Soil Nitrogen Responses to Soil Core Transplanting Along an Altitudinal Gradient in an Eastern Tibetan Forest



Abstract
1. Introduction
2. Materials and Methods
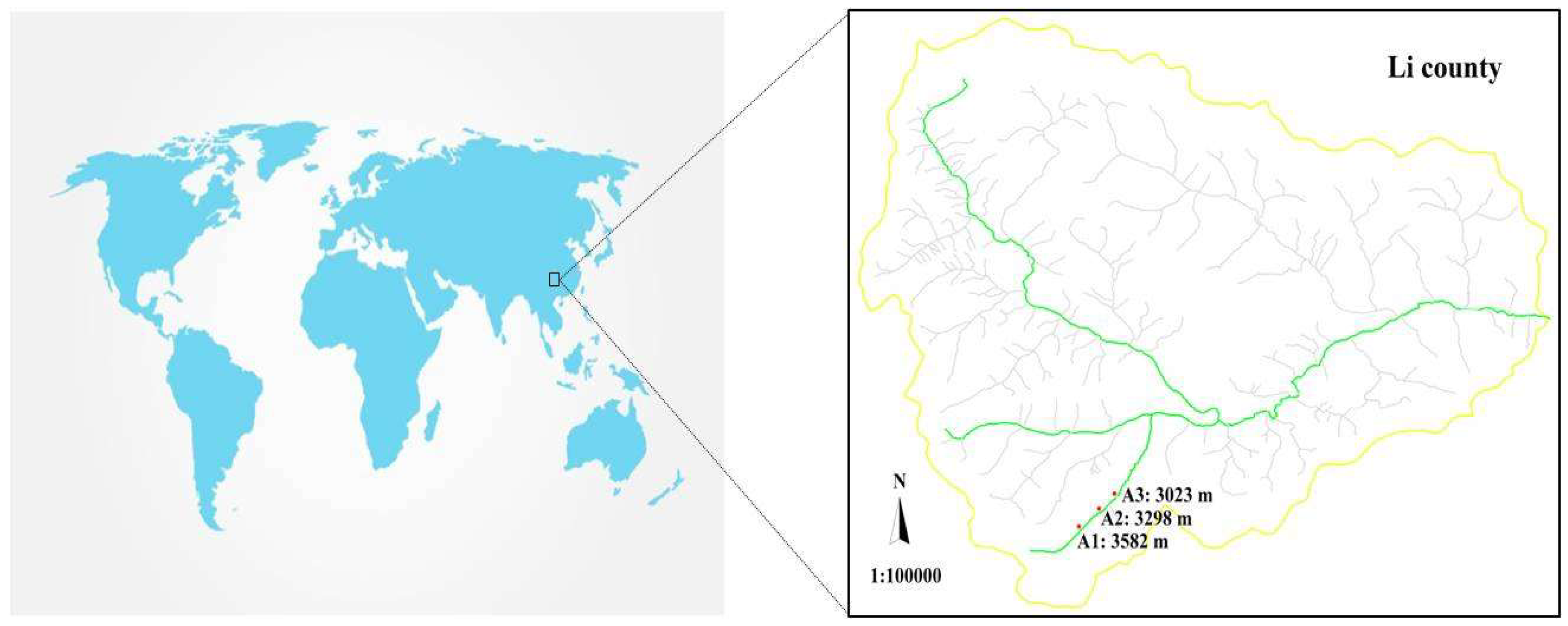
2.1. Site Description
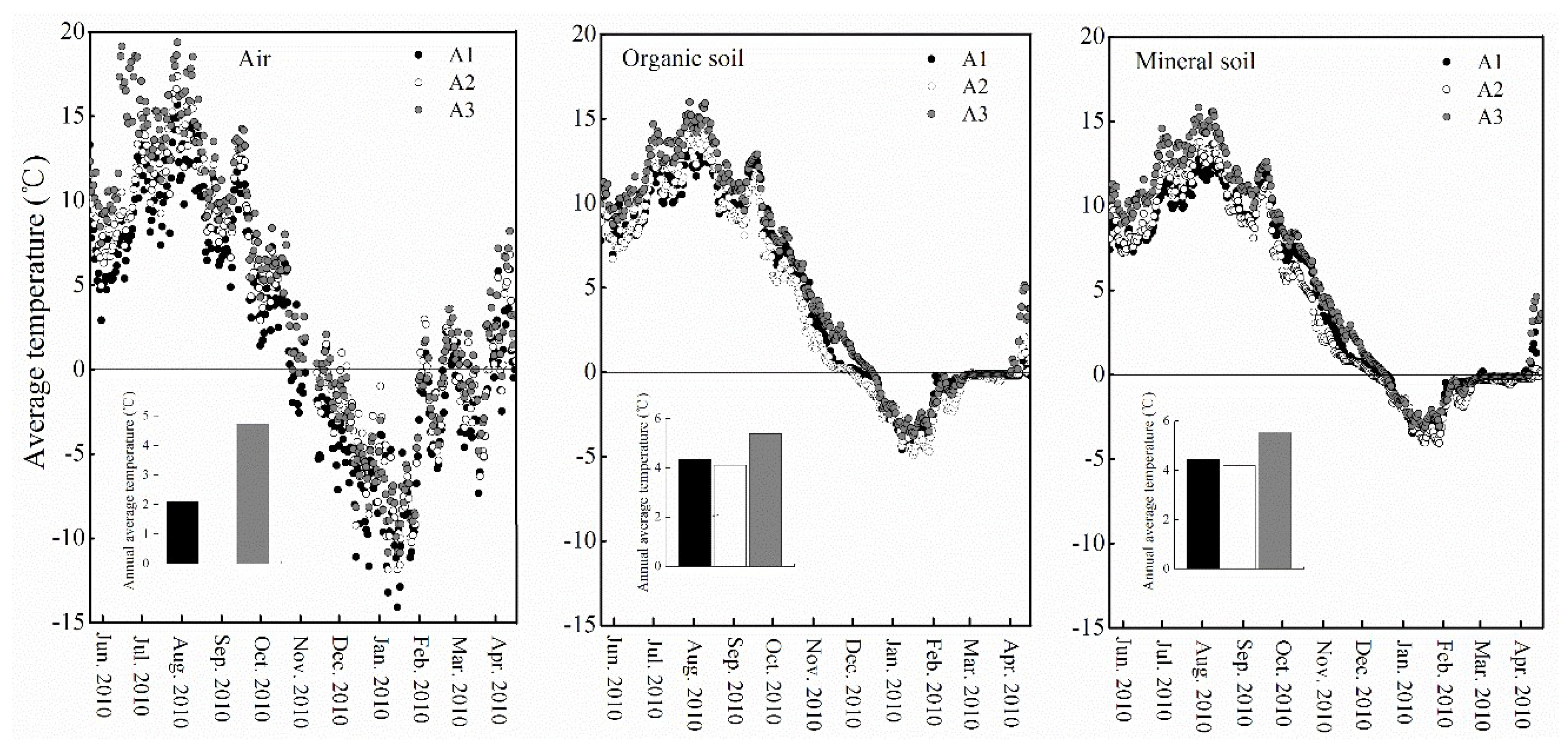
2.2. Temperature Monitoring
2.3. Soil Incubation
2.4. Sample Collection
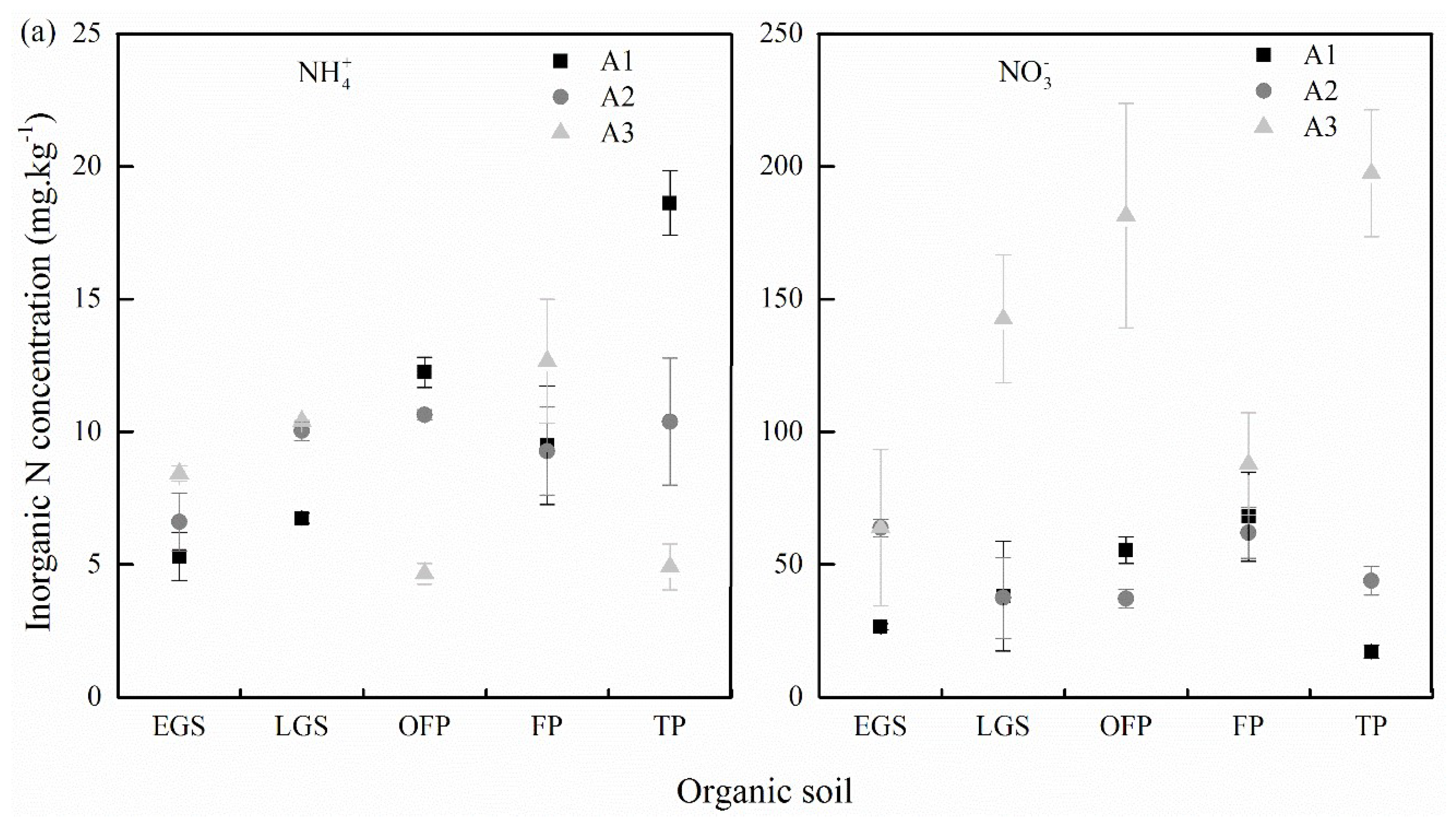
2.5. Inorganic Nitrogen Concentration
2.6. DNA Extraction
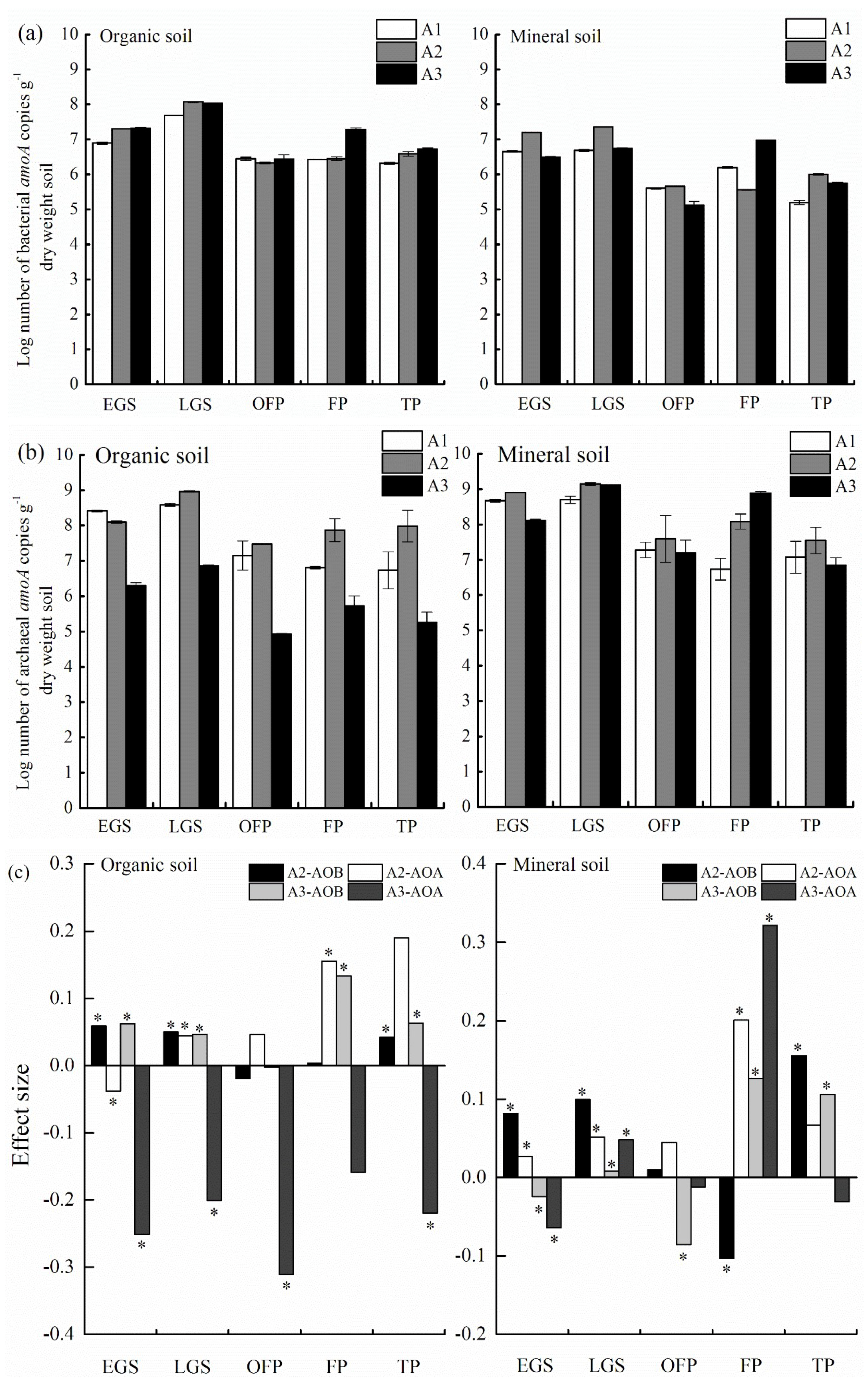
2.7. Quantification of amoA Genes by Real-Time PCR
2.8. Statistical Analyses
3. Results
3.1. Air and Soil Temperatures
3.2. Inorganic Nitrogen Concentration
3.3. Abundance of amoA Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tan, B.; Wu, F.Z.; Yang, W.Q.; He, X.H. Snow removal alters soil microbial biomass and enzyme activity in a Tibetan alpine forest. Appl. Soil Ecol. 2014, 76, 34–41. [Google Scholar] [CrossRef]
- Xu, Z.F.; Hu, R.; Xiong, P.; Wan, C.; Cao, G.; Liu, Q. Initial soil responses to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China: Nutrient availabilities, microbial properties and enzyme activities. Appl. Soil Ecol. 2010, 46, 291–299. [Google Scholar] [CrossRef]
- Xu, Z.F.; Liu, Q.; Yin, H.J. Effects of temperature on soil net nitrogen mineralisation in two contrasting forests on the eastern Tibetan Plateau, China. Soil Res. 2014, 52, 562–567. [Google Scholar] [CrossRef]
- Zhang, L.M.; Wang, M.; Prosser, J.I.; Zheng, Y.M.; He, J.Z. Altitude ammonia-oxidizing bacteria and archaea in soils of Mount Everest. FEMS Microbiol. Ecol. 2009, 70, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.C.; Scalenghe, R.; Freppaz, M. Changes in the seasonal snow cover of alpine regions and its effect on soil processes: A review. Quat. Int. 2007, 162, 172–181. [Google Scholar] [CrossRef]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D. Colder soils in a warmer world: A snow manipulation study in a northern hardwood forest ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Williams, M.W.; Helmig, D.; Blanken, P. White on green: Under-snow microbial processes and trace gas fluxes through snow, Niwot Ridge, Colorado Front Range. Biogeochemistry 2009, 95, 1–12. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, W.; He, X. Temporal dynamics of abiotic and biotic factors on leaf litter of three plant species in relation to decomposition rate along a subalpine elevation gradient. PLoS ONE 2013, 8, e62073. [Google Scholar] [CrossRef] [PubMed]
- Hungate, B.A.; Dukes, J.S.; Shaw, M.R.; Luo, Y.; Field, C.B. Atmospheric science. Nitrogen and climate change. Science 2003, 302, 1512–1513. [Google Scholar] [CrossRef] [PubMed]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef] [PubMed]
- Groffman, P.M.; Hardy, J.P.; Fashu-Kanu, S.; Driscoll, C.T.; Cleavitt, N.L.; Fahey, T.J.; Fisk, M.C. Snow depth, soil freezing and nitrogen cycling in a northern hardwood forest landscape. Biogeochemistry 2011, 102, 223–238. [Google Scholar] [CrossRef]
- Campbell, J.L.; Mitchell, M.J.; Groffman, P.M.; Christenson, L.M.; Hardy, J.P. Winter in northeastern North America: A critical period for ecological processes. Front. Ecol. Environ. 2005, 3, 314–322. [Google Scholar] [CrossRef]
- Freppaz, M.; Williams, B.L.; Edwards, A.C.; Scalenghe, R.; Zanini, E. Simulating soil freeze/thaw cycles typical of winter alpine conditions: Implications for N and P availability. Appl. Soil Ecol. 2007, 35, 247–255. [Google Scholar] [CrossRef]
- Kreyling, J. Winter climate change: A critical factor for temperate vegetation performance. Ecology 2010, 91, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Prosser, J.I. Autotrophic nitrification in bacteria. Adv. Microb. Physiol. 1989, 30, 125–181. [Google Scholar] [CrossRef] [PubMed]
- He, J.Z.; Shen, J.P.; Zhang, L.M.; Zhu, Y.G.; Zheng, Y.M.; Xu, M.G.; Di, H.J. Quantitative analyses of the abundance and composition of ammonia-oxidizing bacteria and ammonia-oxidizing archaea of a Chinese upland red soil under long-term fertilization practices. Environ. Microbiol. 2007, 9, 2364–2374. [Google Scholar] [CrossRef] [PubMed]
- Könneke, M.; Bernhard, A.E.; de la Torre, J.R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, W.; Prosser, J.I.; Schuster, S.C.; Schleper, C. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 2006, 442, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Reigstad, L.J.; Richter, A.; Daims, H.; Urich, T.; Schwark, L.; Schleper, C. Nitrification in terrestrial hot springs of Iceland and Kamchatka. FEMS Microbiol. Ecol. 2008, 64, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Auguet, J.C.; Nomokonova, N.; Camarero, L.; Casamayor, E.O. Seasonal changes of freshwater ammonia-oxidizing archaeal assemblages and nitrogen species in oligotrophic alpine lakes. Appl. Environ. Microbiol. 2011, 77, 1937–1945. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Wu, F.Z.; Yang, W.Q.; Wu, Z.C.; Wang, X.X.; Tan, B. Abundance and composition dynamics of soil ammonia-oxidizing archaea in an alpine fir forest on the eastern Tibetan Plateau of China. Can. J. Microbiol. 2012, 58, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Wu, F.Z.; He, Z.H.; Xu, Z.F.; Liu, Y.; Tan, B.; Yang, W.Q. Characteristics of ammonia-oxidizing bacteria and ammonia-oxidizing archaea abundance in soil organic layer under the subslpine/alpine forest. Acta Ecol. Sin. 2012, 32, 4371–4378. [Google Scholar] [CrossRef][Green Version]
- Henry, H.A.L. Soil freeze–thaw cycle experiments: Trends, methodological weaknesses and suggested improvements. Soil Biol. Biochem. 2007, 39, 977–986. [Google Scholar] [CrossRef]
- Long, X.; Chen, C.; Xu, Z.; Linder, S.; He, J. Abundance and community structure of ammonia oxidizing bacteria and archaea in a Sweden boreal forest soil under 19-year fertilization and 12-year warming. J. Soils Sediments 2012, 12, 1124–1133. [Google Scholar] [CrossRef]
- Avrahami, S.; Conrad, R. Patterns of community change among ammonia oxidizers in meadow soils upon long-term inubation at different temperatures. Appl. Environ. Microbiol. 2003, 69, 6152–6164. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, B. Climatic warming in the Tibetan Plateau during recent decades. Int. J. Climatol. 2000, 20, 1729–1742. [Google Scholar] [CrossRef]
- Chen, H.; Zhu, Q.; Peng, C.; Wu, N.; Wang, Y.; Fang, X.; Gao, Y.; Zhu, D.; Yang, G.; Tian, J.; et al. The impacts of chimate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau. Glob. Chang. Biol. 2013, 19, 2940–2955. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.Q.; Feng, R.F.; Zhang, J.; Wang, K.Y. Carbon stock and biochemical properties in the organic layer and mineral soil under three subalpine forests in Western China. Acta Ecol. Sin. 2007, 27, 4157–4165. [Google Scholar] [CrossRef]
- Feng, R.F.; Yang, W.Q.; Zhang, J. Review on biochemical property in forest soil organic layer and its responses to climate change. Chin. J. Appl. Environ. Biol. 2006, 12, 734–739. [Google Scholar] [CrossRef]
- Gong, Z.T.; Zhang, G.L.; Chen, Z.C. Pedogenesis and Soil Taxonomy; Science Press: Beijing, China, 2007; ISBN 9787030196538. [Google Scholar]
- Tan, B.; Wu, F.Z.; Yang, W.Q.; Liu, L.; Yu, S. Characteristics of soil animal community in the subalpine/alpine forests of western Sichuan during onset of freezing. Acta Ecol. Sin. 2010, 30, 93–99. [Google Scholar] [CrossRef]
- Xu, W.; Li, W.; Jiang, P.; Wang, H.; Bai, E. Distinct temperature sensitivity of soil carbon decomposition in forest organic layer and mineral soil. Sci. Rep. 2014, 4, 6512. [Google Scholar] [CrossRef] [PubMed]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. PNAS 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microbiol. 1997, 63, 4704–4712. [Google Scholar] [CrossRef] [PubMed]
- Binkley, D.; Hart, S.C. The Components of Nitrogen Availability Assessments in Forest Soils. In Advances in Soil Science, 1st ed.; Stewart, B.A., Ed.; Springer: New York, NY, USA, 1989; pp. 57–112. ISBN 978-1461387732. [Google Scholar]
- Levy-Booth, D.J.; Prescott, C.E.; Grayston, S.J. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 2014, 75, 11–25. [Google Scholar] [CrossRef]
- Mellander, P.E.; Löfvenius, M.O.; Laudon, H. Climate change impact on snow and soil temperature in boreal Scots pine stands. Clim. Chang. 2007, 85, 179–193. [Google Scholar] [CrossRef]
- Pederson, G.T.; Gray, S.T.; Woodhouse, C.A.; Betancourt, J.L.; Fagre, D.B.; Littell, J.S.; Emma, W.; Luckman, B.H.; Graumlich, L.J. The unusual nature of recent snowpack declines in the North American cordillera. Science 2011, 333, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q. Effects of Forest Gap on Foliar Litter Decomposition in the Subalpine/Alpine Forest of Western Sichuan. Ph.D. Thesis, Sichuan Agricultural University, Chengdu, China, 2014. Available online: http://cdmd.cnki.com.cn/Article/CDMD-10626-1016050649.htm (accessed on 3 June 2014).
- Freppaz, M.; Williams, M.W.; Seastedt, T.; Filippa, G. Response of soil organic and inorganic nutrients in alpine soil to a 16-year factorial snow and N-fertilization experiment, Colorado Front Range, USA. Appl. Soil Ecol. 2012, 62, 131–141. [Google Scholar] [CrossRef]
- Yin, R.; Xu, Z.F.; Wu, F.Z.; Yang, W.Q.; Li, Z.P.; Xiong, L.; Xiao, S.; Wang, B. Effects of snow pack on wintertime soil nitrogen transformation in two subalpine forests of western Sichuan. Acta Ecol. Sin. 2014, 34, 2061–2067. [Google Scholar] [CrossRef]
- Shen, J.P.; Zhang, L.M.; Di, H.J.; He, J.Z. A review of ammonia-oxidizing bacteria and archaea in Chinese soils. Front. Microbiol. 2012, 3, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Erguder, T.H.; Boon, N.; Wittebolle, L.; Marzorati, M.; Verstraete, W. Environmental factors shaping the ecological niches of ammonia-oxidizing archaea. FEMS Microbiol. Rev. 2009, 33, 855–869. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.A.; McCulloch, J.; Peter Kershaw, G.; Jefferies, R.L. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring. Soil Biol. Biochem. 2006, 38, 2843–2851. [Google Scholar] [CrossRef]
- Walker, V.K.; Palmer, G.R.; Voordouw, G. Freeze-Thaw Tolerance and Clues to the Winter Survival of a Soil Community. Appl. Environ. Microbiol. 2006, 72, 1784–1792. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.L.; Walker, V.K. Selection of low-temperature resistance in bacteria and potential applications. Environ. Technol. 2010, 31, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Drotz, S.H.; Sparrman, T.; Nilsson, M.B.; Schleucher, J.; Öquist, M.G. Both catabolic and anabolic heterotrophic microbial activity proceed in frozen soils. PNAS 2010, 107, 21046–21051. [Google Scholar] [CrossRef] [PubMed]
- Rasche, F.; Knapp, D.; Kaiser, C.; Koranda, M.; Kitzler, B.; Zechmeister-Boltenstern, S.; Richter, A.; Sessitsch, A. Seasonality and resource availability control bacterial and archaeal communities in soils of a temperate beech forest. ISME J. 2011, 5, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Li, M.; Hong, Y.; Gu, J.D. Diversity and abundance of ammonina-oxidizing archaea and bacteria in polluted mangrove sediment. Syst. Appl. Microbiol. 2011, 34, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R. Cold-adapted archaea. Nat. Rev. Microbiol. 2006, 14, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Wu, F.; Yang, W.; Yu, S.; Liu, L.; Wang, A. The dynamics pattern of soil carbon and nutrients as soil thawing proceeded in the alpine/subalpine forest. Acta Agric. Scand. B-SP 2011, 61, 670–679. [Google Scholar] [CrossRef]
- Regan, K.; Stempfhuber, B.; Schloter, M.; Rasche, F.; Prati, D.; Philippot, L.; Boeddinghaus, R.S.; Kandeler, E.; Marhan, S. Spatial and temporal dynamics of nitrogen fixing, nitrifying and denitrifying microbes in an unfertilized grassland soil. Soil Biol. Biochem. 2017, 109, 214–226. [Google Scholar] [CrossRef]
- Prosser, J.I.; Nicol, G.W. Archaeal and bacterial ammonia-oxidisers in soil: The quest for niche specialization and differentiation. Trends Microbiol. 2012, 20, 523–531. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Carbon (g∙kg−1) | Total Nitrogen (g∙kg−1) | (mg∙kg−1) | (mg∙kg−1) | Bulk Density (g∙cm−3) | pH | |
|---|---|---|---|---|---|---|
| Organic soil | 138.56 ± 4.04 | 7.28 ± 0.07 | 18.73 ± 0.36 | 140.75 ± 2.73 | 1.09 ± 0.05 | 5.6 ± 0.3 |
| Mineral soil | 25.03 ± 0.88 | 1.69 ± 0.03 | 11.32 ± 1.35 | 14.04 ± 1.45 | 1.2 ± 0.03 | 5.3 ± 0.2 |
| Altitude | Abundance of AOB | Abundance of AOA | |||
|---|---|---|---|---|---|
| 3582 m | Abundance of AOB | 1 | 0.632 * | 0.251 | 0.445 |
| Abundance of AOA | 1 | −0.323 | −0.163 | ||
| 1 | 0.478 | ||||
| 1 | |||||
| 3298 m | Abundance of AOB | 1 | 0.777 ** | 0.389 | 0.486 |
| Abundance of AOA | −0.093 | 0.076 | |||
| 0.741 * | |||||
| 1 | |||||
| 3032 m | Abundance of AOB | 1 | −0.060 | 0.677 * | 0.392 |
| Abundance of AOA | −0.493 | −0.772 ** | |||
| 0.420 | |||||
| 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Wang, A.; Wu, F.; Xu, Z.; Tan, B.; Liu, Y.; Yang, Y.; Chen, L.; Yang, W. Soil Nitrogen Responses to Soil Core Transplanting Along an Altitudinal Gradient in an Eastern Tibetan Forest. Forests 2018, 9, 239. https://doi.org/10.3390/f9050239
Zhang L, Wang A, Wu F, Xu Z, Tan B, Liu Y, Yang Y, Chen L, Yang W. Soil Nitrogen Responses to Soil Core Transplanting Along an Altitudinal Gradient in an Eastern Tibetan Forest. Forests. 2018; 9(5):239. https://doi.org/10.3390/f9050239
Chicago/Turabian StyleZhang, Li, Ao Wang, Fuzhong Wu, Zhenfeng Xu, Bo Tan, Yang Liu, Yulian Yang, Lianghua Chen, and Wanqin Yang. 2018. "Soil Nitrogen Responses to Soil Core Transplanting Along an Altitudinal Gradient in an Eastern Tibetan Forest" Forests 9, no. 5: 239. https://doi.org/10.3390/f9050239
APA StyleZhang, L., Wang, A., Wu, F., Xu, Z., Tan, B., Liu, Y., Yang, Y., Chen, L., & Yang, W. (2018). Soil Nitrogen Responses to Soil Core Transplanting Along an Altitudinal Gradient in an Eastern Tibetan Forest. Forests, 9(5), 239. https://doi.org/10.3390/f9050239