The Evaluation of Radiation Use Efficiency and Leaf Area Index Development for the Estimation of Biomass Accumulation in Short Rotation Poplar and Annual Field Crops

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Treatments
2.2. Field Measurements and Data Collection
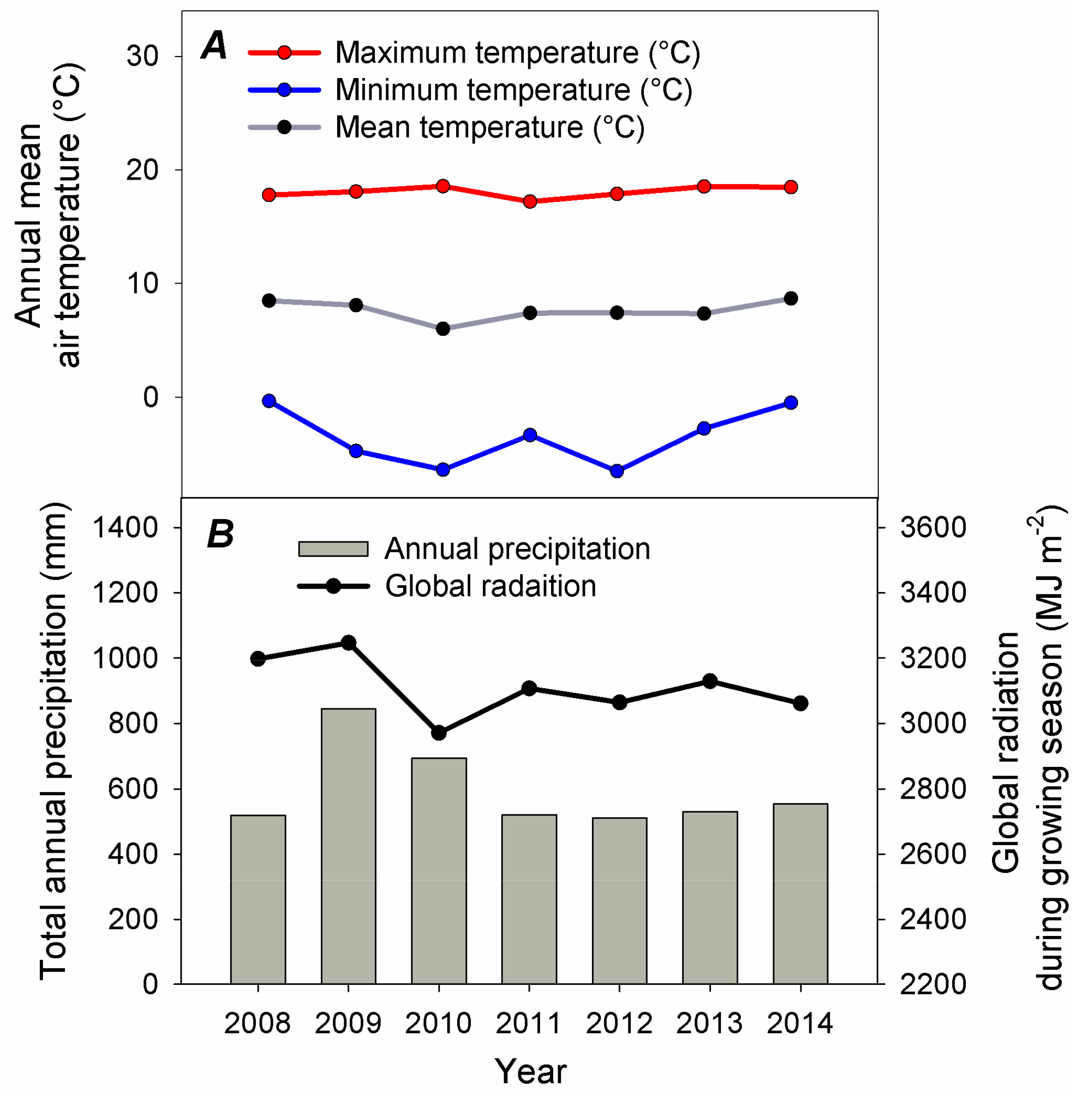
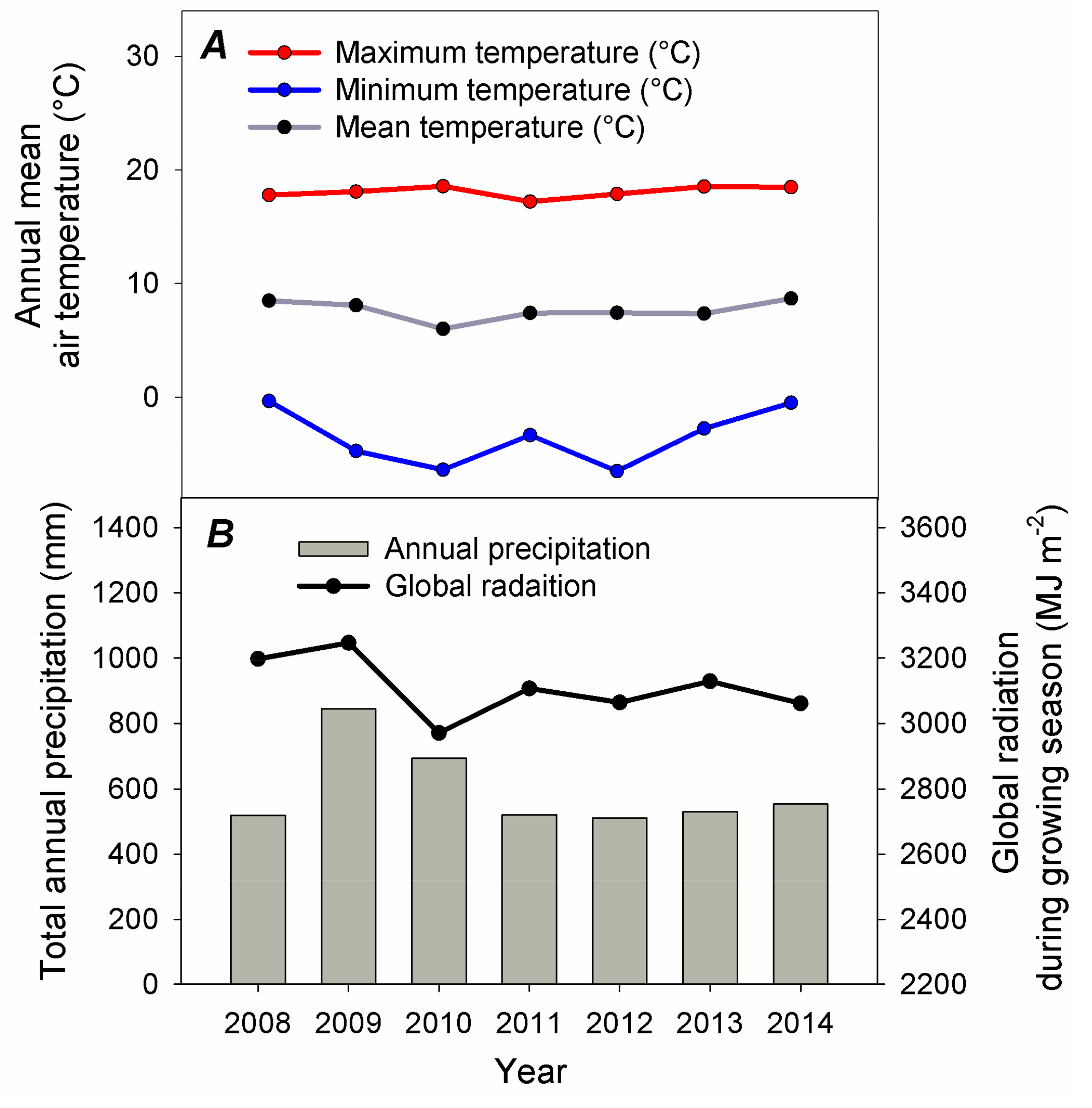
2.3. Meteorological Measurements
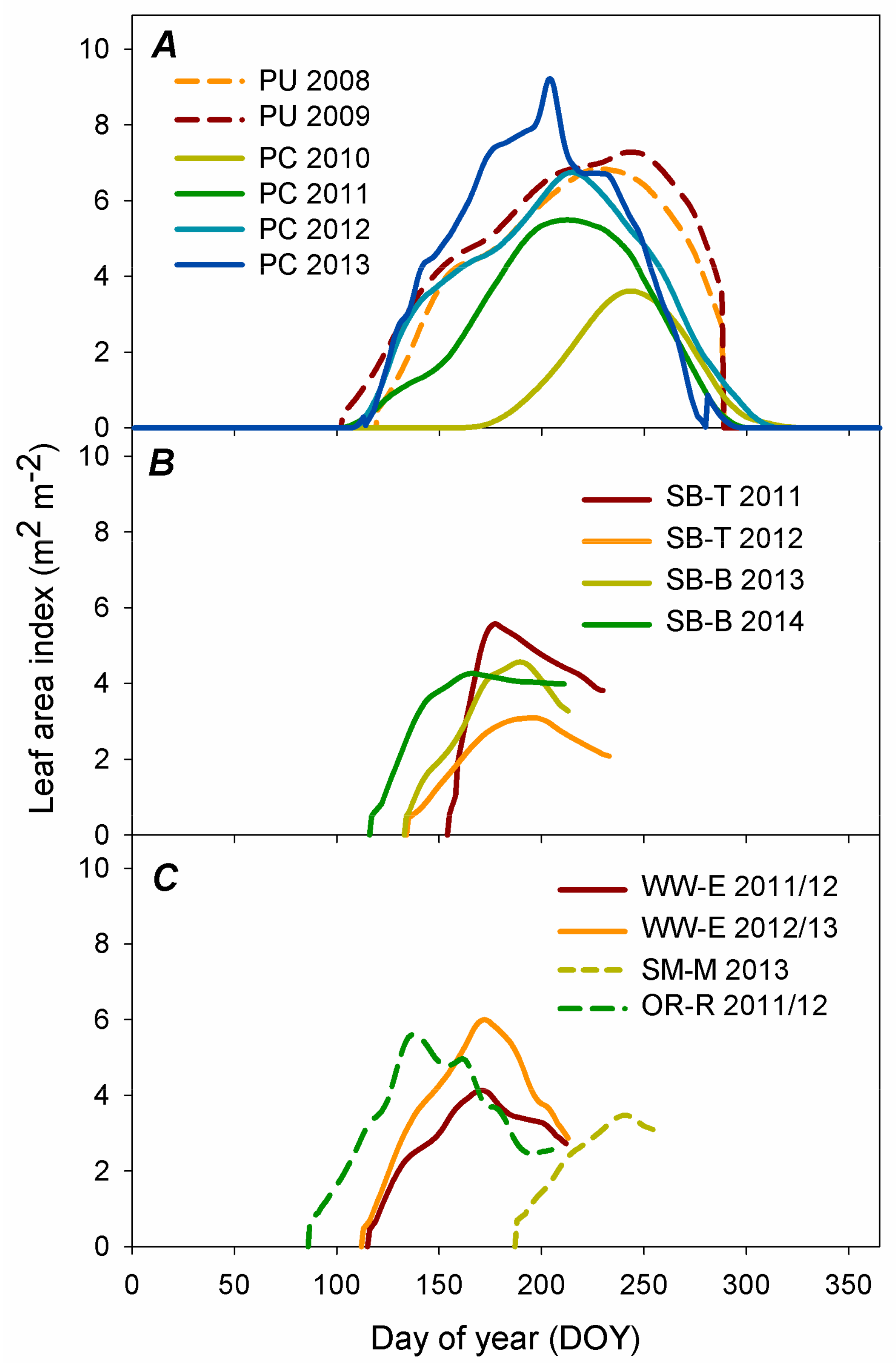
2.4. Leaf Area Development Measurements
2.5. Aboveground Dry Mass (AGDM) Production Estimation
2.6. The Evaluation of Intercepted Photosynthetic Active Radiation (IPAR) and Radiation Use Efficiency (RUE)
2.7. Statistical Analysis
3. Results
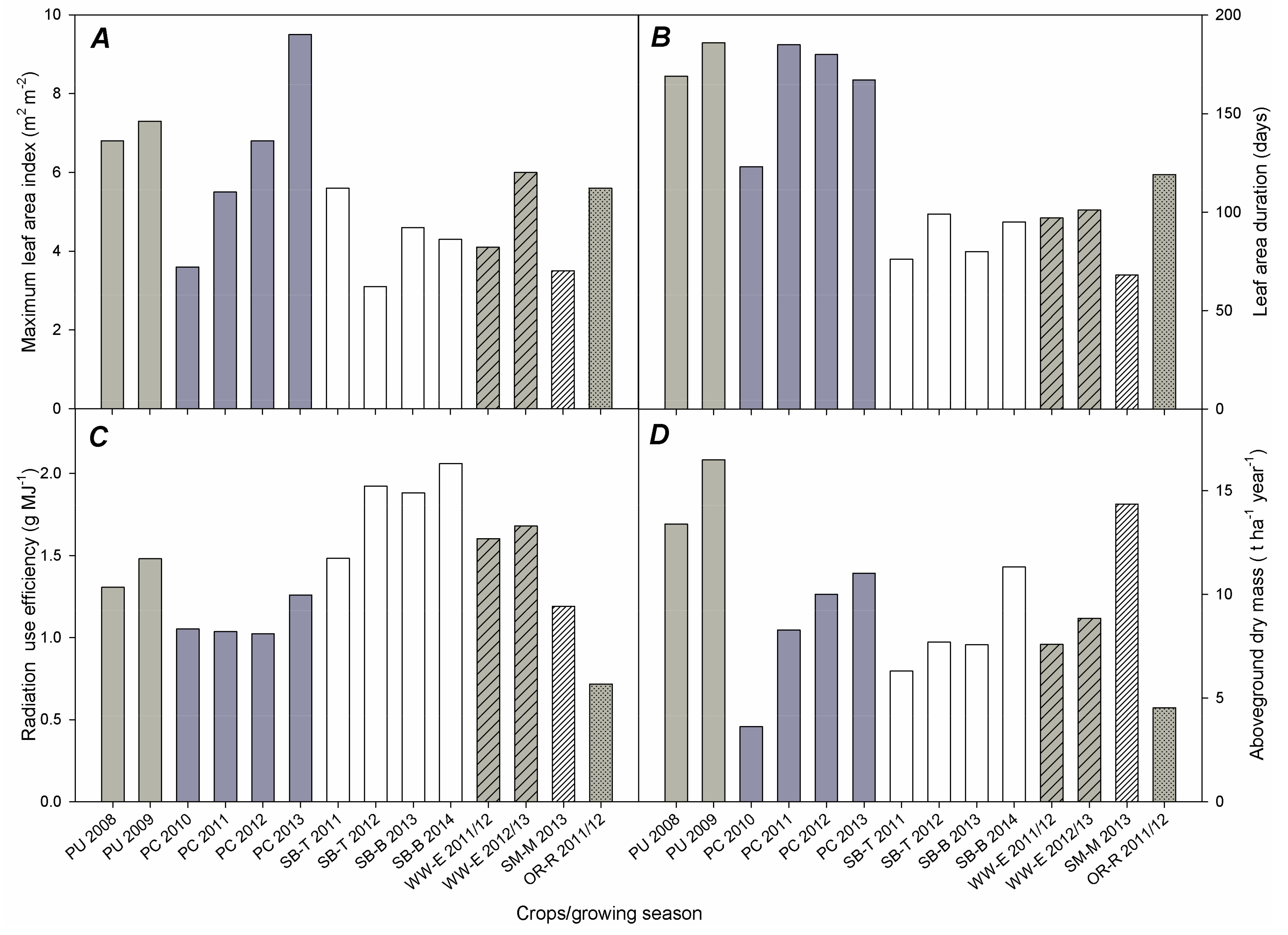
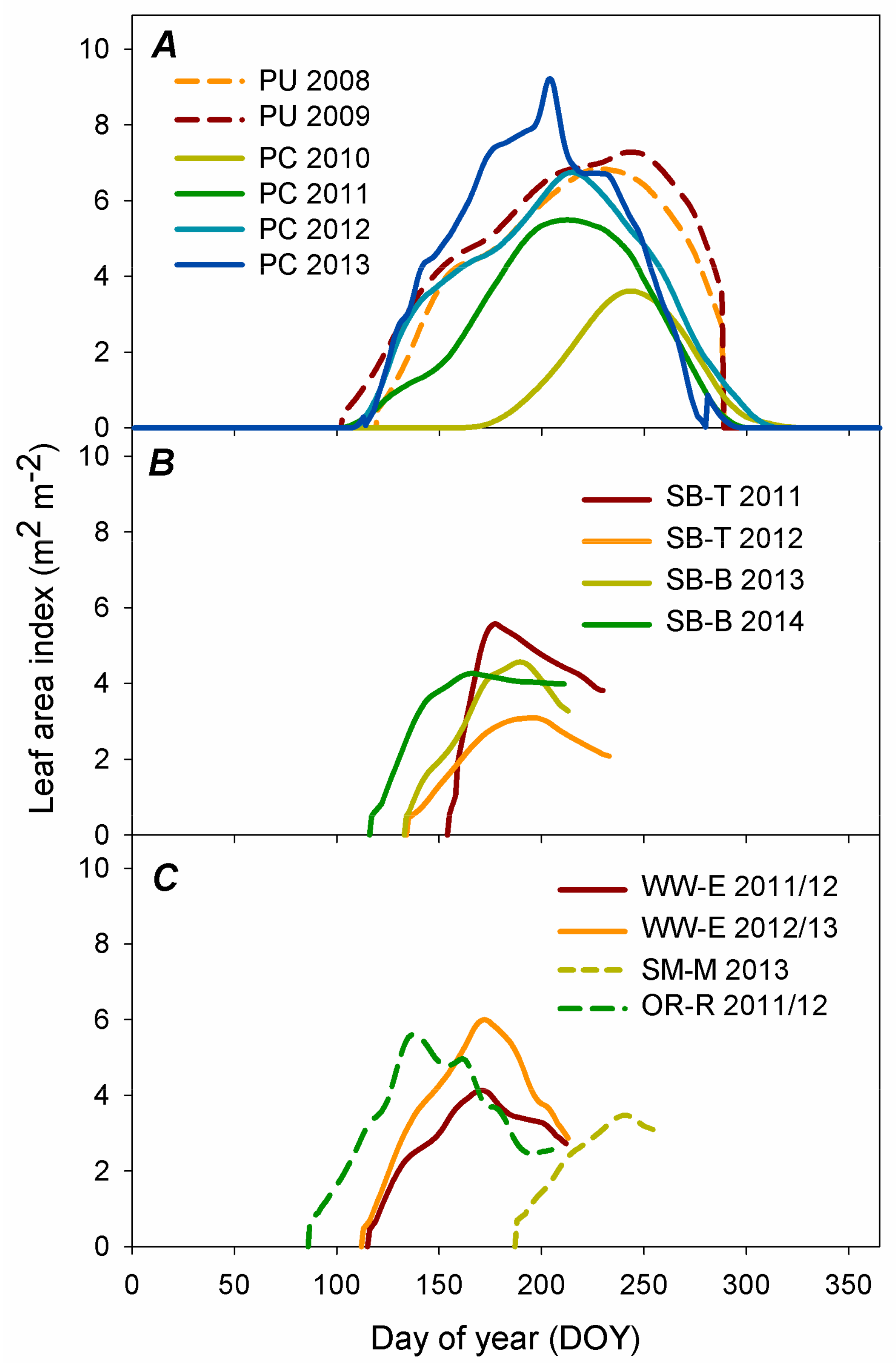
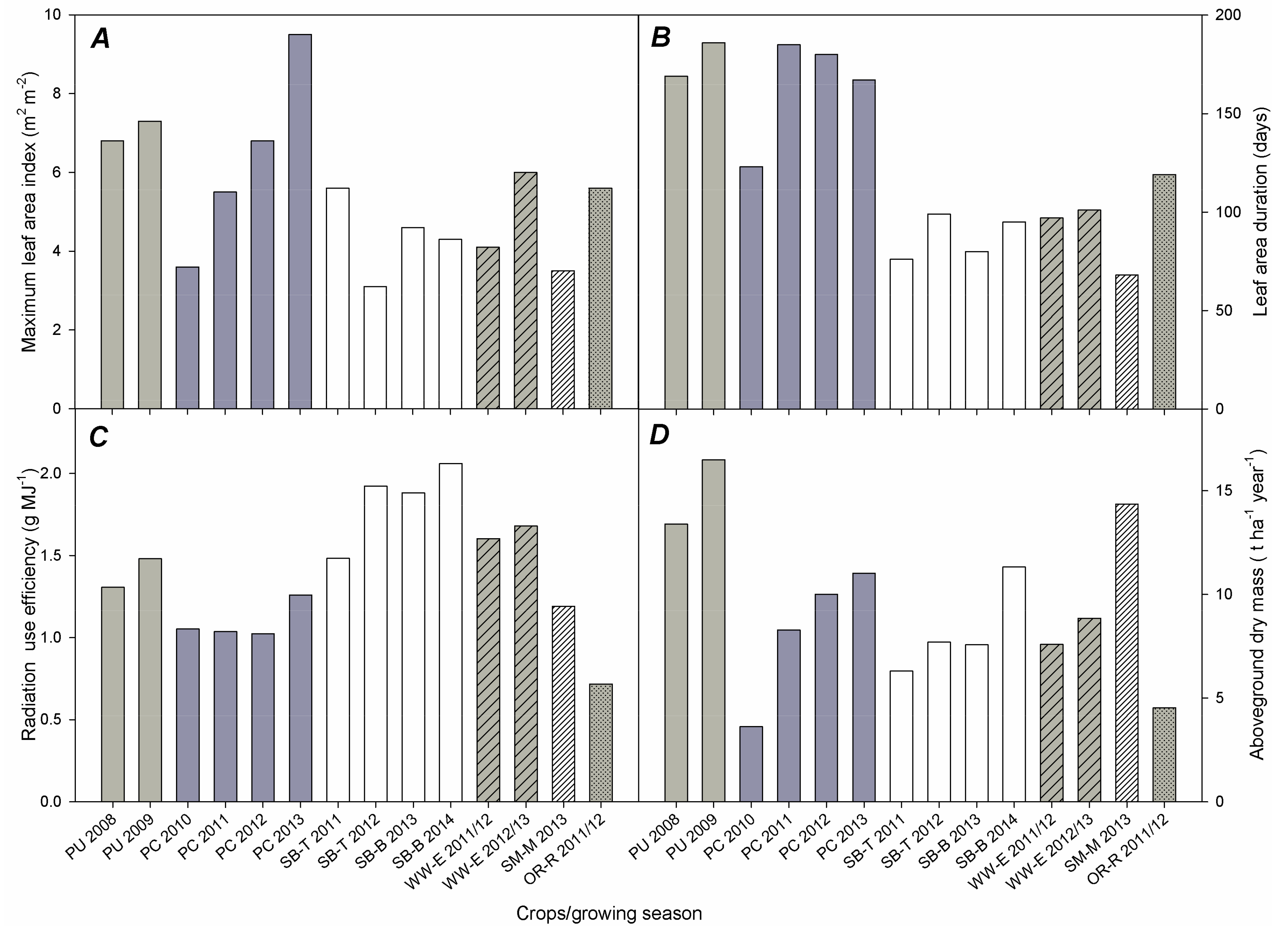
3.1. Leaf Area Development
3.2. Radiation Use Efficiency (RUE)
3.3. Aboveground Dry Mass (AGDM) Production
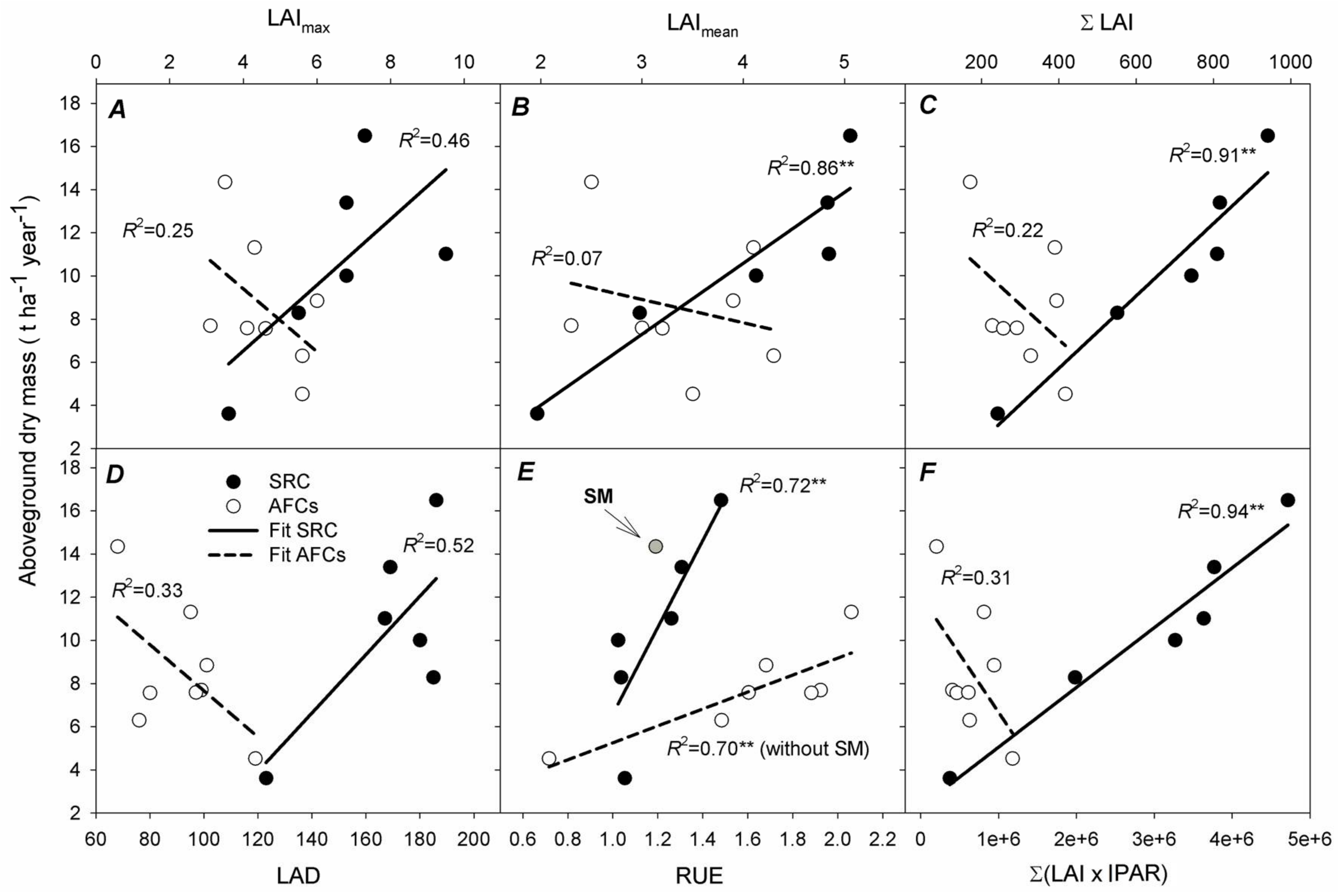
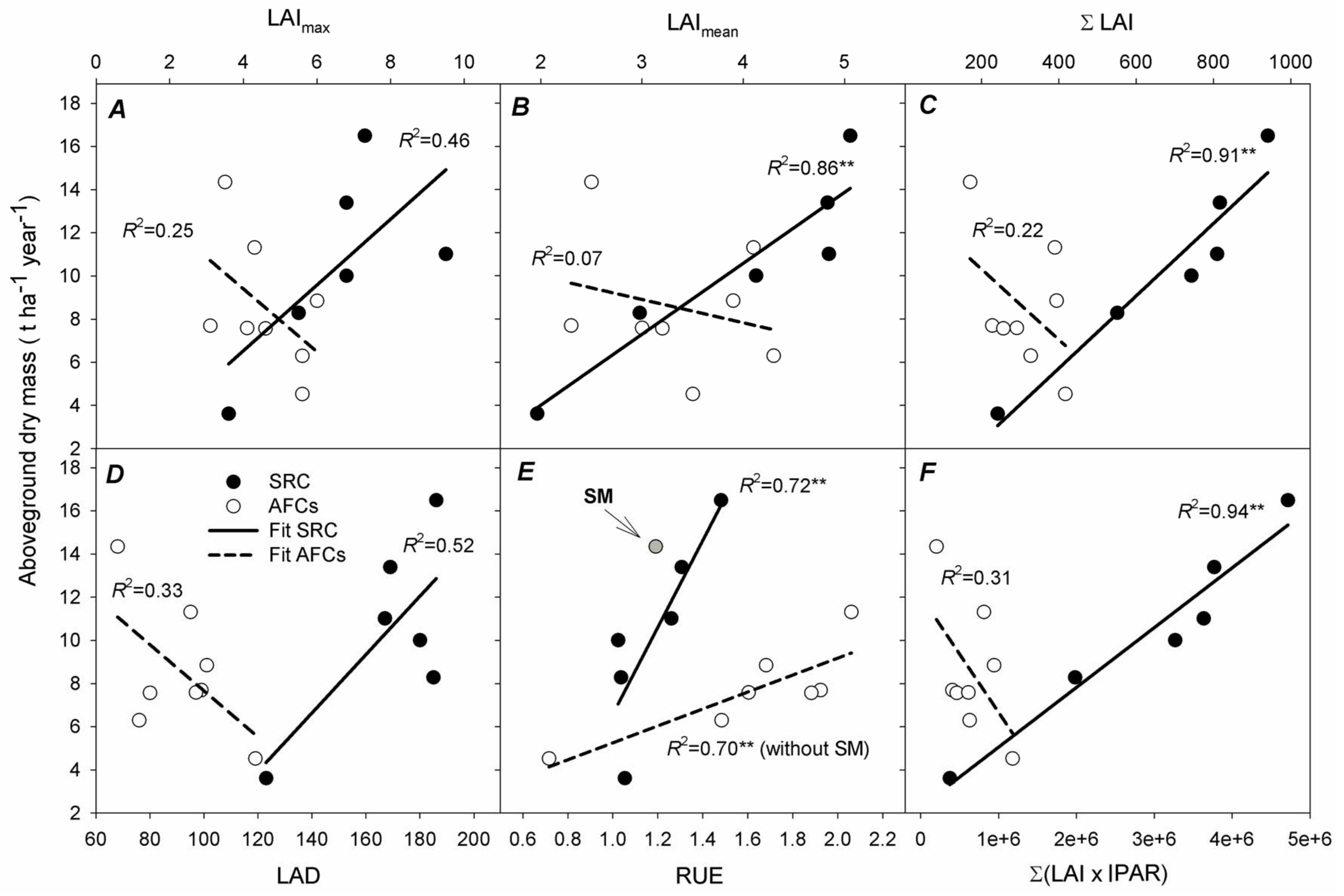
3.4. Determinants of Aboveground Dry Mass (AGDM) Production in SRC Poplar and AFCs
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ∑LAI | Sum of leaf area index |
| AFCs | Annual field crops |
| AGDM | Aboveground dry mass |
| DBH | Diameter at breast height (mm) |
| DM | Dry mass |
| IPAR | Intercepted photosynthetically active radiation (MJ m−2) |
| LAD | Leaf area duration (days) |
| LAI | Leaf area index (m2 m−2) |
| LAImax | Maximum leaf area index |
| LAImean | Mean leaf area index |
| OR | Oilseed rape |
| OR-R | Oilseed rape cultivar Rohan |
| PAR | Photosynthetically active radiation |
| PC | Poplar coppiced |
| PU | Poplar uncoppiced |
| RUE | Radiation use efficiency (g MJ−1) |
| SB | Spring barley |
| SB-B | Spring barley cultivar Bojos |
| SB-T | Spring barley cultivar Tolar |
| SM | Silage maize |
| SM-M | Silage maize cultivar MON3301 |
| SRC | Short rotation coppice |
| WW | Winter wheat |
| WW-E | Winter wheat cultivar Etela |
References
- IPCC (Intergovernmental Panel on Climate Change). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; ISBN 9789291691432. [Google Scholar]
- Pinstrup-Andersen, P. Food security: Definition and measurement. Food Secur. 2009, 1, 5–7. [Google Scholar] [CrossRef]
- Popp, J.; Lakner, Z.; Harangi-Rákos, M.; Fári, M. The effect of bioenergy expansion: Food, energy, and environment. Renew. Sustain. Energy Rev. 2014, 32, 559–578. [Google Scholar] [CrossRef]
- Scarlat, N.; Dallemand, J.F. Recent developments of biofuels/bioenergy sustainability certification: A global overview. Energy Policy 2011, 39, 1630–1646. [Google Scholar] [CrossRef]
- Busetto, L.; Casteleyn, S.; Granell, C.; Pepe, M.; Barbieri, M.; Campos-Taberner, M.; Casa, R.; Collivignarelli, F.; Confalonieri, R.; Crema, A.; et al. Downstream services for rice crop monitoring in Europe: From regional to local scale. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2017, 10, 5423–5441. [Google Scholar] [CrossRef]
- Dimitriou, I.; Baum, C.; Baum, S.; Busch, G.; Schulz, U.; Köhn, J.; Lamersdorf, N.; Leinweber, P.; Aronsson, P.; Weih, M.; et al. Quantifying Environmental Effects of Short Rotation Coppice (SRC) on Biodiversity, Soil and Water; IEA Bioenergy Task 43, Report 2011; IEA: Paris, France, 2011; p. 34. [Google Scholar]
- Kauter, D.; Lewandowski, I.; Claupein, W. Quantity and quality of harvestable biomass from Populus short rotation coppice for solid fuel use—A review of the physiological basis and management influences. Biomass Bioenergy 2003, 24, 411–427. [Google Scholar] [CrossRef]
- Timmer, C.P. The macro dimensions of food security: Economic growth, equitable distribution, and food price stability. Food Policy 2000, 25, 283–295. [Google Scholar] [CrossRef]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Ewert, F. Modelling plant responses to elevated CO2: How important is leaf area index? Ann. Bot. 2004, 93, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Black, T.A. Defining leaf area index for non-flat leaves. Plant Cell Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Monteith, J.L. Reassessment of maximum growth rates for C3 and C4 Crops. Exp. Agric. 1978, 14, 1–5. [Google Scholar] [CrossRef]
- Biscoe, P.V.; Gallagher, J.N. Weather, dry matter production and yield. In Environmental Effects on Crop Physiology; Proceedings of the a Symposium Held at Long Ashton Research Station, University of Bristol, Bristol, UK, 13–16 April 1975; Academic Press: New York, NY, USA, 1977. [Google Scholar]
- Hangs, R.D.; Van Rees, K.C.J.; Schoenau, J.J.; Guo, X. A simple technique for estimating above-ground biomass in short-rotation willow plantations. Biomass Bioenergy 2011, 35, 2156–2162. [Google Scholar] [CrossRef]
- Hikosaka, K. Leaf canopy as a dynamic system: Ecophysiology and optimality in leaf turnover. Ann. Bot. 2005, 95, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Horie, T. Leaf nitrogen, photosynthesis, and crop radiation use efficiency: A review. Crop Sci. 1989, 29, 90. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Shiraiwa, T.; Hammer, G.L. Variation in crop radiation-use efficiency with increased diffuse radiation. Crop Sci. 1992, 32, 1281–1284. [Google Scholar] [CrossRef]
- Medlyn, B.E. Physiological basis of the light use efficiency model. Tree Physiol. 1998, 18, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yan, J.; Li, J.; Sang, T. Yield potential of Miscanthus energy crops in the Loess Plateau of China. GCB Bioenergy 2012, 4, 545–554. [Google Scholar] [CrossRef]
- Stockle, C.O.; Kiniry, J.R. Variability in crop radiation-use efficiency associated with vapor-pressure deficit. Field Crop. Res. 1990, 25, 171–181. [Google Scholar] [CrossRef]
- Larson, P.R.; Isebrands, J.G. The relation between leaf production and wood weight in first-year root sprouts of two Populus clones. Can. J. For. Res. 1972, 2, 98–104. [Google Scholar] [CrossRef]
- Bartelink, H.H. A growth model for mixed forest stands. For. Ecol. Manag. 2000, 134, 29–43. [Google Scholar] [CrossRef]
- Bartelink, H.H.; Kramer, K.; Mohren, G.M.J. Applicability of the radiation-use efficiency concept for simulating growth of forest stands. Agric. For. Meteorol. 1997, 88, 169–179. [Google Scholar] [CrossRef]
- Kemanian, A.R.; Stöckle, C.O.; Huggins, D.R. Variability of barley radiation-use efficiency. Crop Sci. 2004, 44, 1662–1672. [Google Scholar] [CrossRef]
- Jamieson, P.D.; Martin, R.J.; Francis, G.S.; Wilson, D.R. Drought effects on biomass production and radiation-use efficiency in barley. Field Crop. Res. 1995, 43, 77–86. [Google Scholar] [CrossRef]
- Zahoor, A.; Riaz, M.; Ahmad, S.; Ali, H.; Khan, M.B.; Javed, K.; Anjum, M.A.; Zia-Ul-Haq, M.; Khan, M.A. Ontogeny growth and radiation use efficiency of Helianthus annuus L., as affected by hybrids, nitrogenous regimes and planting geometry under irrigated arid conditions. Pak. J. Bot. 2010, 42, 3197–3207. [Google Scholar]
- Ceulemans, R.; McDonald, A.J.S.; Pereira, J.S. A comparison among eucalypt, poplar and willow characteristics with particular reference to a coppice, growth-modelling approach. Biomass Bioenergy 1996, 11, 215–231. [Google Scholar] [CrossRef]
- Broeckx, L.; Vanbeveren, S.; Verlinden, M.; Ceulemans, R. First vs. second rotation of a poplar short rotation coppice: Leaf area development, light interception and radiation use efficiency. iForest-Biogeosci. For. 2015, e1–e9. [Google Scholar] [CrossRef]
- Kiniry, J.R. Biomass accumulation and radiation use efficiency of honey mesquite and eastern red cedar. Biomass Bioenergy 1998, 15, 467–473. [Google Scholar] [CrossRef]
- Kiniry, J.R.; Jones, C.A.; O’toole, J.C.; Blanchet, R.; Cabelguenne, M.; Spanel, D.A. Radiation-use efficiency in biomass accumulation prior to grain-filling for five grain-crop species. Field Crop. Res. 1989, 20, 51–64. [Google Scholar] [CrossRef]
- Landsberg, J.J.; Wright, L.L. Comparisons among Populus clones and intensive culture conditions, using an energy-conservation model. For. Ecol. Manag. 1989, 27, 129–147. [Google Scholar] [CrossRef]
- Iio, A.; Hikosaka, K.; Anten, N.P.R.; Nakagawa, Y.; Ito, A. Global dependence of field-observed leaf area index in woody species on climate: A systematic review. Glob. Ecol. Biogeogr. 2014, 23, 274–285. [Google Scholar] [CrossRef]
- Wright, I.J.; Groom, P.K.; Lamont, B.B.; Poot, P.; Prior, L.D.; Reich, P.B.; Schulze, E.D.; Veneklaas, E.J.; Westoby, M. Leaf trait relationships in Australian plant species. Funct. Plant Biol. 2004, 31, 551–558. [Google Scholar] [CrossRef]
- Asner, G.P.; Scurlock, J.M.O.; Hicke, J.A. Global synthesis of leaf area index observations. Glob. Ecol. Biogeogr. 2003, 12, 191–205. [Google Scholar] [CrossRef]
- Harrington, R.A.; Fownes, J.H.; Meinzer, F.C.; Scowcroft, P.G. Forest growth along a rainfall gradient in Hawaii: Acacia koa stand structure, productivity, foliar nutrients, and water- and nutrient-use efficiencies. Oecologia 1995, 102, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Cannell, M.G.R.; Sheppard, L.J.; Milne, R. Light use efficiency and woody biomass production of Poplar and Willow. Forestry 1988, 61, 125–136. [Google Scholar] [CrossRef]
- Haxeltine, A.; Prentice, I.C. BIOME 3: An equilibrium terrestrial biosphere model based on ecophysiological constraints, resource availability and competition among plant functional types. Glob. Biogeochem. Cycles 1996, 10, 693–709. [Google Scholar] [CrossRef]
- Choudhury, B.J. Modeling radiation- and carbon-use efficiencies of maize, sorghum, and rice. Agric. For. Meteorol. 2001, 106, 317–330. [Google Scholar] [CrossRef]
- Wolf, J.; Evans, L.G.; Semenov, M.A.; Eckersten, H.; Iglesias, A. Comparison of wheat simulation models under climate change. I. Model calibration and sensitivity analyses. Clim. Res. 1996, 7, 253–270. [Google Scholar] [CrossRef]
- Green, D.S.; Kruger, E.L.; Stanosz, G.R.; Isebrands, J.G. Light-use efficiency of native and hybrid poplar genotypes at high levels of intracanopy competition. Can. J. For. Res. 2001, 31, 1030–1037. [Google Scholar] [CrossRef]
- Trnka, M.; Trnka, M.; Fialová, J.; Koutecký, V.; Fajman, M.; Žalud, Z.; Hejduk, S. Biomass production and survival rates of selected poplar clones grown under a short-rotation system on arable land. Plant Soil Environ. 2008, 54, 78–88. [Google Scholar] [CrossRef]
- Fischer, M.; Orság, M.; Trnka, M.; Pohanková, E.; Hlavinka, P.; Tripathi, A.M.; Žalud, Z. Annual and intra-annual water balance components of a short rotation poplar coppice based on sap flow and micrometeorological and hydrological approaches. Acta Hortic. 2013. [Google Scholar] [CrossRef]
- Pohanková, E.; Hlavinka, P.; Takáč, J.; Žalud, Z.; Trnka, M. Calibration and validation of the crop growth model daisy for spring barley in the Czech Republic. Acta Univ. Agric. Silvic. Mendel. Brun. 2015, 63. [Google Scholar] [CrossRef]
- Bonhomme, R. Beware of comparing RUE values calculated from PAR vs solar radiation or absorbed vs intercepted radiation. Field Crop. Res. 2000, 68, 247–252. [Google Scholar] [CrossRef]
- Bréda, N.J.J. Ground-based measurements of leaf area index: A review of methods, instruments and current controversies. J. Exp. Bot. 2003, 54, 2403–2417. [Google Scholar] [CrossRef] [PubMed]
- Campos-Taberner, M.; García-Haro, F.J.; Confalonieri, R.; Martínez, B.; Moreno, Á.; Sánchez-Ruiz, S.; Gilabert, M.A.; Camacho, F.; Boschetti, M.; Busetto, L. Multitemporal monitoring of plant area index in the Valencia rice district with PocketLAI. Remote Sens. 2016, 8. [Google Scholar] [CrossRef]
- Tripathi, A.M.; Fischer, M.; Orság, M.; Marek, M.V.; Žalud, Z.; Trnka, M. Evaluation of indirect measurement method of seasonal patterns of leaf area index in a high-density short rotation coppice culture of poplar. Acta Univ. Agric. Silvic. Mendel. Brun. 2016, 64, 549–556. [Google Scholar] [CrossRef]
- Liu, G.; Xie, Y.; Gao, X.F.; Duan, X.W. Application of SunScan canopy analysis system in measuring leaf area index of soybean. Chin. J. Ecol. 2008, 27, 862–866. [Google Scholar]
- Oguntunde, P.G.; Olukunle, O.J.; Fasinmirin, J.T.; Abiolu, O.A. Performance of the SunScan canopy analysis system in estimating leaf area index of maize. Agric. Eng. Int. CIGR J. 2012, 14, 1–7. [Google Scholar]
- Wilhelm, W.W.; Ruwe, K.; Schlemmer, M.R. Comparison of three leaf area index meters in a corn canopy. Crop Sci. 2000, 40, 1179–1183. [Google Scholar] [CrossRef]
- Schirrmann, M.; Hamdorf, A.; Giebel, A.; Dammer, K.H.; Garz, A. A mobile sensor for leaf area index estimation from canopy light transmittance in wheat crops. Biosyst. Eng. 2015, 140, 23–33. [Google Scholar] [CrossRef]
- Linderson, M.L.; Iritz, Z.; Lindroth, A. The effect of water availability on stand-level productivity, transpiration, water use efficiency and radiation use efficiency of field-grown willow clones. Biomass Bioenergy 2007, 31, 460–468. [Google Scholar] [CrossRef]
- Fischer, M.; Trnka, M.; Kučera, J.; Fajman, M.; Žalud, Z. Biomass productivity and water use relation in short rotation poplar coppice (Populus nigra × P. maximowiczii) in the conditions of Czech Moravian Highlands. Acta Univ. Agric. Silvic. Mendel. Brun. 2011, 59, 141–152. [Google Scholar] [CrossRef]
- Ketterings, Q.M.; Coe, R.; Van Noordwijk, M.; Ambagau, Y.; Palm, C.A. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For. Ecol. Manag. 2001, 146, 199–209. [Google Scholar] [CrossRef]
- Basuki, T.M.; van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. Uber den Lichtfaktor in den Pflanzengesellschaften und seine Bedeutung fur die Stoffproduktion. Jpn. J. Bot. 1953, 14, 22–52. [Google Scholar]
- Kemanian, A.R.; Stöckle, C.O.; Huggins, D.R. Transpiration-use efficiency of barley. Agric. For. Meteorol. 2005, 130, 1–11. [Google Scholar] [CrossRef]
- Heidari, H.; Johansooz, M.R.; Hosseini, S.M.B.; Chaichi, M.R. Alternate furrow irrigation effect on radiation use efficiency and forage quality of foxtail millet (Setaria italica). Ann. Biol. Res. 2012, 3, 2565–2574. [Google Scholar]
- Ajit; Das, D.K.; Chaturvedi, O.P.; Jabeen, N.; Dhyani, S.K. Predictive models for dry weight estimation of above and below ground biomass components of Populus deltoides in India: Development and comparative diagnosis. Biomass Bioenergy 2011, 35, 1145–1152. [Google Scholar] [CrossRef]
- Howe, G.T.; Saruul, P.; Davis, J.; Chen, T.H.H. Quantitative genetics of bud phenology, frost damage, and winter survival in an F2 family of hybrid poplars. Theor. Appl. Genet. 2000, 101, 632–642. [Google Scholar] [CrossRef]
- Tharakan, P.J.; Volk, T.A.; Nowak, C.A.; Ofezu, G.J. Assessment of canopy structure, light interception, and light-use efficiency of first year regrowth of shrub willow (Salix sp.). BioEnergy Res. 2008, 1, 229–238. [Google Scholar] [CrossRef]
- Ceulemans, R.; Stettler, R.F.; Hinckley, T.M.; Isebrands, J.G.; Heilman, P.E. Crown architecture of Populus clones as determined by branch orientation and branch characteristics. Tree Physiol. 1990, 7, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Rae, A.M.; Robinson, K.M.; Street, N.R.; Taylor, G. Morphological and physiological traits influencing biomass productivity in short-rotation coppice poplar. Can. J. For. Res. 2004, 34, 1488–1498. [Google Scholar] [CrossRef]
- Ahmad, S.; Ali, H.; Ismail, M.; Shahzad, M.I.; Nadeem, M.; Anjum, M.A.; Zia-Ul-Haq, M.; Firdous, N.; Khan, M.A. Radiation and nitrogen use efficiencies of C3 winter cereals to nitrogen split application. Pak. J. Bot. 2012, 44, 139–149. [Google Scholar]
- Choudhury, B.J. A sensitivity analysis of the radiation use efficiency for gross photosynthesis and net carbon accumulation by wheat. Agric. For. Meteorol. 2000, 101, 217–234. [Google Scholar] [CrossRef]
- Timlin, D.J.; Fleisher, D.H.; Kemanian, A.R.; Reddy, V.R. Plant density and leaf area index effects on the distribution of light transmittance to the soil surface in maize. Agron. J. 2014, 106, 1828–1837. [Google Scholar] [CrossRef]
- Kuai, J.; Sun, Y.; Zuo, Q.; Huang, H.; Liao, Q.; Wu, C.; Lu, J.; Wu, J.; Zhou, G. The yield of mechanically harvested rapeseed (Brassica napus L.) can be increased by optimum plant density and row spacing. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.J. Comparative physiological studies on the growth of field crops. Ann. Appl. Biol. 1947, 11, 41–76. [Google Scholar] [CrossRef]
- García Del Moral, L.F.; Rharrabti, Y.; Villegas, D.; Royo, C. Evaluation of grain yield and its components in durum wheat under Mediterranean conditions: An ontogenic approach. Agron. J. 2003, 95, 266–274. [Google Scholar] [CrossRef]
- Guttieri, M.J.; Stark, J.C.; O’Brien, K.; Souza, E. Relative sensitivity of spring wheat grain yield and quality parameters to moisture deficit. Crop Sci. 2001, 41, 327–335. [Google Scholar] [CrossRef]
- Richards, R.; Townley-Smith, T. Variation in leaf area development and its effect on water use, yield and harvest index of droughted wheat. Aust. J. Agric. Res. 1987, 38, 983–992. [Google Scholar] [CrossRef]
- Johnson, E. Goodbye to carbon neutral: Getting biomass footprints right. Environ. Impact Assess. Rev. 2009, 29, 165–168. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.; Wang, M.; Xu, J.; Luedeling, E. Responses of spring phenology in temperate zone trees to climate warming: A case study of apricot flowering in China. Agric. For. Meteorol. 2015, 201, 1–7. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, T.; Shao, J.; Guo, S.; Zhu, W.; Yu, Y.J.; Chen, S.L.; Hatano, R. Modeling the biomass of energy crops: Descriptions, strengths and prospective. J. Integr. Agric. 2017, 16, 1197–1210. [Google Scholar] [CrossRef]
- Piedade, M.T.F.; Junk, W.J.; Long, S.P. The productivity of the C4 grass Echinochloa polystachya on the Amazon floodplain. Ecology 1991, 72, 1456–1463. [Google Scholar] [CrossRef]
- Muurinen, S.; Peltonen-Sainio, P. Radiation-use efficiency of modern and old spring cereal cultivars and its response to nitrogen in northern growing conditions. Field Crop. Res. 2006, 96, 363–373. [Google Scholar] [CrossRef]
- Cassman, K.G.; Dobermann, A.; Walters, D.T. Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 2002, 31, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Plenet, D.; Mollier, A.; Pellerin, S. Growth analysis of maize field crops under phosphorus deficiency. II. Radiation-use efficiency, biomass accumulation and yield components. Plant Soil. 2000, 224, 259–272. [Google Scholar] [CrossRef]
- Li, Q.; Liu, M.; Zhang, J.; Dong, B.; Bai, Q. Biomass accumulation and radiation use efficiency of winter wheat under deficit irrigation regimes. Plant Soil Environ. 2009, 2009, 85–91. [Google Scholar] [CrossRef]
- Li, Q.; Chen, Y.; Liu, M.; Zhou, X.; Yu, S.; Dong, B. Effects of irrigation and planting patterns on radiation use efficiency and yield of winter wheat in North China. Agric. Water Manag. 2008, 95, 469–476. [Google Scholar] [CrossRef]
- Goyne, P.J.; Milroy, S.P.; Lilley, J.M.; Hare, J.M. Radiation interception, radiation use efficiency and growth of barley cultivars. Aust. J. Agric. Res. 1993, 44. [Google Scholar] [CrossRef]
- Williams, W.; Loomis, R.; Lepley, C. Vegetative growth of corn as affected by population density. I. Productivity in relation to interception of solar radiation. Crop Sci. 1965, 5, 211–215. [Google Scholar] [CrossRef]
- Kiniry, J.R.; Landivar, J.A.; Witt, M.; Gerik, T.J.; Cavero, J.; Wade, L.J. Radiation-use efficiency response to vapor pressure deficit for maize and sorghum. Field Crop. Res. 1998, 56, 265–270. [Google Scholar] [CrossRef]
- Deckmyn, G.; Laureysens, I.; Garcia, J.; Muys, B.; Ceulemans, R. Poplar growth and yield in short rotation coppice: Model simulations using the process model SECRETS. Biomass Bionergy 2004, 26, 221–227. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Broeckx, L.S.; Van den Bulcke, J.; Van Acker, J.; Ceulemans, R. Comparative study of biomass determinants of 12 poplar (Populus) genotypes in a high-density short-rotation culture. For. Ecol. Manag. 2013, 307, 101–111. [Google Scholar] [CrossRef]
- Bungart, R.; Hüttl, R.F. Growth dynamics and biomass accumulation of 8-year-old hybrid poplar clones in a short-rotation plantation on a clayey-sandy mining substrate with respect to plant nutrition and water budget. Eur. J. For. Res. 2004, 123, 105–115. [Google Scholar] [CrossRef]
- Scaracia-Mugnozza, G.E.; Ceulemans, R.; Heilman, P.E.; Isebrands, J.G.; Stettler, R.F.; Hinckley, T.M. Production physiology and morphology of Populus species and their hybrids grown under short rotation. II. Biomass components and harvest index of hybrid and parental species clones. Can. J. For. Res. 1997, 27, 285–294. [Google Scholar] [CrossRef]
- Dunlap, J.M.; Stettler, R.F. Genetic variation and productivity of Populus trichocarpa and its hybrids. X. Trait correlations in young black cottonwood from four river valleys in Washington. Trees Struct. Funct. 1998, 13, 28–39. [Google Scholar] [CrossRef]
- Heilman, P.E.; Stettler, R.F. Genetic variation and productivity of Populus trichocarpa and its hybrids. IV. Performance in short-rotation coppice. Can. J. For. Res. 1990, 20, 1257–1264. [Google Scholar] [CrossRef]
- Nedomova, L. Adriana, Müller 24—Rape Varieties with Excellent Yield from the BOR, Ltd. A Sbornik z Konference Prosperujici Olejniny; CZU: Prague, Czech Republic, 2012; pp. 120–122. [Google Scholar]
- Diepenbrock, W. Yield analysis of winter oilseed rape (Brassica napus L.): A review. Field Crop. Res. 2000, 67, 35–49. [Google Scholar] [CrossRef]
- Spiertz, J.H.J.; Ellen, J. Effects of nitrogen on crop development and grain growth of winter in relation to assimilate and utilization of assimilates and nutrients. Neth. J. Agric. Sci. 1978, 26, 210–231. [Google Scholar]
- Massad, R.S.; Tuzet, A.; Bethenod, O. The effect of temperature on C4-type leaf photosynthesis parameters. Plant Cell Environ. 2007, 30, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F.; Kubien, D.S. The temperature response of C3 and C4 photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Characteristics | Units | Depth (cm) | |||
|---|---|---|---|---|---|
| 0–24 | 24–66 | 66–94 | 94–130+ | ||
| Silt | wt % | 50 | 46.1 | 38.7 | 19.6 |
| Clay | wt % | 15.8 | 26.3 | 18.6 | 13.3 |
| Bulk density | g/cm3 | 1.55 | 1.64 | 1.59 | 1.64 |
| Organic matter | wt % | 2.65 | 0.28 | 0.14 | 0.14 |
| Total nitrogen | wt % | 0.16 | <0.05 | <0.05 | <0.05 |
| pH (KCl) | 5.9 | 5.4 | 4 | 3.4 | |
| Available P | mg/kg | 148 | 1.3 | 0.9 | 24 |
| Available K | mg/kg | 15 | 91 | 62 | 76 |
| Available Mg | mg/kg | 143 | 230 | 278 | 291 |
| Available Ca | mg/kg | 1230 | 1353 | 748 | 652 |
| Annual Field Crops (AFCs) | Cultivars | Sowing Date | Seeding Rate (kg ha−1) | N Rate (kg ha−1) |
|---|---|---|---|---|
| Spring barley | Tolar | 11 April 2011 | 220 | 60 |
| Spring barley | Tolar | 17 April 2012 | 220 | 70 |
| Spring barley | Bojos | 18 April 2013 | 220 | 30 |
| Spring barley | Bojos | 12 March 2014 | 220 | 30 |
| Winter wheat | Etela | 4 October 2011 | 200 | 15 |
| Winter wheat | Etela | 26 September 2012 | 200 | 15 |
| Silage maize | MON3301 | 10 May 2012 | 35 | 140 |
| Oilseed rape | Rohan | 16 August 2011 | 4.5 | 60 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, A.M.; Pohanková, E.; Fischer, M.; Orság, M.; Trnka, M.; Klem, K.; Marek, M.V. The Evaluation of Radiation Use Efficiency and Leaf Area Index Development for the Estimation of Biomass Accumulation in Short Rotation Poplar and Annual Field Crops. Forests 2018, 9, 168. https://doi.org/10.3390/f9040168
Tripathi AM, Pohanková E, Fischer M, Orság M, Trnka M, Klem K, Marek MV. The Evaluation of Radiation Use Efficiency and Leaf Area Index Development for the Estimation of Biomass Accumulation in Short Rotation Poplar and Annual Field Crops. Forests. 2018; 9(4):168. https://doi.org/10.3390/f9040168
Chicago/Turabian StyleTripathi, Abhishek Mani, Eva Pohanková, Milan Fischer, Matěj Orság, Miroslav Trnka, Karel Klem, and Michal V. Marek. 2018. "The Evaluation of Radiation Use Efficiency and Leaf Area Index Development for the Estimation of Biomass Accumulation in Short Rotation Poplar and Annual Field Crops" Forests 9, no. 4: 168. https://doi.org/10.3390/f9040168
APA StyleTripathi, A. M., Pohanková, E., Fischer, M., Orság, M., Trnka, M., Klem, K., & Marek, M. V. (2018). The Evaluation of Radiation Use Efficiency and Leaf Area Index Development for the Estimation of Biomass Accumulation in Short Rotation Poplar and Annual Field Crops. Forests, 9(4), 168. https://doi.org/10.3390/f9040168