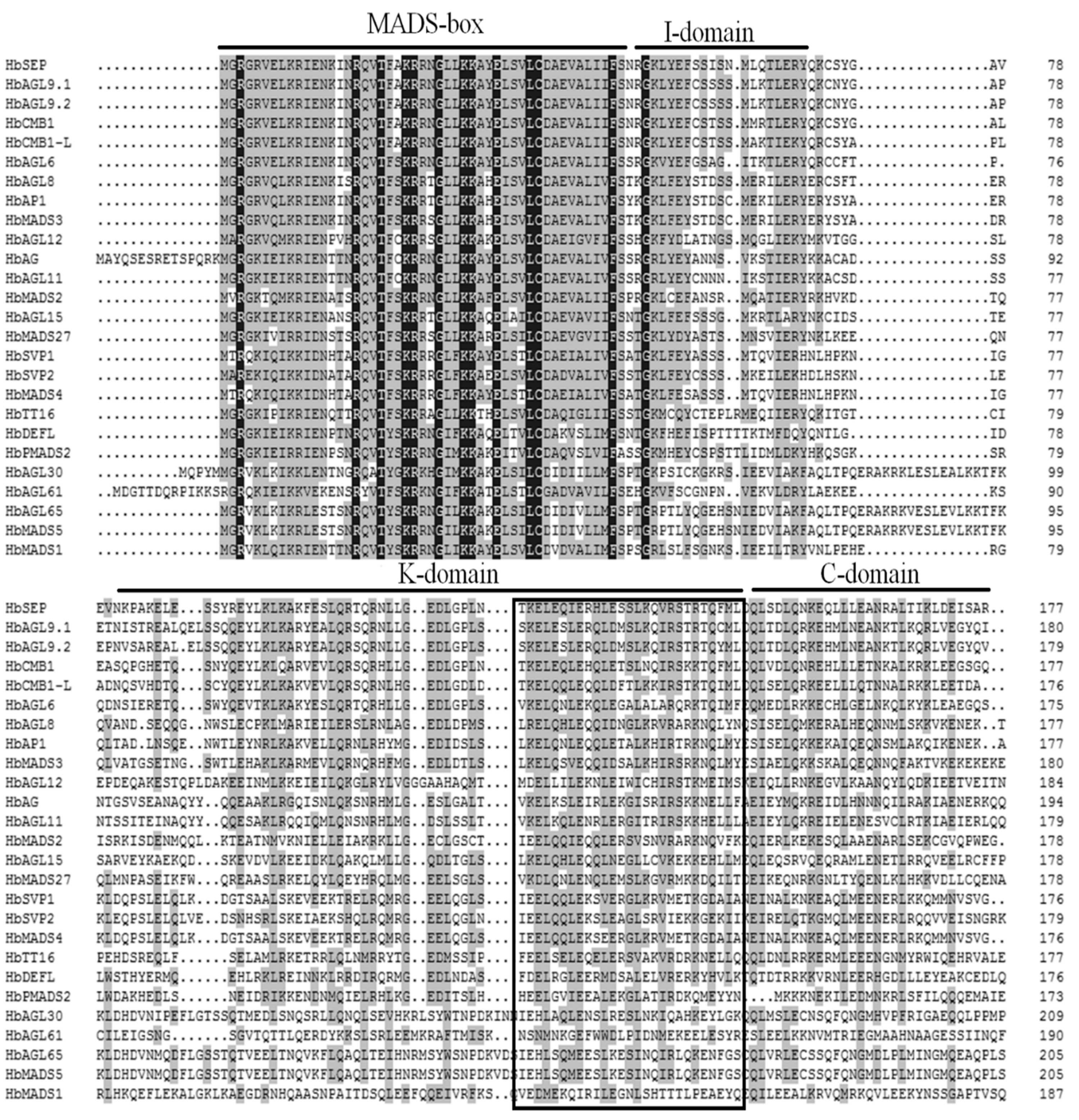
3.2. Analysis of Conserved Domains and Structural Features of HbMad-box Proteins
3.3. Gene Structures and Sequence Characteristics of HbMad-box Genes
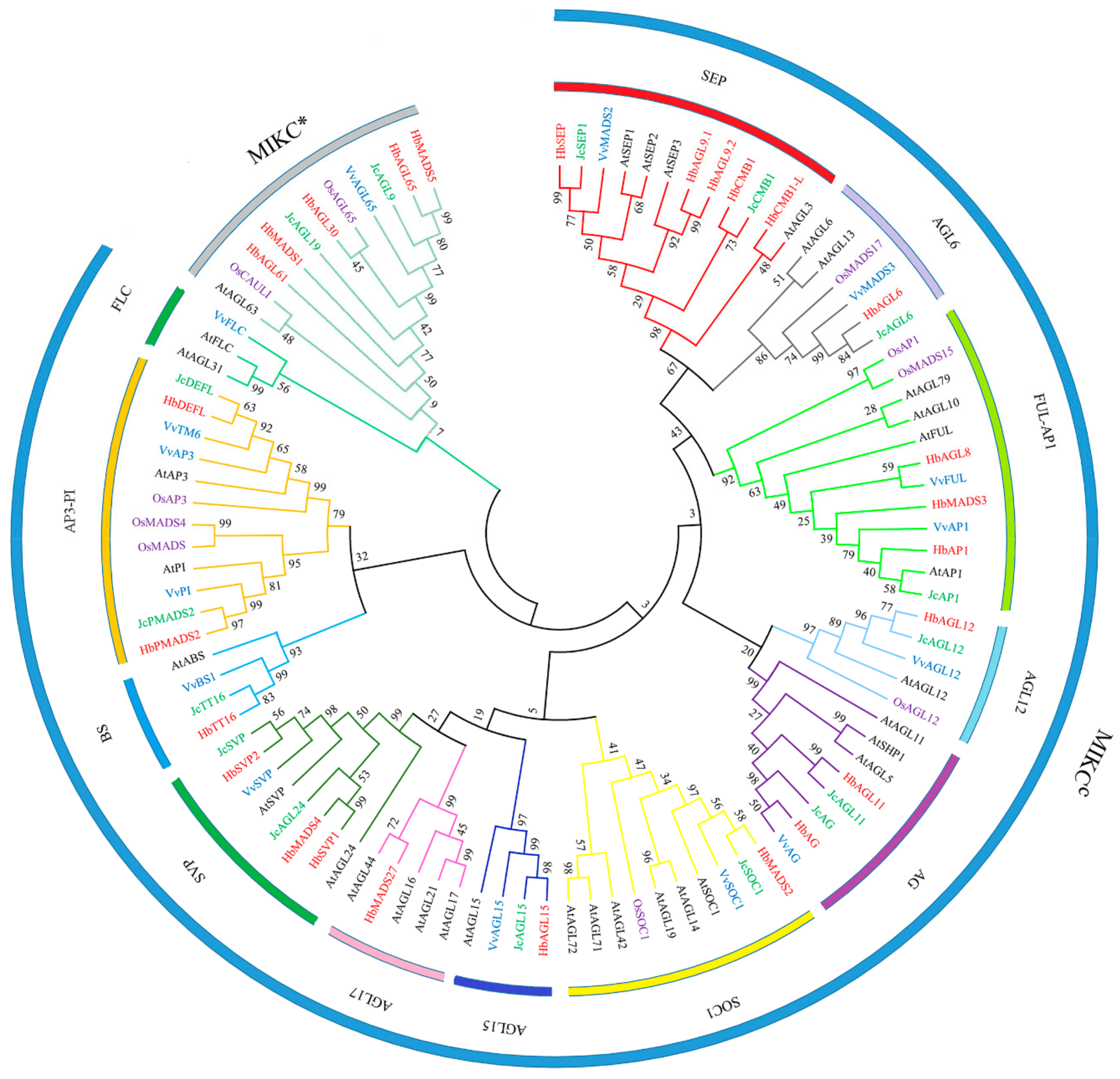
3.4. Phylogenetic Analysis of HbMad-box TFs
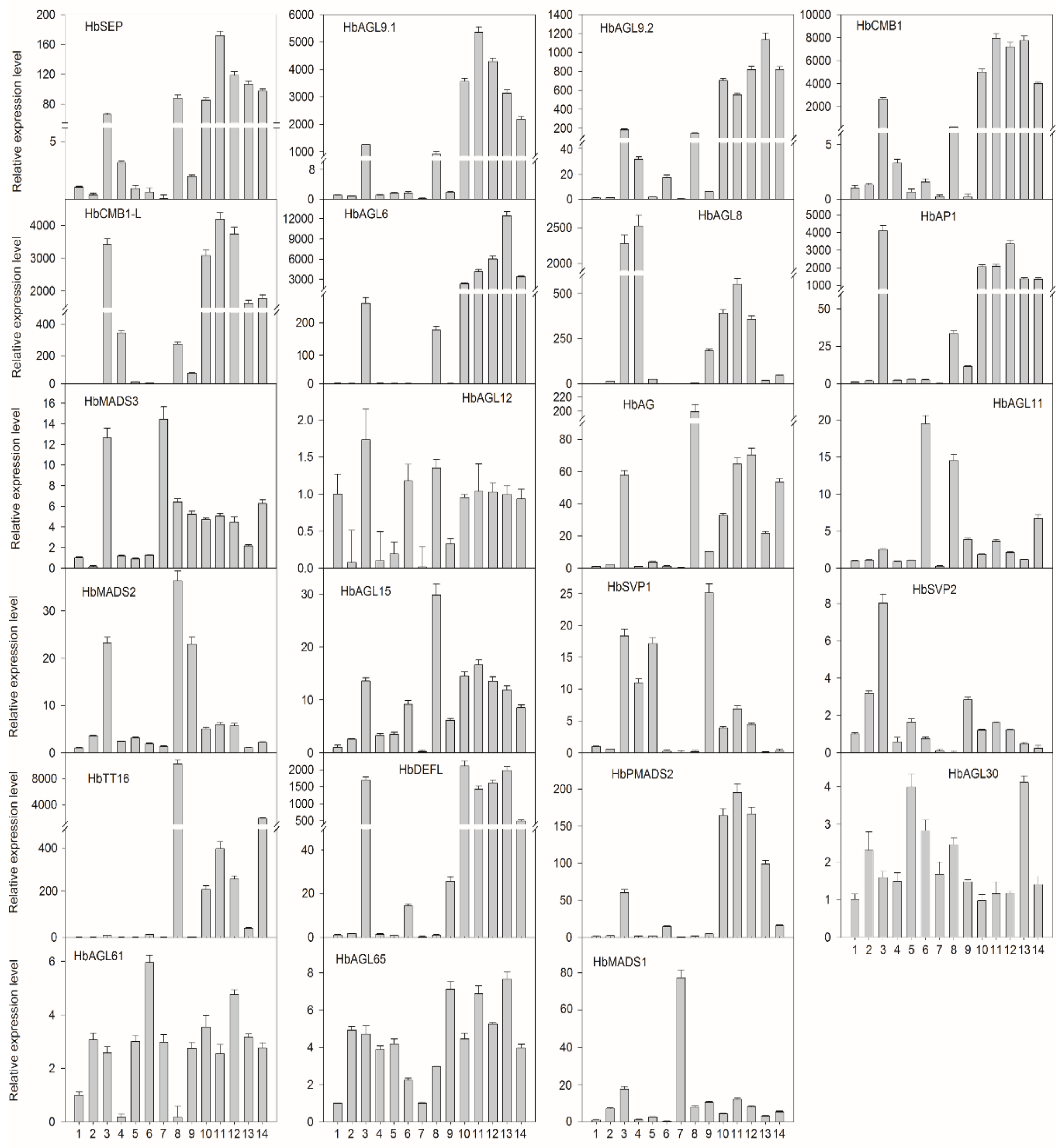
3.5. Expression Profiles of HbMad-box Genes in Various Developmental Stages during Rubber Tree Reproductive Development
3.6. Expression Patterns of HbMADS-box Genes in Response to Abiotic Stress
3.7. Expression Patterns of HbMADS-box Genes in Response to Phytohormones
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, D.; Zeng, R.; Yan, L.; Zhao, M.; Chao, J.; Yu, L. Gene expression analysis and SNP/InDel discovery to investigate yield heterosis of two rubber tree F1 hybrids. Sci. Rep. 2016, 6, 24984. [Google Scholar] [CrossRef] [PubMed]
- Mooibroek, H.; Cornish, K. Alternative sources of natural rubber. Appl. Microbiol. Biotechnol. 2000, 53, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Abnisaa, F.; Daud, W. Potential use of natural rubber to produce liquid fuels using hydrous pyrolysis—A review. RSC Adv. 2016, 6, 68906–68921. [Google Scholar] [CrossRef]
- Pethin, D.; Nakkanong, K.; Nualsri, C. Performance and genetic assessment of rubber tree clones in Southern Thailand. Sci. Agric. 2015, 72, 306–313. [Google Scholar] [CrossRef]
- Ahrends, A.; Hollingsworth, P.M.; Ziegler, A.D.; Fox, J.M.; Chen, H.; Su, Y. Current trends of rubber plantation expansion may threaten biodiversity and livelihoods. Glob. Environ. Chang. 2015, 34, 48–58. [Google Scholar] [CrossRef]
- Dornelas, M.C.; Rodriguez, A.P. The rubber tree (Hevea brasiliensis Muell. Arg.) homologue of the LEAFY/FLORICAULA gene is preferentially expressed in both male and female floral meristems. J. Exp. Bot. 2005, 56, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Cubry, P.; Pujade-Renaud, V.; Garcia, D.; Espeout, S.; Le, G.V.; Granet, F. Development and characterization of a new set of 164 polymorphic EST-SSR markers for diversity and breeding studies in rubber tree (Hevea brasiliensis Mull. Arg.). Plant Breed. 2014, 133, 419–426. [Google Scholar] [CrossRef] [Green Version]
- Ó’Maoiléidigh, D.S.; Wuest, S.E.; Rae, L.; Raganelli, A.; Ryan, P.T.; Kwaśniewska, K.; Wellmer, F. Control of reproductive floral organ identity specification in Arabidopsis by the C function regulator AGAMOUS. Plant Cell 2013, 25, 2482–2503. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Ito, T.; Wellmer, F.; Meyerowitz, E.M. Repression of AGAMOUS-LIKE 24 is a crucial step in promoting flower development. Nat. Genet. 2004, 36, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Patharkar, O.R.; Walker, J.C. Floral organ abscission is regulated by a positive feedback loop. PNAS 2015, 112, 2906–2911. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, B.J.; Lehtishiu, M.D.; Fernandez, D.E. The MADS domain factors AGL15 and AGL18 act redundantly as repressors of the floral transition in Arabidopsis. Plant J. 2007, 50, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- García-Maroto, F.; Carmona, M.J.; Garrido, J.A.; Vilches-Ferrón, M.; Rodríguez-Ruiz, J.; Alonso, D.L. New roles for MADS-box genes in higher plants. Biol. Plant. 2003, 46, 321–330. [Google Scholar] [CrossRef]
- Nam, J.; Ma, H.; Nei, M. Antiquity and evolution of the MADS-box gene family controlling flower development in plants. Mol. Biol. Evol. 2003, 20, 1435–1447. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.E.; Singh, M.B.; Bhalla, P.L. Novel members of the AGAMOUS LIKE 6 subfamily of MIKCc-type MADS-box genes in soybean. BMC Plant Biol. 2013, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Leseberg, C.H.; Li, A.; Kang, H.; Duvall, M.; Mao, L. Genome-wide analysis of the MADS-box gene family in populus trichocarpa. Gene 2006, 378, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Kofuji, R.; Sumikawa, N.; Yamasaki, M.; Kondo, K.; Ueda, K.; Ito, M.; Hasebe, M. Evolution and divergence of the MADS-box gene family based on genome-wide expression analyses. Mol. Biol. Evol. 2003, 20, 1963–1977. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.; Fornara, F.; Vincent, C.; Andrés, F.; Nordström, K.; Göbel, U. Analysis of the Arabidopsis shoot meristem transcriptome during floral transition identifies distinct regulatory patterns and a leucine-rich repeat protein that promotes flowering. Plant Cell 2012, 24, 444–462. [Google Scholar] [CrossRef] [PubMed]
- Honma, T.; Goto, K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 2001, 409, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Lalusin, A.G.; Nishita, K.; Kim, S.H.; Ohta, M.; Fujimura, T. A new MADS-box gene (IbMADS10) from sweet potato (Ipomoea batatas L. Lam) is involved in the accumulation of anthocyanin. Mol. Genet. Genom. 2006, 275, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Su, X.; Zhou, X. A MADS-box gene of Populus deltoides expressed during flower development and in vegetative organs. Tree Physiol. 2008, 28, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xu, B.; Hu, L.; Li, M.; Su, W.; Wu, J. Involvement of a banana MADS-box transcription factor gene in ethylene-induced fruit ripening. Plant Cell Rep. 2009, 28, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Tani, E.; Polidoros, A.N.; Tsaftaris, A.S. Characterization and expression analysis of Fruitfull- and shatterproof-like genes from peach (Prunus persica) and their role in split-pit formation. Tree Physiol. 2007, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Zhao, D.; Han, C.; Wang, J.; Hao, Z.; Tao, J. Cloning and expression of floral organ development-related genes in herbaceous peony (Paeonia lactiflora, Pall.). Mol. Biol. Rep. 2014, 41, 6493–6530. [Google Scholar] [CrossRef] [PubMed]
- Bemer, M.; Heijmans, K.; Airoldi, C.; Davies, B.; Angenent, G. An atlas of type I MADS box gene expression during female gametophyte and seed development in Arabidopsis. Plant Physiol. 2010, 154, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Thakare, D.; Tang, W.; Hill, K.; Perry, S.E. The MADS-domain transcriptional regulator AGAMOUS-LIKE15 promotes somatic embryo development in Arabidopsis and soybean. Plant Physiol. 2008, 146, 1663–1672. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Masiero, S.; Vanzulli, S.; Lardelli, P.; Kater, M. M.; Colombo, L. AGL23, a type I MADS-box gene that controls female gametophyte and embryo development in Arabidopsis. Plant J. 2008, 54, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Woo, Y.; Ryu, S.; Shin, Y.; Kim, W.; Park, K.; Lee, I.; An, G. Further characterization of a rice AGL12 group MADS-Box gene, OsMADS26. Plant Physiol. 2008, 147, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Elitzur, T.; Vrebalov, J.; Giovannoni, J.J.; Goldschmidt, E.E.; Friedman, H. The regulation of MADS-box gene expression during ripening of banana and their regulatory interaction with ethylene. J. Exp. Bot. 2010, 61, 1523–1535. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.; Angosto, T.; Gomez, P.; Payan, C.; Capel, J.; Huijser, P. Tomato flower abnormalities induced by low temperatures are associated with changes of expression of MADS-Box genes. Plant Physiol. 1998, 117, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K. MADS-box gene family in rice: Genome-wide identification, organization and expression profiling during reproductive development and stress. BMC Genom. 2007, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, F.; Kurz, B.; Melzer, S.; Bernier, G.; Jacqmard, A. Cytokinin and gibberellin activate SaMADS A, a gene apparently involved in regulation of the floral transition in Sinapis alba. Plant J. 2000, 24, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Ando, S.; Sato, Y.; Kamachi, S.; Sakai, S. Isolation of a MADS-box gene (ERAF17) and correlation of its expression with the induction of formation of female flowers by ethylene in cucumber plants (Cucumis sativus L.). Planta 2001, 213, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Perry, S.E. Control of expression and auto regulation of AGL15, a member of the MADS-box family. Plant J. 2005, 41, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Wei, L.R.; Guo, D.; Wang, Y.; Zhu, J.H.; Chen, X.T. HbMADS4, a MADS-box transcription factor from Hevea brasiliensis, negatively regulates HbSRPP. Front. Plant Sci. 2016, 7, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.B.; Tu, M.; Hua, Y.W.; Huang, H.S. Cloning and expression analysis of HbAGL62 of rubber tree (Hevea brasiliensis Arg.). Chin. J. Trop. Crops. 2012, 33, 1766–1771. [Google Scholar]
- Wang, H.; Li, L.; Liang, Z.; Wu, Y.J. Cloning and bioinformatic analysis of the gene HbMADS-27 from Hevea brasiliensis. Chin. J. Trop. Agric. 2013, 33, 19–24. [Google Scholar]
- Li, H.L.; Wang, Y.; Guo, D.; Tian, W.M.; Peng, S.Q. Three MADS-box genes of Hevea brasiliensis expressed during somatic embryogenesis and in the laticifer cells. Mol. Biol. Rep. 2011, 38, 4045–4052. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Chen, X.; Zhu, J.S.; Huang, H.S. Overexpression of a Hevea brasiliensis ErbB-3 Binding protein 1 Gene Increases Drought Tolerance and Organ Size in Arabidopsis. Front. Plant Sci. 2016, 7, 1703. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Aileni, M.; Abbagani, S.; Zhang, P. A reliable and efficient method for total RNA isolation from various members of spurge family (Euphorbiaceae). Phytochem. Anal. 2010, 21, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, V. Statistical Approaches in Eukaryotic Gene Prediction. In Handbook of Statistical Genetics; John Wiley & Sons, Ltd.: Clifton, NJ, USA, 2004; pp. 7–39. [Google Scholar]
- Tang, C.; Huang, D.; Yang, J.; Liu, S.; Sakr, S.; Li, H. The sucrose transporter HbSUT3 plays an active role in sucrose loading to laticifer and rubber productivity in exploited trees of Hevea brasiliensis (para rubber tree). Plant Cell Environ. 2010, 33, 1708–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, X.; He, B.; Wang, C.; Fang, Y.; Qi, J.; Tang, C. Molecular identification and characterization of the pyruvate decarboxylase gene family associated with latex regeneration and stress response in rubber tree. Plant Physiol. Biochem. 2015, 87, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, T.; Saito, M.; Yamada, E.; Fujita, K.; Yamagishi, N.; Yoshikawa, N. Isolation and characterization of the C-class MADS-box gene involved in the formation of double flowers in Japanese gentian. BMC Plant Biol. 2015, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, D.; Ping, J.; Li, S.; Chen, Z.; Ma, J. Innovation of a regulatory mechanism modulating semi-determinate stem growth through artificial selection in soybean. PLoS Genet. 2016, 12, e1005818. [Google Scholar] [CrossRef] [PubMed]
- Par Enicová, L.; Folter, S.D.; Kieffer, M.; Horner, D.S.; Favalli, C.; Busscher, J. Molecular and Phylogenetic Analyses of the Complete MADS-Box Transcription Factor Family in Arabidopsis: New openings to the MADS world. Plant Cell 2003, 15, 1538–1551. [Google Scholar] [CrossRef]
- Díazriquelme, J.; Lijavetzky, D.; Martínezzapater, J.M.; Carmona, M.J. Genome-wide analysis of MIKCc-type MADS-box genes in grapevine. Plant Physiol. 2009, 149, 354–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinta, G.; Khan, A.; Abdelgawad, H.; Verma, V.; Srivastava, A.K. Unveiling the redox control of plant reproductive development during abiotic stress. Front. Plant Sci. 2016, 7, 700. [Google Scholar] [CrossRef] [PubMed]
- Fromm, S.; Braun, H.; Peterhansel, C. Mitochondrial gamma carbonic anhydrases are required for complex I assembly and plant reproductive development. New Phytol. 2016, 211, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.M.; Huang, T.; Huang, Y.; Chen, D.H. Phylogenetic analysis of MADS-box gene family in Physcomitrella patens. Plant Physiol. J. 2015, 51, 197–206. [Google Scholar] [CrossRef]
- Saha, G.; Park, J.I.; Jung, H.J.; Ahmed, N.U. Genome-wide identification and characterization of MADS-box family genes related to organ development and stress resistance in Brassica rapa. BMC Genom. 2015, 16, 178. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Arya, P.; Gupta, K.; Randhawa, V.; Acharya, V.; Singh, A.K. Comparative phylogenetic analysis and transcriptional profiling of MADS-box gene family identified DAM and FLC-like genes in apple (Malusx domestica). Sci. Rep. 2016, 6, 20695. [Google Scholar] [CrossRef] [PubMed]
- Grimplet, J.; Martínez-Zapater, J.M.; Carmona, M.J. Structural and functional annotation of the MADS-box transcription factor family in grapevine. BMC Genom. 2016, 17, 80. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.; Kim, J.; Lee, S.; An, G.; Ma, H.; Nei, M. Type I MADS-box genes have experienced faster birth-and-death evolution than type II MADS-box genes in angiosperms. PNAS 2004, 101, 1910–1915. [Google Scholar] [CrossRef] [PubMed]
- Smaczniak, C.; Immink, R.G.; Angenent, G.C.; Kaufmann, K. Developmental and evolutionary diversity of plant MADS-domain factors: Insights from recent studies. Development 2012, 139, 3081–3098. [Google Scholar] [CrossRef] [PubMed]
- Masiero, S.; Li, M.A.; Will, I.; Hartmann, U.; Saedler, H.; Huijser, P. Incomposita: A MADS-box gene controlling prophyll development and floral meristem identity in antirrhinum. Development 2004, 131, 5981–5990. [Google Scholar] [CrossRef] [PubMed]
- Toshitsugu, N.; Junji, K.; Masaki, F.; Mamiko, K.; Nao, I.; Hideo, M. Macrocalyx and jointless interact in the transcriptional regulation of tomato fruit abscission zone development. Plant Physiol. 2012, 158, 439–450. [Google Scholar] [CrossRef]
- Huang, Q.; Li, W.; Fan, R.; Chang, Y. New mads-box gene in fern: Cloning and expression analysis of dfmads1 from Dryopteris fragrans. PLoS ONE 2014, 9, e86349. [Google Scholar] [CrossRef] [PubMed]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fiume, E.; Coen, O.; Pechoux, C.; Lepiniec, L.; Magnani, E. Endosperm and nucellus develop antagonistically in Arabidopsis seeds. Plant Cell 2016, 28, 1343–1360. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Deng, W.; Peng, F.; Truksa, M.; Singer, S.; Snyder, C.L. Brassica napus TT16 homologs with different genomic origins and expression levels encode proteins that regulate a broad range of endothelium-associated genes at the transcriptional level. Plant J. 2013, 74, 663–677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, H.; Zhang, D.; Liu, Y.; Fu, J.; Shi, Y. Characterization and expression analysis of six MADS-box genes in maize (Zea mays L.). J. Plant Physiol. 2012, 169, 797–806. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Accession No. | ORF Length (bp) | Protein | ||
|---|---|---|---|---|---|
| Length (aa) | Mw (kDa) | PI | |||
| HbSEP | KY471153 | 732 | 243 | 28.00 | 8.88 |
| HbAGL9.1 | KY471149 | 732 | 243 | 27.77 | 8.21 |
| HbAGL9.2 | KY471165 | 723 | 240 | 27.40 | 8.8 |
| HbCMB1 | KY471159 | 729 | 242 | 27.57 | 8.36 |
| HbCMB1-L | KY471157 | 729 | 242 | 27.87 | 7.6 |
| HbAGL6 | KY471160 | 729 | 242 | 27.63 | 8.73 |
| HbAGL8 | KY471148 | 759 | 252 | 29.16 | 7.72 |
| HbAP1 | KY471161 | 726 | 241 | 28.16 | 8.19 |
| HbMADS3 | GU142915 | 720 | 239 | 27.61 | 9.16 |
| HbAGL12 | KY471152 | 609 | 202 | 23.09 | 6.92 |
| HbAG | KY471158 | 726 | 241 | 27.78 | 9.08 |
| HbAGL11 | KY471166 | 669 | 222 | 25.52 | 9.34 |
| HbMADS2 | GU142914 | 654 | 217 | 24.93 | 9.23 |
| HbAGL15 | KY471151 | 768 | 255 | 29.27 | 6.68 |
| HbMADS27 | KY486278 | 717 | 238 | 27.49 | 9.25 |
| HbSVP1 | KY471154 | 633 | 210 | 23.69 | 6.86 |
| HbSVP2 | KY471156 | 702 | 234 | 26.60 | 6.06 |
| HbMADS4 | KY486276 | 633 | 210 | 23.67 | 6.36 |
| HbTT16 | KY471155 | 705 | 234 | 27.46 | 7.69 |
| HbDEFL | KY471162 | 675 | 224 | 26.23 | 8.43 |
| HbPMADS2 | KY471147 | 639 | 212 | 25.12 | 8.4 |
| HbAGL30 | KY471163 | 1116 | 371 | 41.86 | 7.14 |
| HbAGL61 | KY471164 | 642 | 213 | 24.24 | 8.32 |
| HbAGL65 | KY471150 | 741 | 246 | 28.41 | 8.68 |
| HbMADS5 | KY486277 | 1077 | 358 | 40.74 | 6.38 |
| HbMADS1 | GU142913 | 738 | 245 | 27.78 | 8.34 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, M.; Wang, Y.; Pan, R.; Li, W. Genome-Wide Identification and Characterization of MADS-box Family Genes Related to Floral Organ Development and Stress Resistance in Hevea brasiliensis Müll. Arg. Forests 2018, 9, 304. https://doi.org/10.3390/f9060304
Wei M, Wang Y, Pan R, Li W. Genome-Wide Identification and Characterization of MADS-box Family Genes Related to Floral Organ Development and Stress Resistance in Hevea brasiliensis Müll. Arg. Forests. 2018; 9(6):304. https://doi.org/10.3390/f9060304
Chicago/Turabian StyleWei, Mingming, Yajie Wang, Ranran Pan, and Weiguo Li. 2018. "Genome-Wide Identification and Characterization of MADS-box Family Genes Related to Floral Organ Development and Stress Resistance in Hevea brasiliensis Müll. Arg." Forests 9, no. 6: 304. https://doi.org/10.3390/f9060304
APA StyleWei, M., Wang, Y., Pan, R., & Li, W. (2018). Genome-Wide Identification and Characterization of MADS-box Family Genes Related to Floral Organ Development and Stress Resistance in Hevea brasiliensis Müll. Arg. Forests, 9(6), 304. https://doi.org/10.3390/f9060304