Hydrology of Drained Peatland Forest: Numerical Experiment on the Role of Tree Stand Heterogeneity and Management

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
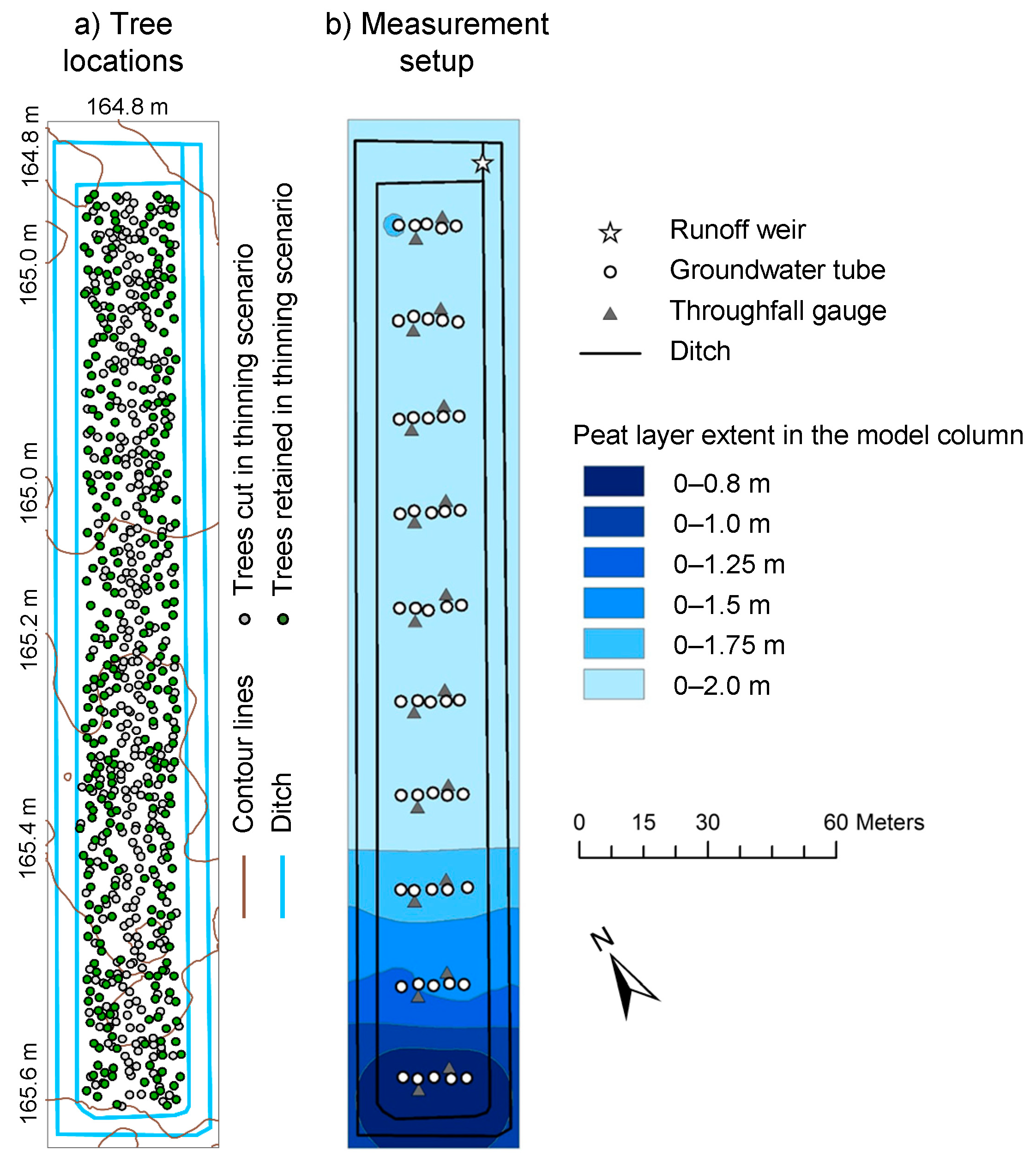
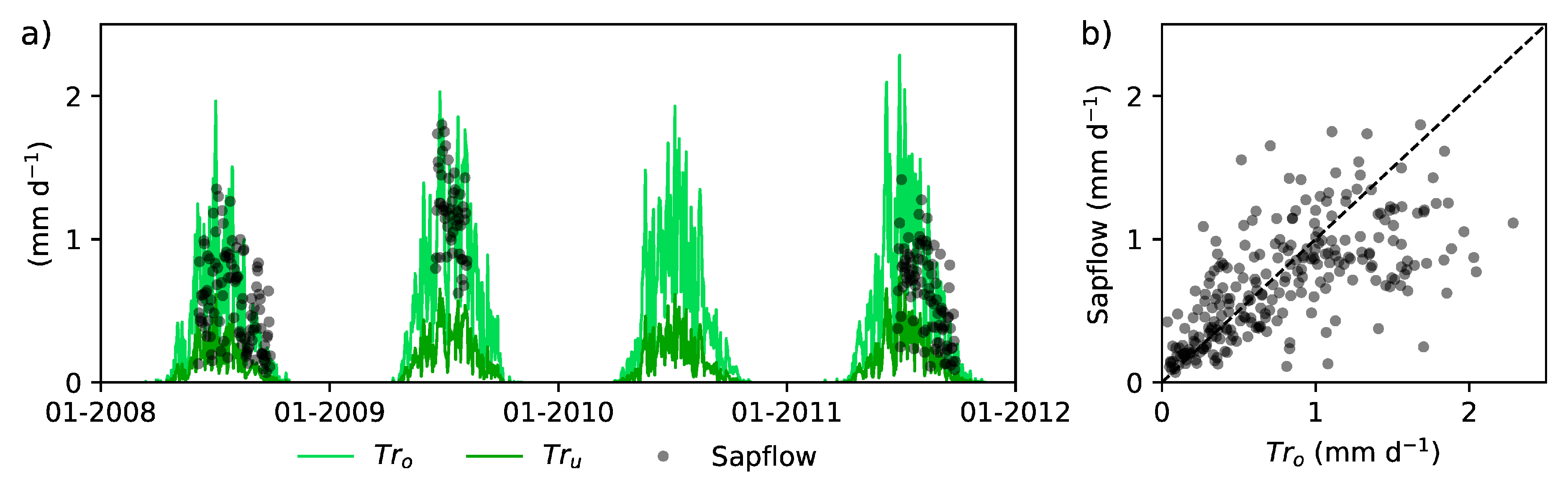
2.1. Sattasuo Experimental Area and Measurements
2.2. 3-D Distributed Hydrological Model FLUSH
2.3. Discretization and Parameterization
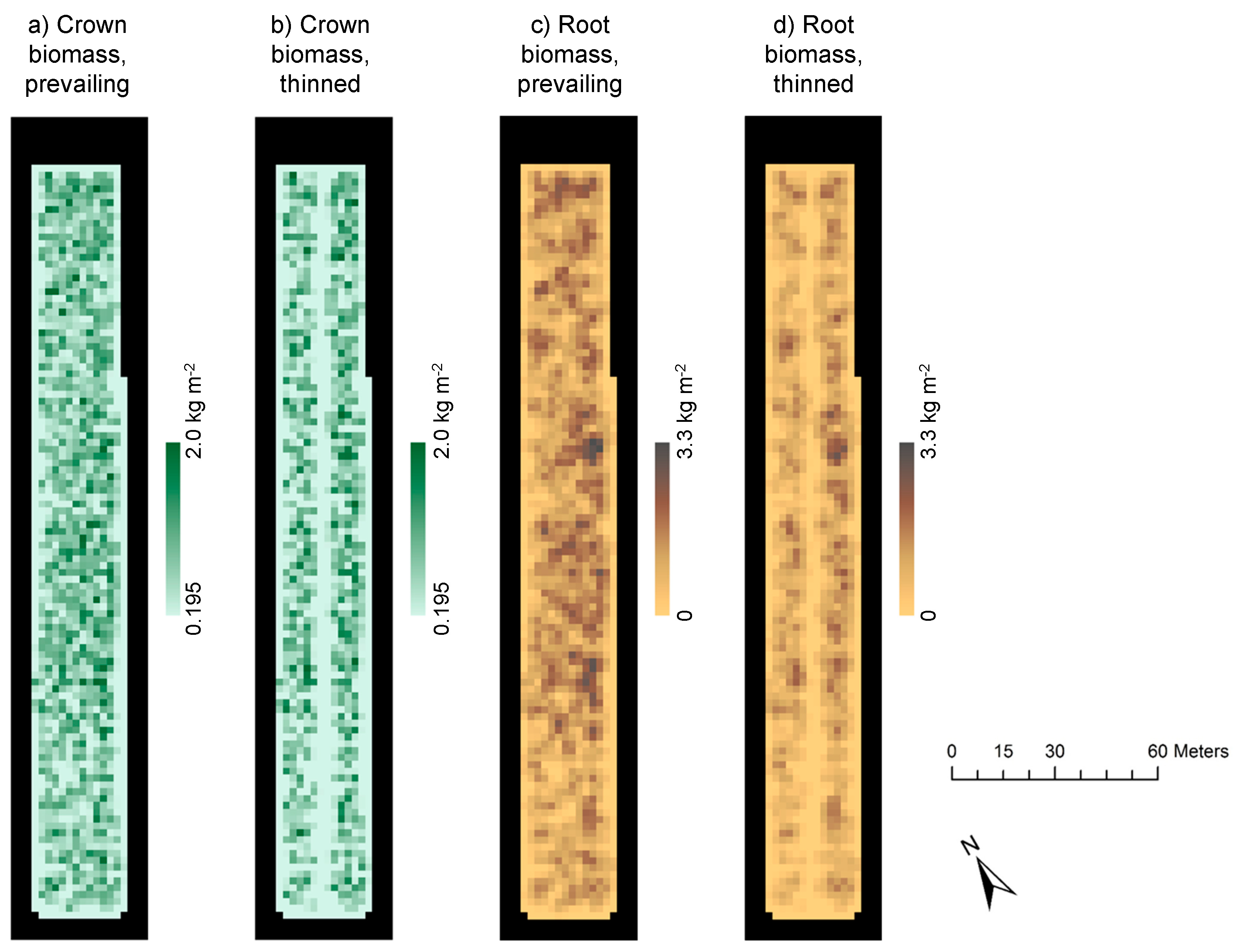
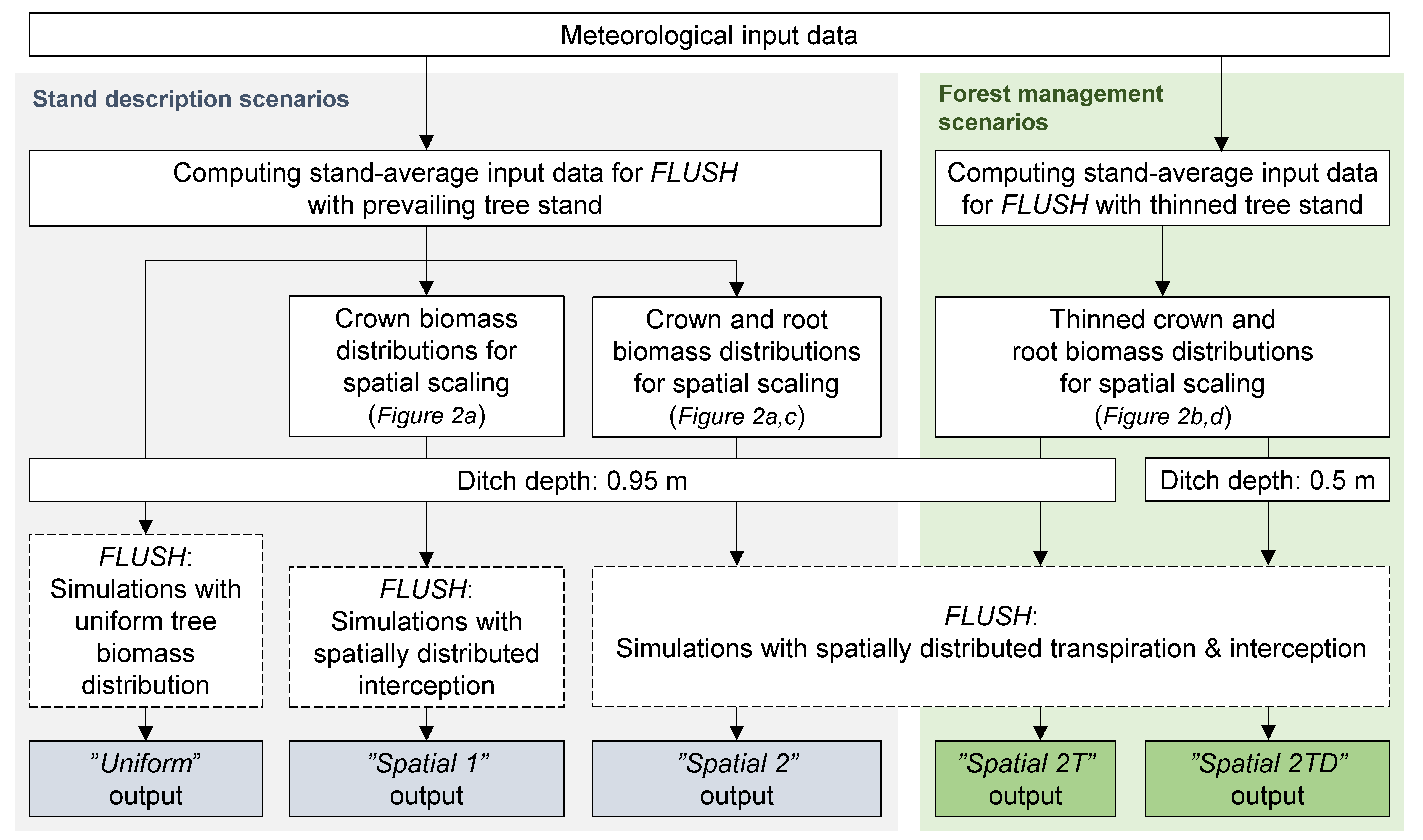
2.4. Computing and Spatial Scaling of Inputs for the FLUSH Model
2.5. Model Scenarios
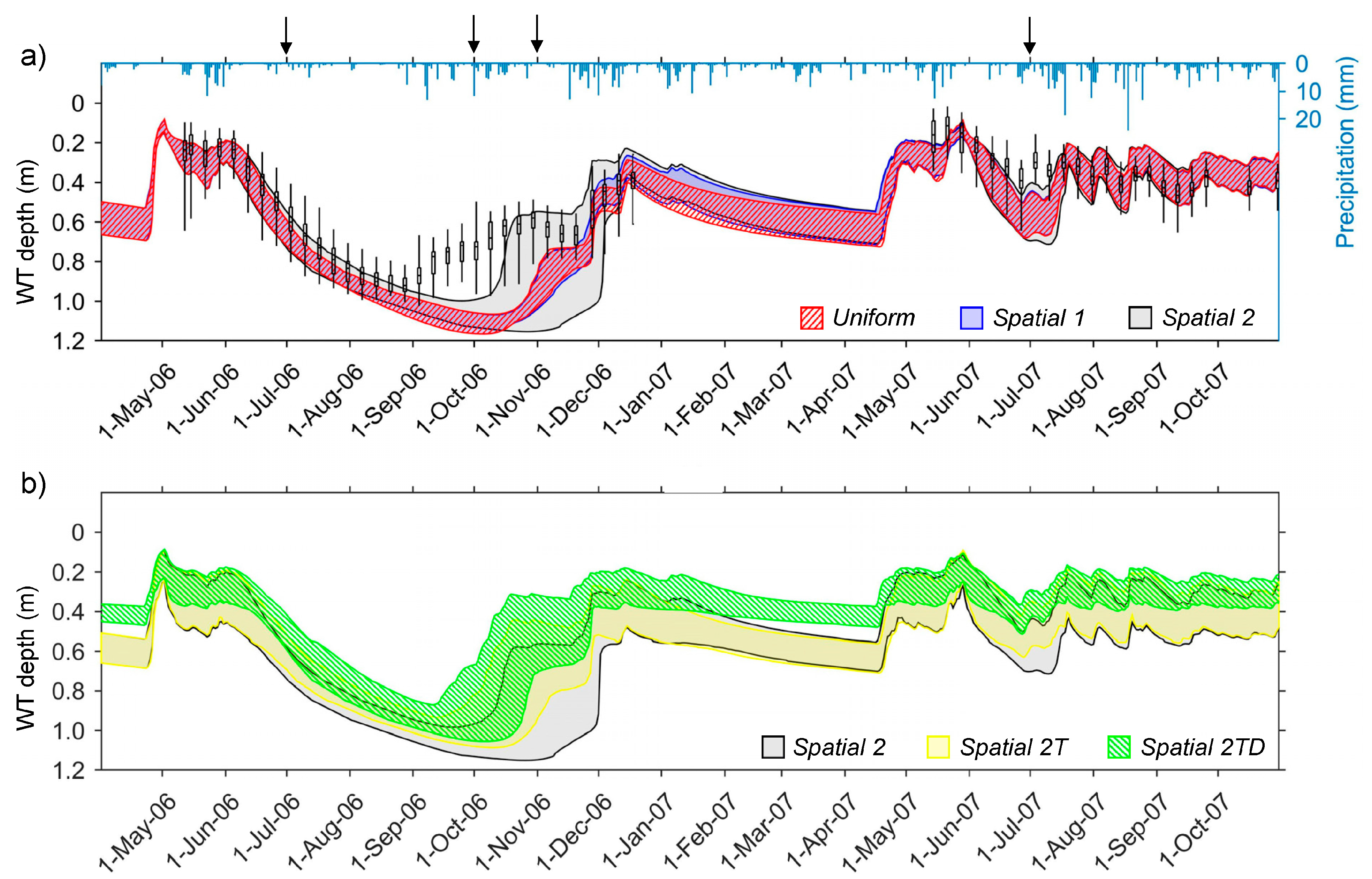
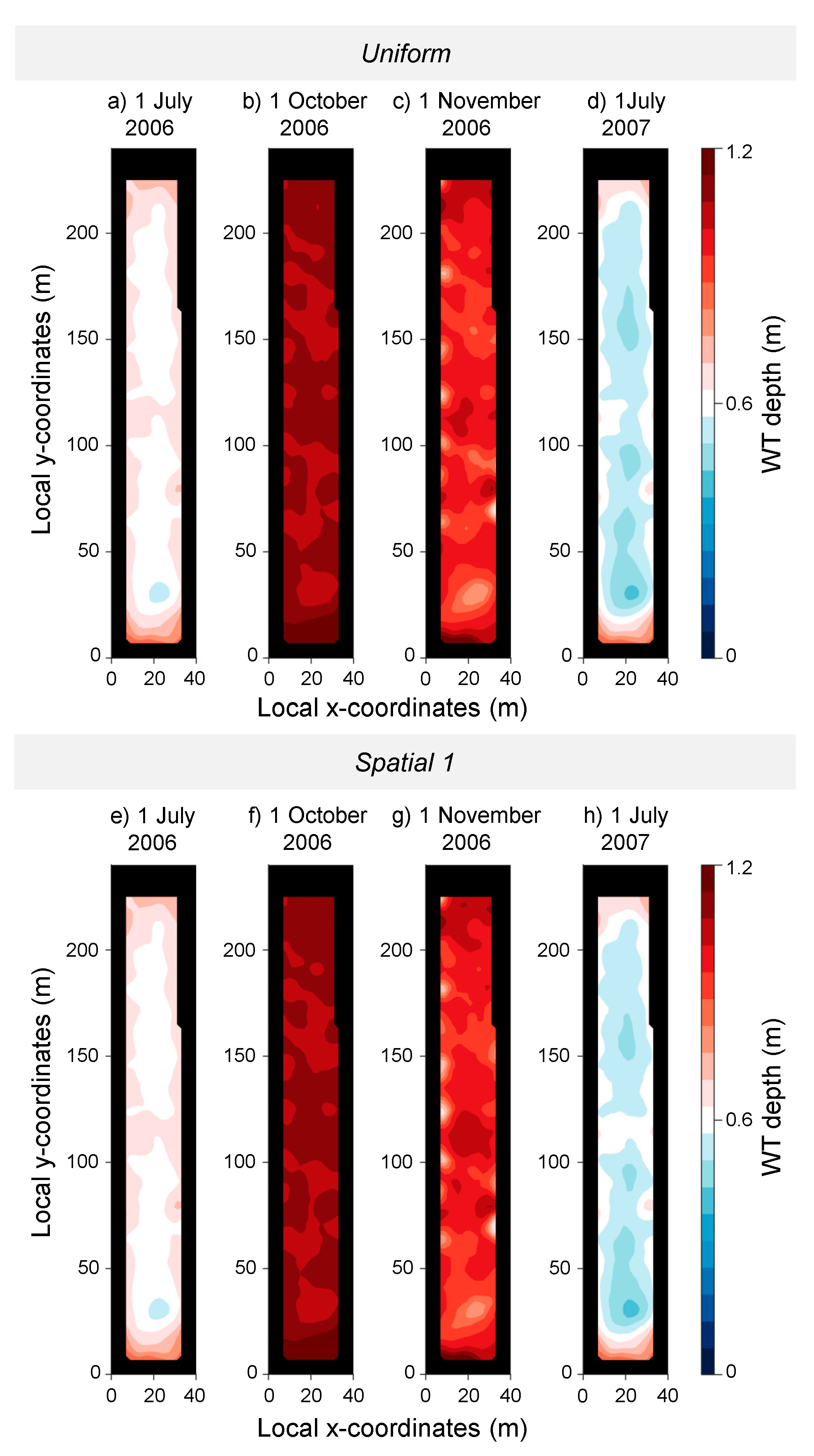
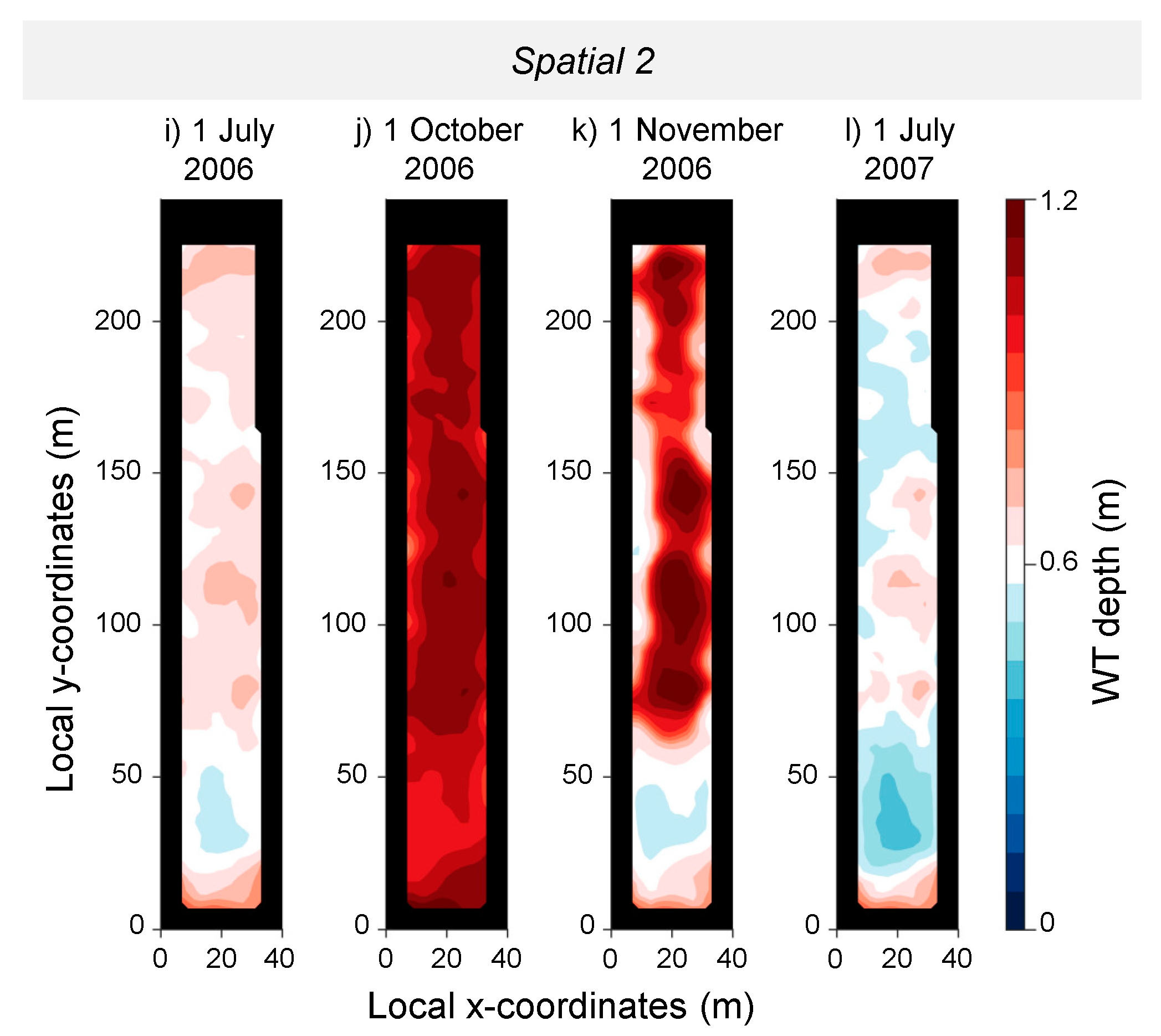
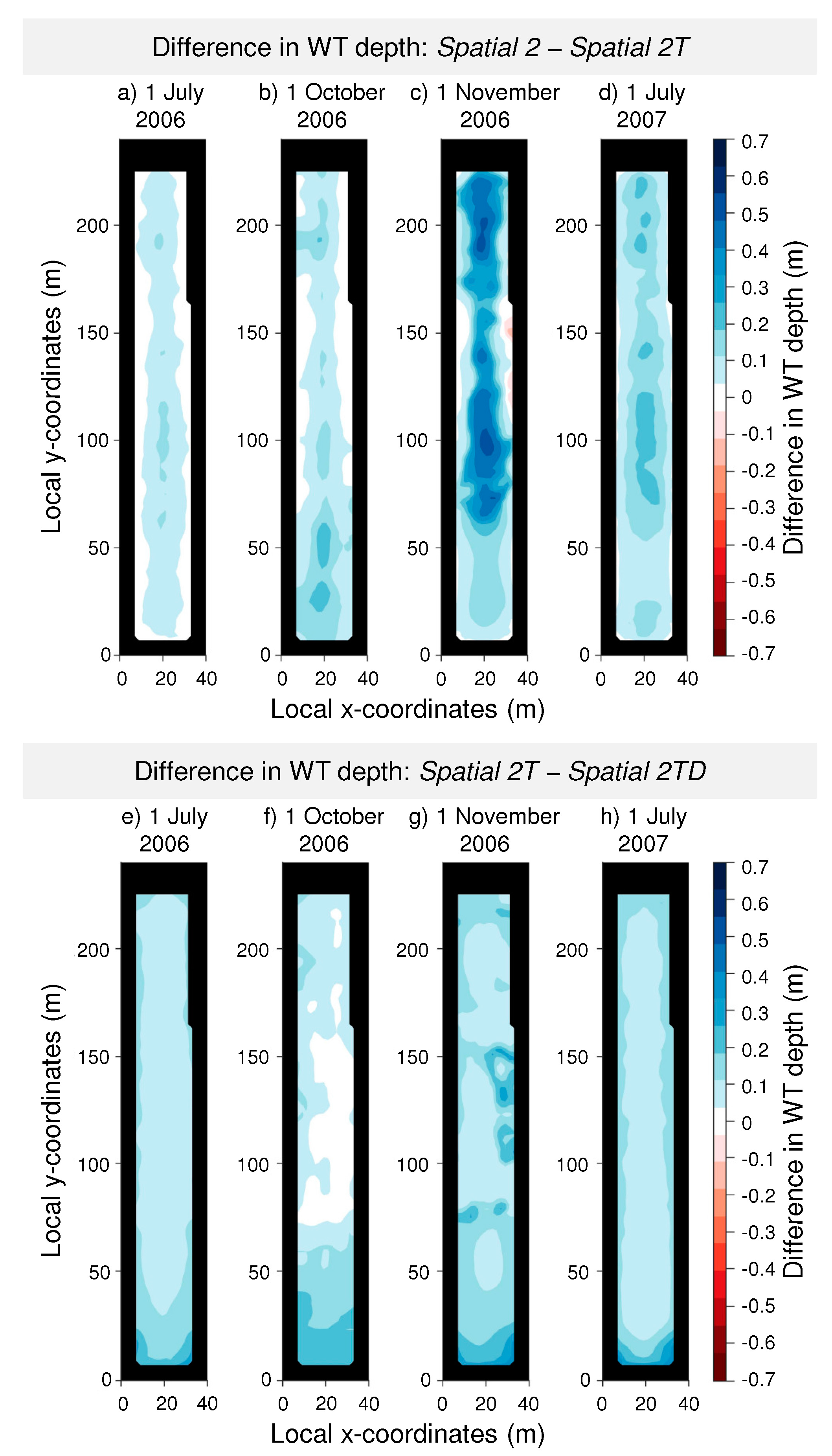
3. Results
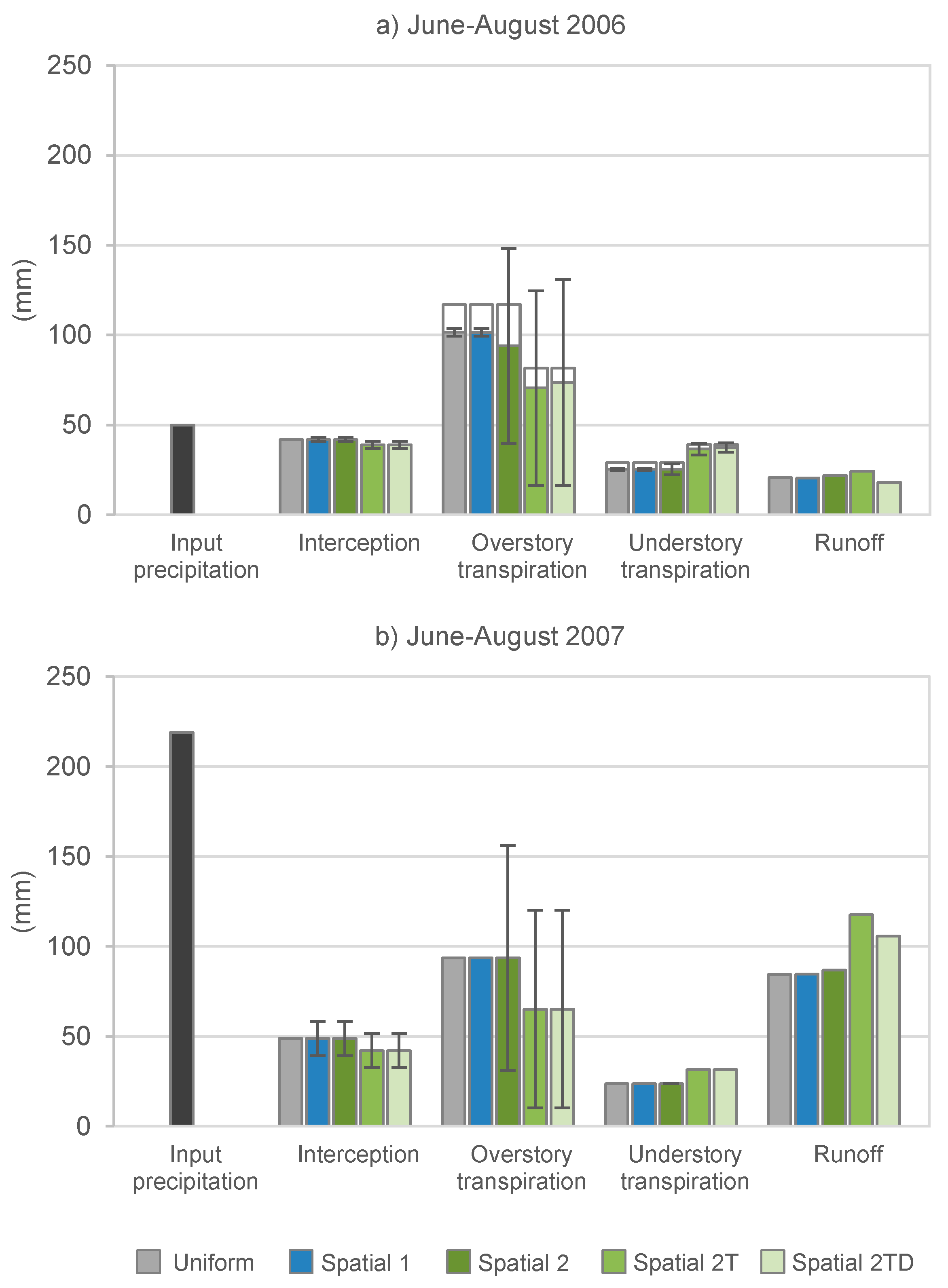
3.1. Stand Description Scenarios: The Role of Spatially Distributed Interception and Transpiration in Water Balance and WT Levels
3.2. Forest Management Scenarios: The Effects of Forest Management on WT Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Calculation of Interception Rate
Appendix A.2. Calculation of Spatial Distribution of Interception
Appendix A.3. Calculation of Over- and Understory Potential Transpiration Rates

References
- Hökkä, H.; Kaunisto, S.; Korhonen, K.T.; Päivänen, J.; Reinikainen, A.; Tomppo, E. Suomen suometsät 1951–1994. Metsät. Aikak. 2002, 2A, 201–357. [Google Scholar] [CrossRef]
- Paavilainen, E.; Päivänen, J. Peatland Forestry: Ecology and Principles; Springer: Berlin, Germany, 1995; ISBN 3-540-58252-5. [Google Scholar]
- Préfontaine, G.; Jutras, S. Variation in stand density, black spruce individual growth and plant community following 20 years of drainage in post-harvest boreal peatlands. For. Ecol. Manag. 2017, 400, 321–331. [Google Scholar] [CrossRef]
- Päivänen, J.; Hånell, B. Peatland Ecology and Forestry–A Sound Approach; University of Helsinki Department of Forest Sciences Publications: Helsinki, Finland, 2012; Volume 3. [Google Scholar]
- Sarkkola, S.; Hökkä, H.; Koivusalo, H.; Nieminen, M.; Ahti, E.; Päivänen, J.; Laine, J. Role of tree stand evapotranspiration in maintaining satisfactory drainage conditions in drained peatlands. Can. J. For. Res. 2010, 40, 1485–1496. [Google Scholar] [CrossRef]
- Sarkkola, S.; Nieminen, M.; Koivusalo, H.; Laurén, A.; Ahti, E.; Launiainen, S.; Nikinmaa, E.; Marttila, H.; Laine, J.; Hökkä, H. Domination of growing-season evapotranspiration over runoff makes ditch network maintenance in mature peatland forests questionable. Mires Peat 2013, 11, 1–11. [Google Scholar]
- Seuna, P. Long-Term Influence of Forestry Drainage on the Hydrology of an Open Bog in Finland; Publications of the Water Research Institute, National Board of Waters: Helsinki, Finland, 1981; Volume 43, pp. 3–14. [Google Scholar]
- Korkalainen, T.; Lauren, A.; Koivusalo, H.; Kokkonen, T. Impacts of peatland drainage on the properties of typical water flow paths determined from a digital elevation model. Hydrol. Res. 2008, 39, 359–368. [Google Scholar] [CrossRef]
- Hökkä, H.; Repola, J.; Laine, J. Quantifying the interrelationship between tree stand growth rate and water table level in drained peatland sites within Central Finland. Can. J. For. Res. 2008, 38, 1775–1783. [Google Scholar] [CrossRef]
- Hökkä, H.; Uusitalo, J.; Lindeman, H.; Ala-Ilomäki, J. Performance of weather parameters in predicting growing season water table depth variations on drained forested peatlands—A case study from southern Finland. Silva Fenn. 2016, 50, 1–12. [Google Scholar] [CrossRef]
- Haahti, K.; Koivusalo, H.; Hökkä, H.; Nieminen, M.; Sarkkola, S. Vedenpinnan syvyyden spatiaaliseen vaihteluun vaikuttavat tekijät ojitetussa suometsikössä. Suo 2012, 63, 107–121. [Google Scholar]
- Kaila, A.; Sarkkola, S.; Laurén, A.; Ukonmaanaho, L.; Koivusalo, H.; Xiao, L.; O’Driscoll, C.; Asam, Z.U.Z.; Tervahauta, A.; Nieminen, M. Phosphorus export from drained Scots pine mires after clear-felling and bioenergy harvesting. For. Ecol. Manag. 2014, 325, 99–107. [Google Scholar] [CrossRef]
- Ukonmaanaho, L.; Starr, M.; Kantola, M.; Laurén, A.; Piispanen, J.; Pietilä, H.; Perämäki, P.; Merilä, P.; Fritze, H.; Tuomivirta, T.; et al. Impacts of forest harvesting on mobilization of Hg and MeHg in drained peatland forests on black schist or felsic bedrock. Environ. Monit. Assess. 2016, 188, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ojanen, P.; Minkkinen, K.; Penttilä, T. The current greenhouse gas impact of forestry-drained boreal peatlands. For. Ecol. Manag. 2013, 289, 201–208. [Google Scholar] [CrossRef]
- Nieminen, M.; Hökkä, H.; Laiho, R.; Juutinen, A.; Ahtikoski, A.; Pearson, M.; Kojola, S.; Sarkkola, S.; Launiainen, S.; Valkonen, S.; et al. Could continuous cover forestry be an economically and environmentally feasible management option on drained boreal peatlands? For. Ecol. Manag. 2018, 424, 78–84. [Google Scholar] [CrossRef]
- Vítková, L.; Ní Dhubháin, Á. Transformation to continuous cover forestry: A review. Ir. For. 2013, 70, 119–140. [Google Scholar]
- Tian, S.; Youssef, M.A.; Skaggs, R.W.; Amatya, D.M.; Chescheir, G.M. DRAINMOD-FOREST: Integrated Modeling of Hydrology, Soil Carbon and Nitrogen Dynamics, and Plant Growth for Drained Forests. J. Environ. Qual. 2012, 41, 764. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, E.J.; Flewelling, J.W.; Skaggs, R.W. Hydrologic model for drained forest watershed. J. Irrig. Drain. Eng. 1992, 118, 242–255. [Google Scholar] [CrossRef]
- He, H.; Jansson, P.E.; Svensson, M.; Björklund, J.; Tarvainen, L.; Klemedtsson, L.; Kasimir, A. Forests on drained agricultural peatland are potentially large sources of greenhouse gases—Insights from a full rotation period simulation. Biogeosciences 2016, 13, 2305–2318. [Google Scholar] [CrossRef]
- Koivusalo, H.; Ahti, E.; Laurén, A.; Kokkonen, T.; Karvonen, T.; Nevalainen, R.; Finér, L. Impacts of ditch cleaning on hydrological processes in a drained peatland forest. Hydrol. Earth Syst. Sci. 2008, 12, 1211–1227. [Google Scholar] [CrossRef]
- Smolander, M. Vesitase Ojitetussa Suometsikössä. Master’s Thesis, Aalto University, Espoo, Finland, 2011. [Google Scholar]
- Haahti, K.; Younis, B.A.; Stenberg, L.; Koivusalo, H. Unsteady flow simulation and erosion assessment in a ditch network of a drained peatland forest catchment in eastern Finland. Water Resour. Manag. 2014, 28, 5175–5197. [Google Scholar] [CrossRef]
- Nieminen, M.; Palviainen, M.; Sarkkola, S.; Laurén, A.; Marttila, H.; Finér, L. A synthesis of the impacts of ditch network maintenance on the quantity and quality of runoff from drained boreal peatland forests. Ambio 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.; Albertson, J.; Zi, T.; Xu, X.; Kiely, G. How does afforestation affect the hydrology of a blanket peatland? A modelling study. Hydrol. Process. 2013, 27, 3577–3588. [Google Scholar] [CrossRef]
- Haahti, K.; Warsta, L.; Kokkonen, T.; Younis, B.A.; Koivusalo, H. Distributed hydrological modeling with channel network flow of a forestry drained peatland site. Water Resour. Res. 2016, 52. [Google Scholar] [CrossRef]
- Warsta, L.; Karvonen, T.; Koivusalo, H.; Paasonen-Kivekäs, M.; Taskinen, A. Simulation of water balance in a clayey, subsurface drained agricultural field with three-dimensional FLUSH model. J. Hydrol. 2013, 476, 395–409. [Google Scholar] [CrossRef]
- Hökkä, H.; Laine, J. Suopuustojen rakenteen kehitys ojituksen jälkeen. Silva Fenn. 1988, 22, 45–65. [Google Scholar] [CrossRef]
- Päivänen, J. Tree stand structure on pristine peatlands and its change after forest drainage. Int. Peat J. 1999, 9, 66–72. [Google Scholar]
- Sarkkola, S.; Hökkä, H.; Penttilä, T. Natural development of stand structure in peatland scots pine following drainage: Results based on long-term monitoring of permanent sample plots. Silva Fenn. 2004, 38, 405–412. [Google Scholar] [CrossRef]
- Calders, K.; Newnham, G.; Burt, A.; Murphy, S.; Raumonen, P.; Herold, M.; Culvenor, D.; Avitabile, V.; Disney, M.; Armston, J.; et al. Nondestructive estimates of above-ground biomass using terrestrial laser scanning. Methods Ecol. Evol. 2015, 6, 198–208. [Google Scholar] [CrossRef]
- Kotivuori, E.; Korhonen, L.; Packalen, P. Nationwide airborne laser scanning based models for volume, biomass and dominant height in Finland. Silva Fenn. 2016, 50, 1–28. [Google Scholar] [CrossRef]
- The Finnish Meteorological Institute’s Open Data. Available online: https://en.ilmatieteenlaitos.fi/open-data (accessed on 20 November 2017).
- Vasander, H.; Laine, J. Site type classification on drained peatlands. In Finland-Fenland-Research and Sustainable Utilisation of Mires and Peat; Korhonen, R., Korpela, L., Sarkkola, S., Eds.; Finnish Peatland Society and Maahenki Ltd.: Helsinki, Finland, 2008; pp. 146–151. [Google Scholar]
- National Land Survey of Finland. File Service of Open Data. Available online: https://tiedostopalvelu.maanmittauslaito.fi/tp/kartta?lang=en (accessed on 12 December 2017).
- Granier, A. Une nouvelle methode pour la measure du flux de seve brute dans le tronc des arbres. Ann. des Sci. For. 1985, 42, 193–200. [Google Scholar] [CrossRef]
- Warsta, L. Modelling Water Flow and Soil Erosion in Clayey, Subsurface Drained Agricultural Fields. Ph.D. Thesis, Aalto University, Espoo, Finland, 2011. [Google Scholar]
- Launiainen, S.; Salmivaara, A.; Kieloaho, A.; Peltoniemi, M.; Guan, M. Spatiotemporal variability of evapotranspiration in boreal forest catchments: A novel upscaling by process models and open GIS data. In Proceedings of the Geophysical Research Abstracts, EGU General Assembly, Vienna, Austria, 8–13 April 2018; Volume 20, p. 13286. [Google Scholar]
- Van Genuchten, M.T. A Closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- Lagergren, F.; Lindroth, A. Transpiration response to soil moisture in pine and spruce trees in Sweden. Agric. For. Meteorol. 2002, 112, 67–85. [Google Scholar] [CrossRef]
- Päivänen, J. Hydraulic conductivity and water retention in peat soils. Acta For. Fenn. 1973, 129, 1–70. [Google Scholar] [CrossRef]
- O’Kelly, B.C.; Pichan, S.P. Effects of decomposition on the compressibility of fibrous peat—A review. Geomech. Geoeng. 2013, 8, 286–296. [Google Scholar] [CrossRef]
- Lewis, C.; Albertson, J.; Xu, X.; Kiely, G. Spatial variability of hydraulic conductivity and bulk density along a blanket peatland hillslope. Hydrol. Process. 2012, 26, 1527–1537. [Google Scholar] [CrossRef]
- Vakkilainen, P. Hydrologian perusteita. In Maan vesi-ja Ravinnetalous; Ojitus, Kastelu ja Ympäristö. 2. Täydennetty Painos; Paasonen-Kivekäs, M., Peltomaa, R., Vakkilainen, P., Äijö, H., Eds.; Salaojayhdistys ry: Helsinki, Finland, 2016; pp. 73–128. [Google Scholar]
- Marklund, L.G. Biomassafunktioner för tall, gran och björk i Sverige. SLU Rapp. 1988, 45, 1–73. [Google Scholar]
- Čermák, J.; Riguzzi, F.; Ceulemans, R. Scaling up from the individual tree to the stand level in Scots pine. I. Needle distribution, overall crown and root geometry. Ann. Sci. For. 1998, 55, 63–88. [Google Scholar] [CrossRef]
- Nagel, J.; Albert, M.; Schmidt, M. Das waldbauliche Prognose- und Entscheidungsmodell BWINPro 6.1. Forst und Holz 2002, 57, 486–493. [Google Scholar]
- Sarkkola, S.; Hökkä, H.; Ahti, E.; Koivusalo, H.; Nieminen, M. Depth of water table prior to ditch network maintenance is a key factor for tree growth response. Scand. J. For. Res. 2012, 27, 649–658. [Google Scholar] [CrossRef]
- Lyr, H.; Erdmann, A.; Hoffmann, G.; Köhler, S. Uber den diurnalen Wachtumsrhytmus von Gehölzen. Flora 1968, 157, 615–624. [Google Scholar]
- Chertov, O.G.; Komarov, A.S.; Nadporozhskaya, M.; Bykhovets, S.S.; Zudin, S.L. ROMUL—A model of forest soil organic matter dynamics as a substantial tool for forest ecosystem modeling. Ecol. Model. 2001, 138, 289–308. [Google Scholar] [CrossRef]
- Päivänen, J. Sateen jakaantuminen erilaisissa metsiköissä. Silva Fenn. 1966, 119, 1–37. [Google Scholar] [CrossRef]
- Koivusalo, H.; Kokkonen, T. Snow processes in a forest clearing and in a coniferous forest. J. Hydrol. 2002, 262, 145–164. [Google Scholar] [CrossRef]
- Ilvesniemi, H.; Pumpanen, J.; Duursma, R.; Hari, P.; Keronen, P.; Kolari, P.; Kulmala, M.; Mammarella, I.; Nikinmaa, E.; Rannik, Ü.; et al. Water balance of a boreal Scots pine forest. Boreal Environ. Res. 2010, 15, 375–396. [Google Scholar]
- Pypker, T.G.; Levia, D.F.; Staelens, J.; Van Stan, J.T., II. Canopy Structure in Relation to Hydrological and Biogeochemical Fluxes. In Forest Hydrology and Biogeochemistry; Levia, D., Carlyle-Moses, D., Tanaka, T., Eds.; Springer: Dordrech, The Netherlands, 2011; ISBN 978-94-007-1363-5. [Google Scholar]
- Granier, A.; Loustau, D.; Bréda, N. A generic model of forest canopy conductance dependent on climate, soil water availability and leaf area index. Ann. For. Sci. 2000, 57, 755–765. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Harley, P.C. Scaling carbon dioxide and water vapour exchange from leaf to canopy in a deciduous forest. II. Model testing and application. Plant. Cell Environ. 1995, 18, 1157–1173. [Google Scholar] [CrossRef]
- Launiainen, S.; Katul, G.G.; Kolari, P.; Lindroth, A.; Lohila, A.; Aurela, M.; Varlagin, A.; Grelle, A.; Vesala, T. Do the energy fluxes and surface conductance of boreal coniferous forests in Europe scale with leaf area? Glob. Chang. Biol. 2016, 22, 4096–4113. [Google Scholar] [CrossRef] [PubMed]
- Laine-Kaulio, H.; Backnäs, S.; Karvonen, T.; Koivusalo, H.; McDonnell, J.J. Lateral subsurface stormflow and solute transport in a forested hillslope: A combined measurement and modeling approach. Water Resour. Res. 2014, 50, 8159–8178. [Google Scholar] [CrossRef]
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. For. Ecol. Manag. 2015, 338, 124–138. [Google Scholar] [CrossRef]
- O’Driscoll, C.; O’Connor, M.; de Eyto, E.; Poole, R.; Rodgers, M.; Zhan, X.; Nieminen, M.; Xiao, L. Whole-tree harvesting and grass seeding as potential mitigation methods for phosphorus export in peatland catchments. For. Ecol. Manag. 2014, 319, 176–185. [Google Scholar] [CrossRef]
- Launiainen, S.; Katul, G.G.; Lauren, A.; Kolari, P. Coupling boreal forest CO2, H2O and energy flows by a vertically structured forest canopy—Soil model with separate bryophyte layer. Ecol. Modell. 2015, 312, 385–405. [Google Scholar] [CrossRef]
- Muukkonen, P.; Mäkipää, R. Empirical biomass models of understorey vegetation in boreal forests according to stand and site attributes. Boreal Environ. Res. 2006, 11, 355–369. [Google Scholar]
- Williams, T.G.; Flanagan, L.B. Effect of changes in water content on photosynthesis, transpiration and discrimination against 13CO2 and C18O16 in Pleurozium and Sphagnum. Pleur. Sphagnum 1996, 108, 38–46. [Google Scholar]
- Hock, R. Temperature index melt modelling in mountain areas. J. Hydrol. 2003, 282, 104–115. [Google Scholar] [CrossRef]
- Peltoniemi, M.; Pulkkinen, M.; Aurela, M.; Pumpanen, J.; Kolari, P.; Mäkelä, A. A semi-empirical model of boreal forest gross primary production, evapotranspiration, and soil water–calibration and sensitivity analysis. Boreal Environ. Res. 2015, 20, 151–171. [Google Scholar]
- Härkönen, S.; Lehtonen, A.; Manninen, T.; Tuominen, S.; Peltoniemi, M. Estimating forest leaf area index using satellite images: Comparison of k-NN based Landsat-NFI LAI with MODIS-RSR based LAI product for Finland. Boreal Environ. Res. 2015, 20, 181–195. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth (m) | θs (m3 m−3) | θr (m3 m−3) | α (m−1) | β (−) | KVsat (m h−1) | KHsat (m h−1) |
|---|---|---|---|---|---|---|
| 0–0.1 | 0.95 | 0.098 | 33.85 | 1.4 | 3 * | 10 × KVsat |
| 0.1–0.2 | 0.95 | 0.098 | 33.85 | 1.4 | 1 * | 10 × KVsat |
| 0.2–0.3 | 0.9141 | 0 | 1.0469 | 1.3115 | 0.03 | 10 × KVsat |
| 0.3–0.4 | 0.801 | 6.74 × 10−8 | 1.383 | 1.3047 | 0.01 | 10 × KVsat |
| 0.4–0.6 | 0.801 | 6.74 × 10−8 | 1.383 | 1.3047 | 0.002 | 10 × KVsat |
| 0.6–2.0 (peat) | 0.801 | 6.74 × 10−8 | 1.383 | 1.3047 | 0.0001 | KVsat |
| 0.6–2.0 (mineral) | 0.55 | 0 | 1.1 | 1.8 | 0.0036 | KHsat |
| Amount of Time Suffering from Wetness (%) | ||
|---|---|---|
| Scenario | July–August 2006 | July–August 2007 |
| Spatial 2 | 0 | 5 |
| Spatial 2T | 0 | 42 |
| Spatial 2TD | 0 | 61 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stenberg, L.; Haahti, K.; Hökkä, H.; Launiainen, S.; Nieminen, M.; Laurén, A.; Koivusalo, H. Hydrology of Drained Peatland Forest: Numerical Experiment on the Role of Tree Stand Heterogeneity and Management. Forests 2018, 9, 645. https://doi.org/10.3390/f9100645
Stenberg L, Haahti K, Hökkä H, Launiainen S, Nieminen M, Laurén A, Koivusalo H. Hydrology of Drained Peatland Forest: Numerical Experiment on the Role of Tree Stand Heterogeneity and Management. Forests. 2018; 9(10):645. https://doi.org/10.3390/f9100645
Chicago/Turabian StyleStenberg, Leena, Kersti Haahti, Hannu Hökkä, Samuli Launiainen, Mika Nieminen, Ari Laurén, and Harri Koivusalo. 2018. "Hydrology of Drained Peatland Forest: Numerical Experiment on the Role of Tree Stand Heterogeneity and Management" Forests 9, no. 10: 645. https://doi.org/10.3390/f9100645
APA StyleStenberg, L., Haahti, K., Hökkä, H., Launiainen, S., Nieminen, M., Laurén, A., & Koivusalo, H. (2018). Hydrology of Drained Peatland Forest: Numerical Experiment on the Role of Tree Stand Heterogeneity and Management. Forests, 9(10), 645. https://doi.org/10.3390/f9100645