Allometric Equations for Estimating Biomass and Carbon Stocks in the Temperate Forests of North-Western Mexico

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
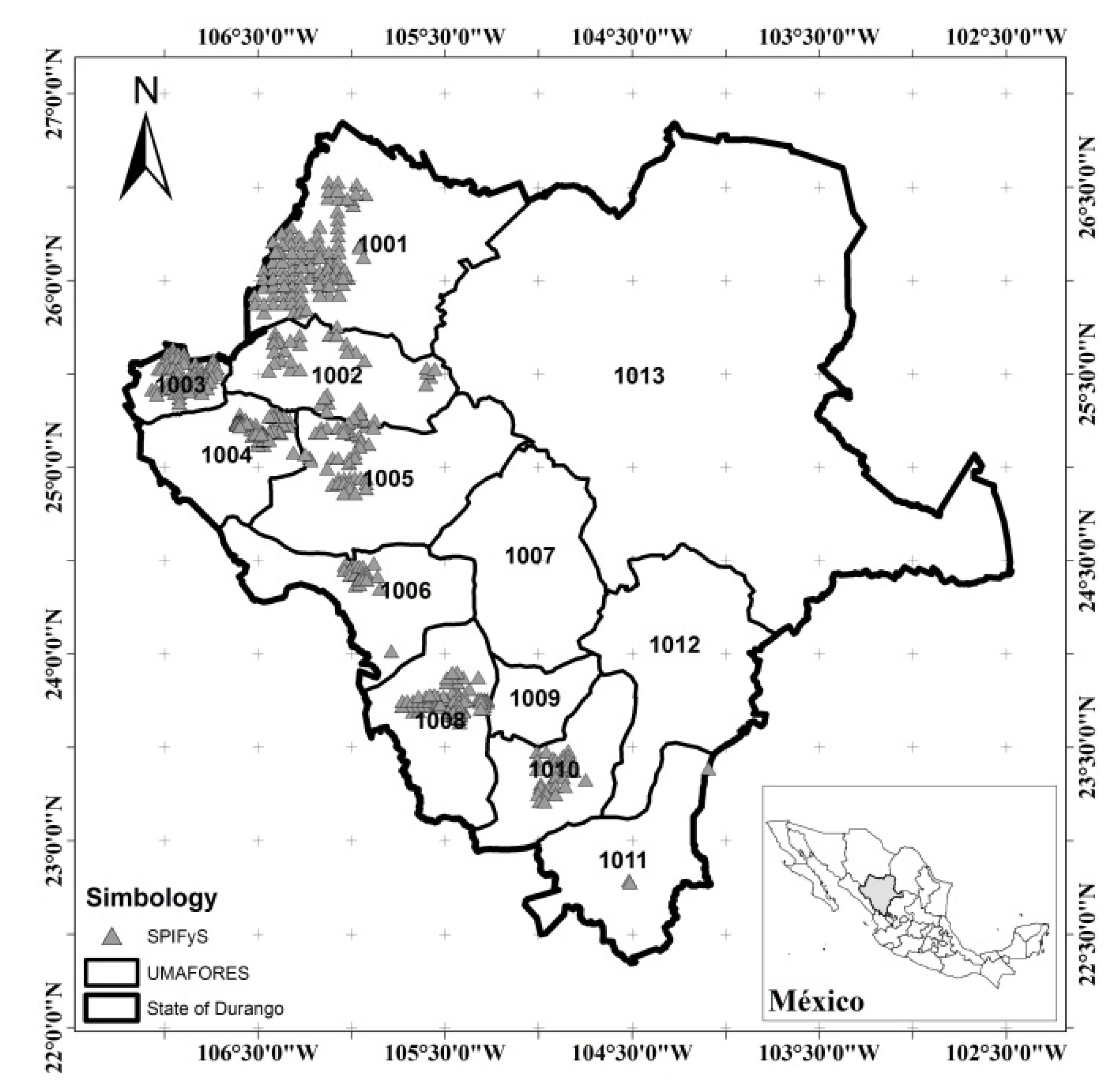
2.1. Study Area
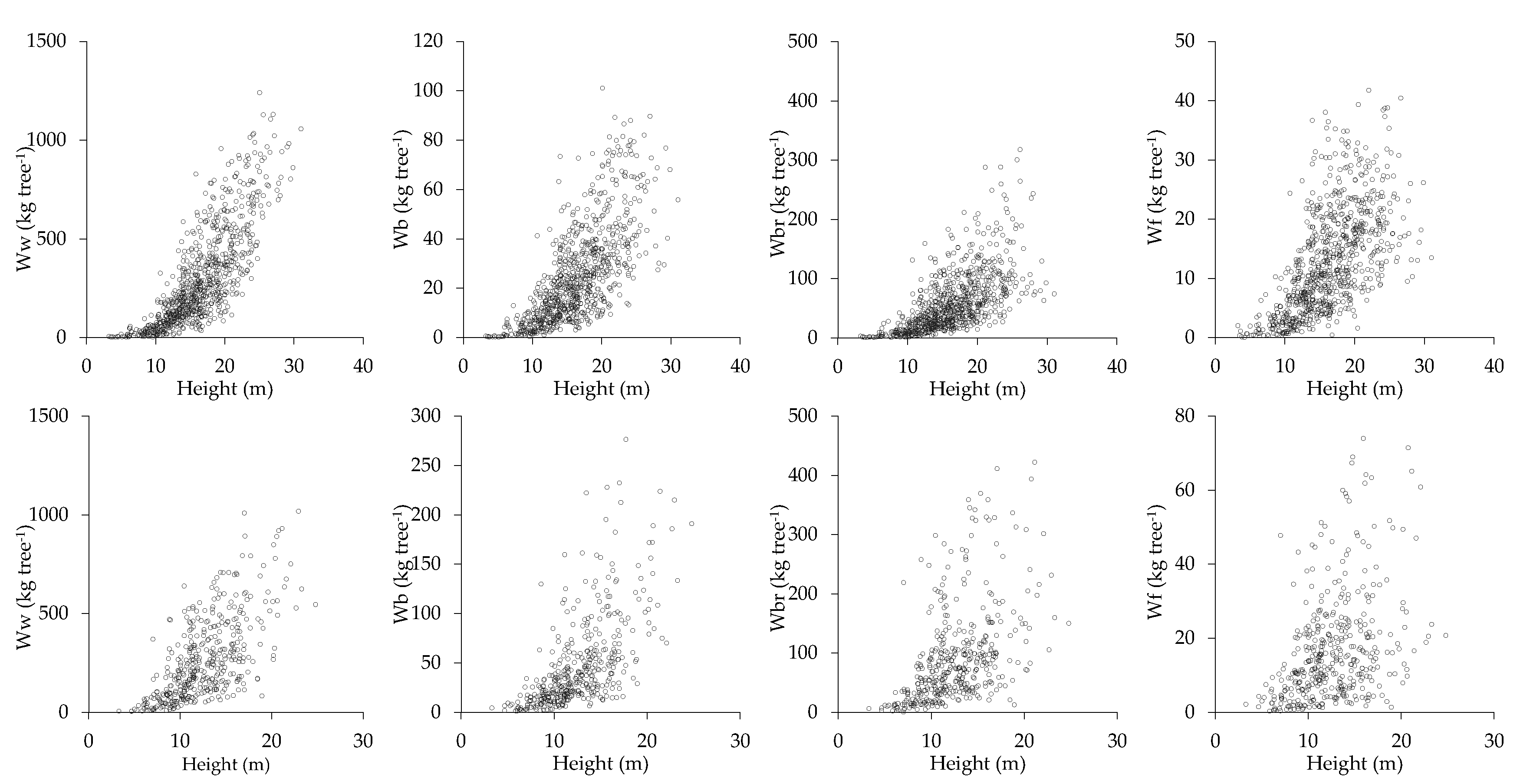
2.2. Biomass Data
2.3. Procedures for Developing the Species-Specific Biomass Equations
2.3.1. Basic Models
2.3.2. Simultaneous Fitting of Tree Biomass Components and Total AGB
2.3.3. Comparison of Equations
2.4. Applying the Proposed Above-Ground Biomass Equations
2.5. Machine Learning Techniques (MLTs)
3. Results
3.1. Tree-Level Biomass Equations
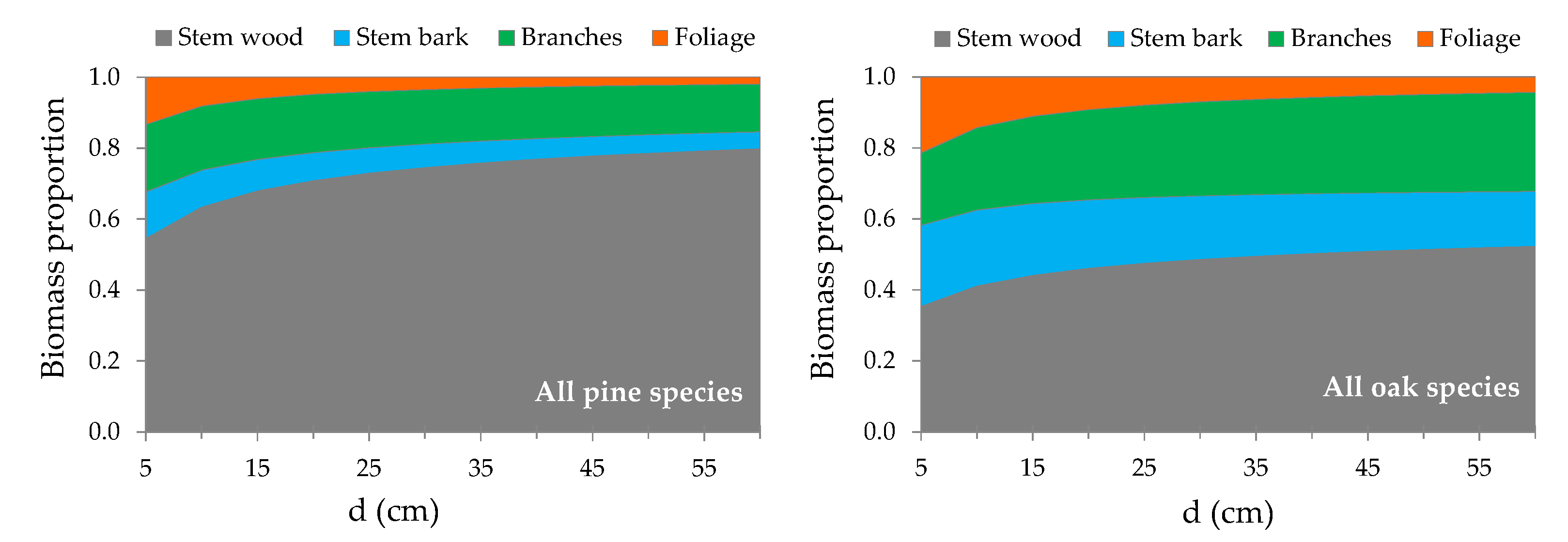
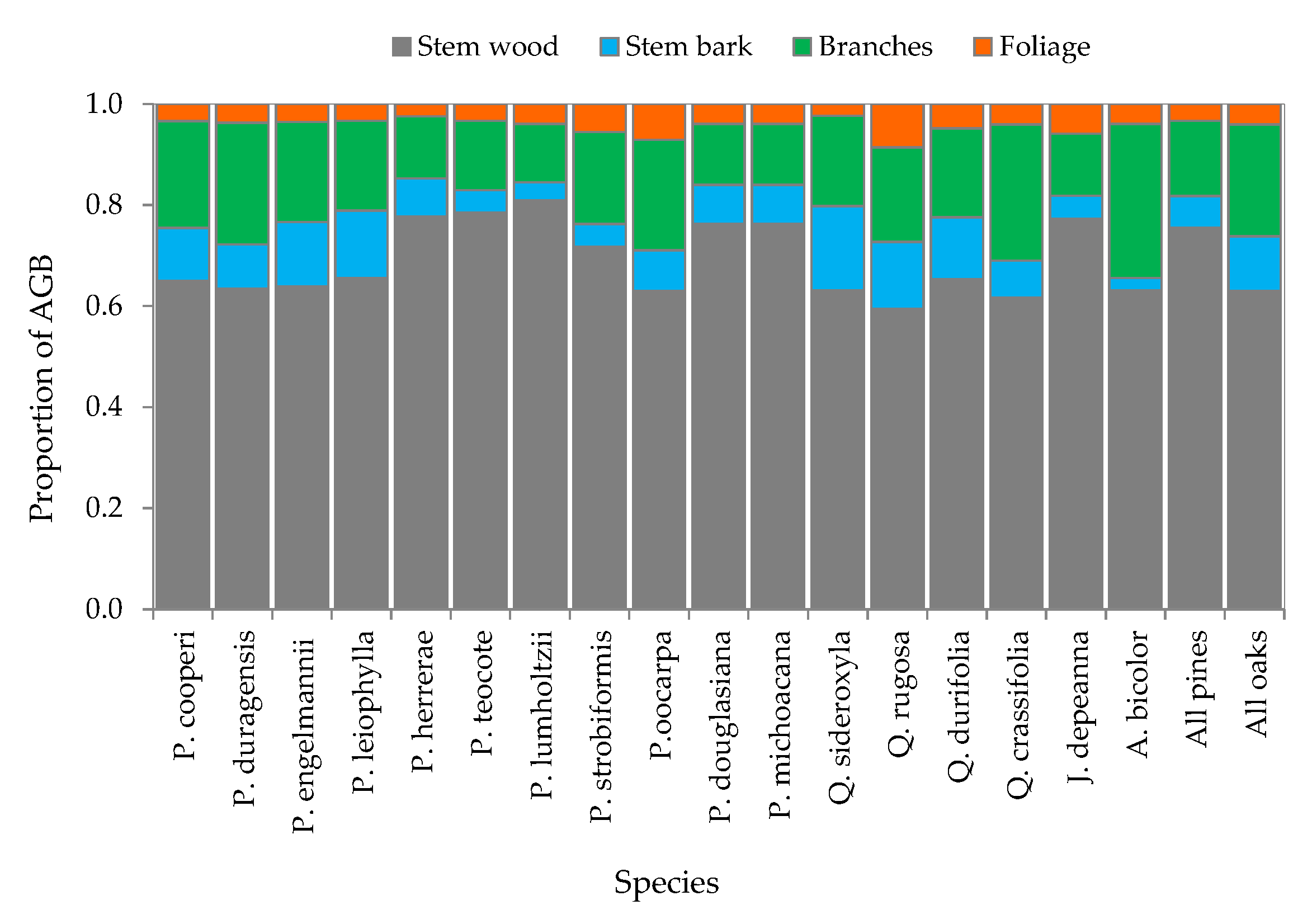
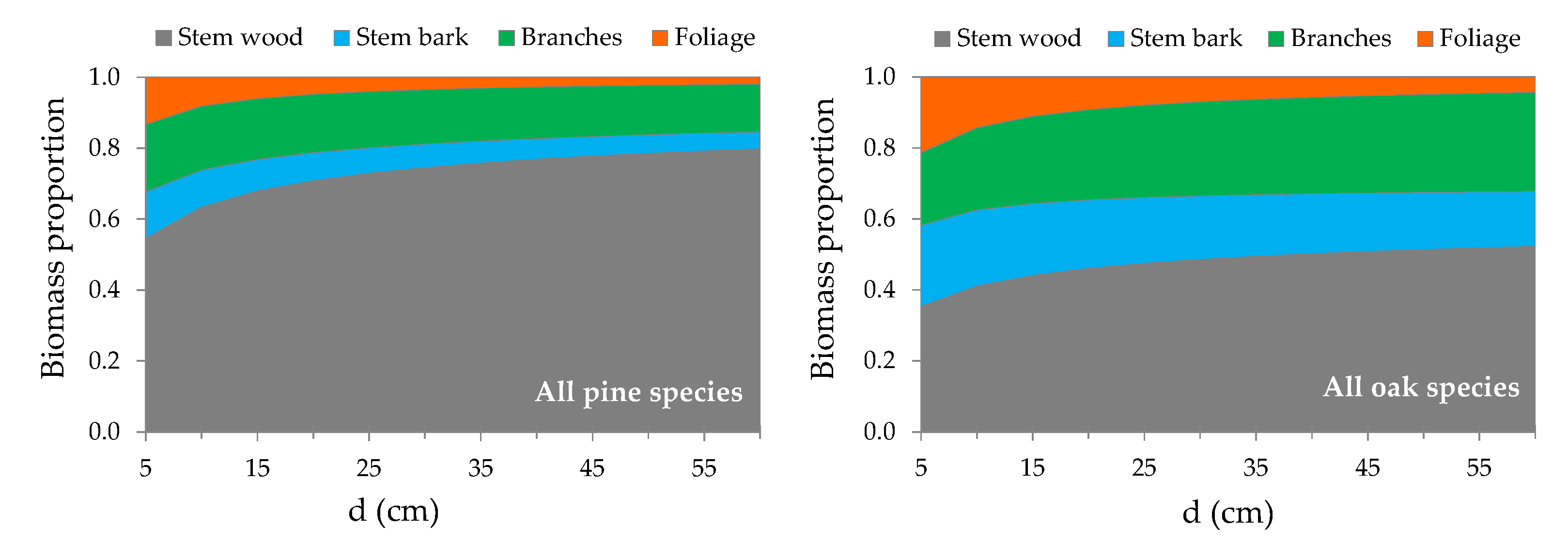
3.2. Above-Ground Biomass Allocation
3.3. Carbon Fractions in Different Tree Components
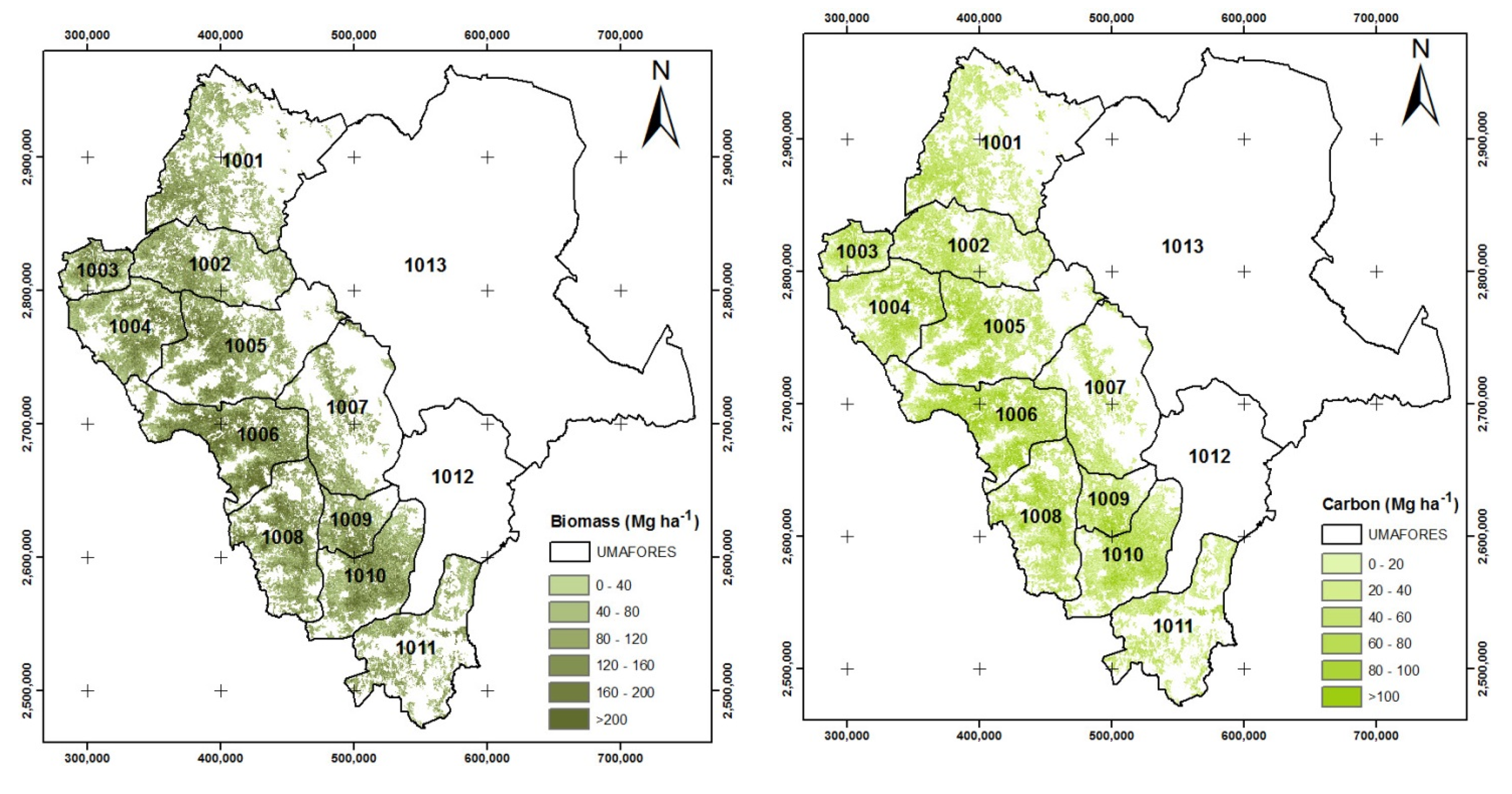
3.4. Biomass and Carbon Estimates in the Permanent Research Plots
4. Discussion
4.1. The Allometric Equations
4.2. Contribution of Components to Total AGB
4.3. Machine Learning Techniques (MLT)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Data Availability
References
- Hollinger, D. Defining a Landscape-Scale Monitoring Tier for the North American Carbon Program. In Field Measurements for Forest Carbon Monitoring; Hoover, C.M., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 3–17. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Gómez-García, E.; Diéguez-Aranda, U.; Barrio-Anta, M.; Crecente-Campo, F. Aboveground stand-level biomass estimation: A comparison of two methods for major forest species in northwest Spain. Ann. Forest Sci. 2012, 69, 735–746. [Google Scholar] [CrossRef]
- Hall, G.M.J.; Wiser, S.K.; Allen, R.B.; Beets, P.N.; Goulding, C.J. Strategies to estimate national forest carbon stocks from inventory data: The 1990 New Zealand baseline. Glob. Chang. Biol. 2001, 7, 389–403. [Google Scholar] [CrossRef]
- Wirth, C.; Schumacher, J.; Schulze, E.D. Generic biomass functions for Norway spruce in Central Europe—A meta-analysis approach toward prediction and uncertainty estimation. Tree Physiol. 2004, 24, 121–139. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Ecología y Cambio Climático (INECC) y Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). Primer Informe Bienal de Actualización ante la Convención Marco de las Naciones Unidas sobre el Cambio Climático; INECC/SEMARNAT: Mexico City, México, 2015. [Google Scholar]
- Salimon, C.I.; Putz, F.E.; Menezes-Filho, L.; Anderson, A.; Silveira, M.; Brown, I.F.; Oliveira, L.C. Estimating state-wide biomass carbon stocks for a REDD plan in Acre, Brazil. Forest Ecol. Manag. 2011, 262, 555–560. [Google Scholar] [CrossRef]
- Acosta-Mireles, M.; Vargas-Hernández, J.; Velázquez-Martínez, A.; Etchevers-Barra, J.D. Estimación de la biomasa aérea mediante el uso de relaciones alométricas en seis especies arbóreas en Oaxaca, México. Agrociencia 2002, 36, 725–736. [Google Scholar]
- Návar, J.J. Allometric equations for tree species and carbon stocks for forests of northwestern Mexico. Forest Ecol. Manag. 2009, 257, 427–434. [Google Scholar] [CrossRef]
- Návar-Cháidez, J.J. Biomass allometry for tree species of northwestern Mexico. Trop. Subtrop. Agroecosyst. 2010, 12, 507–519. Available online: https://www.cabdirect.org/cabdirect/abstract/20103282813 (accessed on 13 September 2015).
- Rodríguez-Ortíz, G.; Aldrete, A.; González-Hernández, V.A.; De los Santos-Posadas, H.M.; Gómez-Guerrero, A.; Fierros-González, A.M. ¿Afectan los aclareos la acumulación de biomasa aérea en una plantación de Pinus patula? Agrociencia 2011, 45, 719–732. [Google Scholar]
- Silva-Arredondo, F.M.; Návar-Cháidez, J.J. Estimación de factores de expansión de carbono en comunidades forestales templadas del norte de Durango, México. Rev. Chapingo Ser. Cienc. 2009, 15, 155–163. [Google Scholar]
- Villegas-Jiménez, D.E.; Rodríguez-Ortiz, G.; Velasco-Velasco, V.A.; Ruiz-Luna, J.; Carrillo-Rodríguez, J.C.; Ramírez-Sánchez, S.E. Partición de biomasa aérea en procedencias de Pinus greggii plantadas en el sur de México. Rev. Fitotec. Mex. 2013, 36, 421–427. [Google Scholar]
- Díaz-Franco, R.; Acosta-Mireles, M.; Carrillo, F.A.; Rodríguez, E.B.; Ayala, E.F.; Barra, J.D. Determinación de ecuaciones alométricas para estimar biomasa y carbono en Pinus patula Schl. et Cham. Madera Bosques 2007, 13, 25–34. [Google Scholar] [CrossRef]
- Mendoza-Ponce, A.; Galicia, L. Aboveground and belowground biomass and carbon pools in highland temperate forest landscape in Central Mexico. Forestry 2010, 83, 497–506. [Google Scholar] [CrossRef]
- Búrquez, A.; Martínez-Yrízar, A.; Núñez, S.; Quintero, T.; Aparicio, A. Aboveground biomass in three Sonoran Desert communities: Variability within and among sites using replicated plot harvesting. J. Arid. Environ. 2010, 74, 1240–1247. [Google Scholar] [CrossRef]
- Aguirre-Salado, C.A.; Treviño-Garza, E.J.; Aguirre-Calderón, O.A.; Jiménez-Pérez, J.; González-Tagle, M.A.; Miranda-Aragón, L.; Valdez-Lazalde, J.; Aguirre-Salado, A.; Sánchez-Díaz, G. Forest Cover Mapping in North-Central Mexico: A Comparison of Digital Image Processing Methods. GIScience Remote Sens. 2012, 49, 895–914. [Google Scholar] [CrossRef]
- Méndez-González, J.; Luckie-Navarrete, S.L.; Capo-Arteaga, M.A.; Nájera-Luna, J.A. Ecuaciones alométricas y estimación de incrementos en biomasa aérea y carbono en una plantación mixta de Pinus devoniana Lindl. y P. pseudostrobus Lindl., en Guanajuato, México. Agrociencia 2011, 45, 479–491. [Google Scholar]
- Rojas-García, F.; De Jong, B.J.; Martínez-Zurimendí, P.; Paz-Pellat, F. Database of 478 allometric equations to estimate biomass for Mexican trees and forests. Ann. Forest Sci. 2015, 72, 835–864. [Google Scholar] [CrossRef]
- Challenger, A. Utilización de los Ecosistemas Terrestres de México. Pasado, Presente y Future; Comisión Nacional para el Conocimiento y uso de la Biodiversidad (CONABIO): Mexico City, México, 1998. [Google Scholar]
- CONABIO. Estrategia Nacional Sobre Biodiversidad de México; Comisión Nacional para el Conocimiento y uso de la Biodiversidad: Mexico City, México, 2000. [Google Scholar]
- SEMARNAT. Anuario Estadístico de la Producción Forestal 2013; In Secretaría de Medio Ambiente y Recursos Naturales: Mexico City, México, 2014. [Google Scholar]
- Du, L.; Zhou, T.; Zou, Z.; Zhao, X.; Huang, K.; Wu, H. Mapping Forest Biomass Using Remote Sensing and National Forest Inventory in China. Forests 2014, 5, 1267–1283. [Google Scholar] [CrossRef]
- Zianis, D.; Mencuccini, M. On simplifying allometric analyses of forest biomass. Forest Ecol. Manag. 2004, 187, 311–332. [Google Scholar] [CrossRef]
- Walker, W.; Baccini, A.; Nepstad, M.; Horning, N.; Knight, D.; Braun, E.; Bausch, A. Guía de Campo para la Estimación de Biomasa y Carbono Forestal; Versión 1.0; Woods Hole Research Center: Falmouth, MA, USA, 2011. [Google Scholar]
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef]
- López-Serrano, P.; López-Sánchez, C.; Díaz-Varela, R.; Corral-Rivas, J.; Solís-Moreno, R.; Vargas-Larreta, B.; Alvarez-Gonzalez, J.G. Estimating biomass of mixed and uneven-aged forests using spectral data and a hybrid model combining regression trees and linear models. iForest 2015. [Google Scholar] [CrossRef]
- Hernández-Stefanoni, J.L.; Gallardo-Cruz, J.A.; Meave, J.A.; Dupuy, J.M. Combining geostatistical models and remotely sensed data to improve tropical plant richness mapping. Ecol. Indic. 2011, 11, 1046–1056. [Google Scholar] [CrossRef]
- Aguirre-Salado, C.A.; Treviño-Garza, E.J.; Aguirre-Calderón, O.A.; Jiménez-Pérez, J.; González-Taglel, M.A.; Valdéz-Lazalde, J.R.; Sánchez-Díaz, G.; Haapanen, R.; Aguirre-Salado, A.I.; Miranda-Aragón, L. Mapping aboveground biomass by integrating geospatial and forest inventory data through a k-nearest neighbour strategy in North Central Mexico. J. Arid Land 2014, 1, 80–96. [Google Scholar] [CrossRef]
- Hutchinson, M.E. Continent-wide data assimilation using thin plate smoothing splines. In Data assimilation systems; BMRC Research Report No.27; Jasper, J.D., Ed.; Bureau of Meteorology Research Centre: Melbourne, Australia, 1991; pp. 104–113. [Google Scholar]
- García-Arévalo, A.; González-Elizondo, S. Pináceas de Durango, 2nd ed.; Instituto de Ecología-Comisión Nacional Forestal: Durango, México, 2003. [Google Scholar]
- Parresol, B.R. Additivity of nonlinear biomass equations. Can. J. Forest Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Bi, H.; Long, Y.; Turner, J.; Lei, Y.; Snowdon, P.; Li, Y.; Harper, R.; Zerihun, A.; Ximenes, F. Additive prediction of aboveground biomass for Pinus radiata (D. Don) plantations. Forest Ecol. Manag. 2010, 259, 2301–2314. [Google Scholar] [CrossRef]
- Institute S. SAS/STAT 9.2. User´s Guide Release; SAS Institute Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Greene, W.H. Econometric Analysis, 4th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 1999. [Google Scholar]
- Corral-Rivas, J.J.; Vargas, B.; Wehenkel, C.; Aguirre, O.; Álvarez, J.; Rojo, A. Guía para el Establecimiento de Sitios de Inventario Periódico Forestal y de Suelos del Estado de Durango; Facultad de Ciencias Forestales, Universidad Juárez del Estado de Durango: Durango, México, 2009. [Google Scholar]
- ESPA Ordering Interface. Available online: https://espa.cr.usgs.gov (accessed on 8 May 2017).
- Masek, J.G.; Vermote, E.F.; Saleous, N.E.; Wolfe, R.; Hall, F.G.; Huemmrich, K.F.; Gao, F.; Kutler, J.; Lim, T.-K. A Landsat Surface Reflectance Dataset for North America, 1990–2000. IEEE Geosci. Remote Sens. Lett. 2006, 3, 68–72. [Google Scholar] [CrossRef]
- NASA. Landsat 7 Science Data Users Handbook; NASA: Washington, DC, USA, 2011.
- Rouse, J.W.; Haas, R.H.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. Remote Sens. 1974, 1, 309–313. [Google Scholar]
- Huete, A.R. A soil-adjusted vegetation index (SAVI). Remote Sens. Environ. 1988, 25, 295–309. [Google Scholar] [CrossRef]
- Glenn, E.; Huete, A.; Nagler, P.; Nelson, S. Relationship between remotely-sensed Vegetation Indices, Canopy Attributes and Plant Physiological Processes: What Vegetation Indices Can and Cannot Tell Us About the Landscape. Sensors 2008, 8, 2136–2160. [Google Scholar] [CrossRef] [PubMed]
- McNab, W.H. Terrain Shape Index: Quantifying effect of minor landforms on tree height. Forest Sci. 1989, 35, 91–104. [Google Scholar]
- Moore, I.D.; Norton, T.W.; Williams, J.E. Modelling environmental heterogeneity in forested landscapes. J. Hydrol. 1993, 150, 717–747. [Google Scholar] [CrossRef]
- INEGI. Mexican Continuous Elevation 3.0 (CEM3.0). Available online: http://www.inegi.org.mx/geo/contenidos/datosrelieve/continental/descarga.aspx (accessed on 8 May 2015).
- Wilson, J.P.; Gallant, J.C. Digital terrain analysis. In Terrain Analysis: Principles and Applications; Wilson, J.P., Gallant, J.C., Eds.; John Wiley and Sons, Inc.: New York, NY, USA, 2000; pp. 1–26. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Huang, D.; Knyazikhin, Y.; Wang, W.; Deering, D.W.; Stenberg, P.; Shabanov, N.; Tan, B.; Myneni, R.B. Stochastic transport theory for investigating the three-dimensional canopy structure from space measurements. Remote Sens. Environ. 2008, 11, 35–50. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Hostert, P.; St-Louis, V.; Radeloff, V.C. Using image texture to map farmland field size: A case study in Eastern Europe. J. Land Use Sci. 2009, 4, 85–107. [Google Scholar] [CrossRef]
- Shevade, S.K.; Keerthi, S.S.; Bhattacharyya, C.; Murthy, K.R.K. Improvements to the SMO algorithm for SVM regression. Neural Networks. IEEE Trans. Neural Netw. 2000, 11, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Fassnacht, F.E.; Hartig, F.; Latifi, H.; Berger, C.; Hernández, J.; Corvalán, P.; Kochf, B. Importance of sample size, data type and prediction method for remote sensing-based estimations of aboveground forest biomass. Remote Sens. Environ. 2014, 154, 102–114. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Hall, M.; Frank, E.; Holmes, G.; Pfahringer, B.; Reutemann, P.; Witten, I.H. The WEKA data mining software: An update. SIGKDD Explor. 2009, 11, 10–18. [Google Scholar] [CrossRef]
- IPCC National Greenhouse Gas Inventories Programme. 2006 Guidelines for National Greenhouse Gas Inventories; Institute for Global Environmental Strategies (IGES): Hayama, Kanagawa, Japan, 2006. [Google Scholar]
- António, N.; Tomé, M.; Tomé, J.; Soares, P.; Fontes, L. Effect of tree, stand, and site variables on the allometry of Eucalyptus globulus tree biomass. Can. J. Forest Res. 2007, 37, 895–906. [Google Scholar] [CrossRef]
- Blujdea, V.N.B.; Pilli, R.; Dutca, I.; Ciuvat, L.; Abrudan, I.V. Allometric biomass equations for young broadleaved trees in plantations in Romania. Forest Ecol. Manag. 2012, 264, 172–184. [Google Scholar] [CrossRef]
- González-García, M.; Hevia, A.; Majada, J.; Barrio-Anta, M. Above-ground biomass estimation at tree and stand level for short rotation plantations of Eucalyptus nitens (Deane & Maiden) Maiden in Northwest Spain. Biomass Bioenergy 2013, 54, 147–157. [Google Scholar] [CrossRef]
- Johansson, T. Biomass equations for determining fractions of pendula and pubescent birches growing on abandoned farmland and some practical implications. Biomass Bioenergy 1999, 16, 223–238. [Google Scholar] [CrossRef]
- Porté, A.; Trichet, P.; Bert, D.; Loustau, D. Allometric relationships for branch and tree woody biomass of Maritime pine (Pinus pinaster Ait.). Forest Ecol. Manag. 2002, 158, 71–83. [Google Scholar] [CrossRef]
- Jenkins, J.; Chojnacky, D.S.; Birdsey, R. National scale biomass estimators for United States tree species. Forest Sci. 2003, 49, 12–35. [Google Scholar]
- Reed, D.; Tomé, M. Total aboveground biomass and net dry matter accumulation by plant component in young Eucalyptus globulus in response to irrigation. Forest Ecol. Manag. 1998, 103, 21–32. [Google Scholar] [CrossRef]
- Bartelink, H.H. Allometric relationships on biomass and needle area of Douglas-fir. Forest Ecol. Manag. 1996, 86, 193–203. [Google Scholar] [CrossRef]
- Menéndez-Miguélez, M.; Canga, E.; Barrio-Anta, M.; Majada, J.; Álvarez-Álvarez, P. A three level system for estimating the biomass of Castanea sativa Mill. coppice stands in north-west Spain. Forest Ecol. Manag. 2013, 291, 417–426. [Google Scholar] [CrossRef]
- Lambert, M.C.; Ung, C.H.; Raulier, F. Canadian national tree aboveground biomass equations. Can. J. Forest Res. 2005, 35, 1996–2018. [Google Scholar] [CrossRef]
- Bi, H.; Turner, J.; Lambert, M. Additive biomass equations for native eucalypt forest trees of temperate Australia. Trees 2004, 18, 467–479. [Google Scholar] [CrossRef]
- Zhao, D.; Kane, M.; Markewitz, D.; Teskey, R.; Clutter, M. Additive tree biomass equations for midrotation loblolly pine plantations. Forest Sci. 2015, 61, 613–623. [Google Scholar] [CrossRef]
- Feldpausch, T.R.; Lloyd, J.; Lewis, S.L.; Brienen, R.J.; Gloor, M.; Monteagudo Mendoza, A.; Lopez-Gonzalez, G.; Banin, L.; Abu Salim, K.; Affum-Baffoe, K.; et al. Tree height integrated into pantropical forest biomass estimates. Biogeosciences 2012, 9, 3381–3403. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhao, P. Improving the accuracy of tree-level aboveground biomass equations with height classification at a large regional scale. Forest Ecol. Manag. 2013, 289, 153–163. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.; Parker, W. Estimating biomass of white spruce seedlings with vertical photo imagery. New Forests 2000, 20, 145–162. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Poulain, M.; Klein, D.; Krause, K.; Peña-Rojas, K.; Schmidt, H.; Schulte, A. Allometric above-belowground biomass equations for Nothofagus pumilio (Poepp. & Endl.) natural regeneration in the Chilean Patagonia. Ann. Forest Sci. 2009, 66, 513. [Google Scholar] [CrossRef]
- Zeng, W.; Tang, S.; Huag, G.; Zhang, M. Population classification and sample structure on modeling of single tree biomass equations for national biomass estimation in China. Forest Resour. Manag. 2010, 3, 16–23. [Google Scholar]
- Parresol, B.R. Assessing tree and stand biomass: A Review with examples and critical comparisons. Forest Sci. 1999, 45, 573–593. [Google Scholar]
- Carvalho, J.P.; Parresol, B.R. Additivity in tree biomass components of Pyrenean oak (Quercus pyrenaica Willd.). Forest Ecol. Manag. 2003, 179, 269–276. [Google Scholar] [CrossRef]
- Lima, A.; Suwa, R.; de Mello Ribeiro, G.; Kajimoto, T.; dos Santos, J.; da Silva, R.P.; de Souza, C.A.S.; de Barros, P.C.; Noguchi, H.; Ishizuka, M.; et al. Allometric models for estimating above- and below-ground biomass in Amazonian forests at São Gabriel da Cachoeira in the upper Rio Negro, Brazil. Forest Ecol. Manag. 2012, 277, 163–172. [Google Scholar] [CrossRef]
- Hunter, M.O.; Keller, M.; Victoria, D.; Morton, D.C. Tree height and tropical forest biomass estimation. Biogeosciences 2013, 10, 8385–8399. [Google Scholar] [CrossRef]
- Fonseca, W.; Rey, B.J.M.; Alice, F.E. Carbon accumulation in the biomass and soil of different aged secondary forests in the humid tropics of Costa Rica. Forest Ecol. Manag. 2011, 262, 1400–1408. [Google Scholar] [CrossRef]
- Redondo-Brenes, A.; Montagnini, F. Growth, productivity, aboveground biomass, and carbon sequestration of pure and mixed native tree plantations in the Caribbean lowlands of Costa Rica. Forest Ecol. Manag. 2006, 232, 168–178. [Google Scholar] [CrossRef]
- Correia, A.C.; Tomé, M.; Pacheco, C.A.; Faias, S.; Días, A.C.; Freire, J.; Carvalho, P.O.; Pereira, J.S. Biomass allometry and carbon factors for a Mediterranean pine (Pinus pinea L.) in Portugal. Forest Syst. 2010, 19, 418–433. [Google Scholar] [CrossRef]
- Pajtík, J.; Konôpka, B.; Lukac, M. Biomass functions and expansion factors in young Norway spruce (Picea abies [L.] Karst) trees. Forest Ecol. Manag. 2008, 256, 1096–1103. [Google Scholar] [CrossRef]
- Aquino-Ramírez, M.; Velázquez-Martínez, A.; Castellanos-Bolaños, J.F.; De los Santos-Posadas, H.M.; Etchevers-Barra, J.D. Partición de la biomasa aérea en tres especies arbóreas tropicales. Agrociencia 2015, 49, 299–314. [Google Scholar]
- Lu, D.; Chen, Q.; Wang, G.; Moran, E.; Batistella, M.; Zhang, M.; Vaglio Laurin, G.; Saah, D. Aboveground forest biomass estimation with Landsat and LiDAR data and uncertainty analysis of the estimates. Int. J. Forest Res. 2012, 2012, 1–16. [Google Scholar] [CrossRef]
- Lu, D.; Chen, Q.; Wang, G.; Liu, L.; Li, G.; Moran, E. A survey of remote sensing-based aboveground biomass estimation methods in forest ecosystems. Int. J. Digit. Earth 2014, 1–43. [Google Scholar] [CrossRef]
- López-Serrano, P.M.; López-Sánchez, C.A.; Álvarez-González, J.G.; García-Gutiérrez, J. A Comparison of Machine Learning Techniques Applied to Landsat-5 TM Spectral Data for Biomass Estimation. Can. J. Remote Sens. 2016, 42, 690–705. [Google Scholar] [CrossRef]
- López-Serrano, P.M.; Corral-Rivas, J.J.; Díaz-Varela, R.A.; Álvarez-González, J.G.; López-Sánchez, C.A. Evaluation of radiometric and atmospheric correction algorithms for aboveground forest biomass estimation using Landsat 5 TM data. Remote Sens. 2016, 8, 369. [Google Scholar] [CrossRef]
- López-Serrano, P.M.; López Sánchez, C.A.; Solís-Moreno, R.; Corral-Rivas, J.J. Geospatial Estimation of above Ground Forest Biomass in the Sierra Madre Occidental in the State of Durango, Mexico. Forests 2016, 7, 70. [Google Scholar] [CrossRef]
- López-Sánchez, C.A.; García-Ramírez, P.; Resl, R.; Hernández-Díaz, J.C.; López-Serrano, P.M.; Wehenkel, C. Modelling dasometric attributes of mixed and uneven-aged forests using Landsat-8 OLI spectral data in the Sierra Madre Occidental, Mexico. iForest 2017, 10, 288–295. [Google Scholar] [CrossRef]
- García-Gutiérrez, J.; Martínez-Álvarez, F.; Troncoso, A.; Riquelme, J.C. A comparison of machine learning regression techniques for LiDAR-derived estimation of forest variables. Neurocomputing 2015, 167, 24–31. [Google Scholar] [CrossRef]
- Guo, Z.D.; Hu, H.F.; Pan, Y.D.; Birdsey, R.A.; Fang, J.Y. Increasing biomass carbon stocks in trees outside forests in China over the last three decades. Biogeosciences 2014, 11, 4115–4122. [Google Scholar] [CrossRef]
- Tian, X.; Li, Z.; Su, Z.; Chen, E.; van der Tol, C.; Li, X.; Guo, Y.; Li, L.; Ling, F. Estimating montane forest above-ground biomass in the upper reaches of the Heihe River Basin using Landsat-TM data. Int. J. Remote Sens. 2014, 35, 7339–7362. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Species (n) | Value | Variable | Biomass Component | |||||
|---|---|---|---|---|---|---|---|---|
| d | h | Ww | Wb | Wbr | Wf | Wt | ||
| Pinus cooperi (103) | Max | 52.3 | 28.0 | 1586.5 | 101.0 | 317.8 | 28.8 | 1833.8 |
| Min | 5.5 | 4.2 | 2.3 | 0.3 | 1.5 | 0.2 | 4.1 | |
| Mean | 28.9 | 17.1 | 397.5 | 30.6 | 97.2 | 12.5 | 537.8 | |
| SD | 10.9 | 5.0 | 346.0 | 23.7 | 81.4 | 7.4 | 444.3 | |
| P. durangensis (130) | Max | 44.5 | 29.8 | 1027.5 | 89.2 | 312.0 | 38.4 | 1319.0 |
| Min | 8.7 | 6.9 | 8.9 | 1.3 | 3.0 | 0.4 | 14.4 | |
| Mean | 25.9 | 16.6 | 269.2 | 37.2 | 106.6 | 11.8 | 424.8 | |
| SD | 8.5 | 4.4 | 210.9 | 23.4 | 67.5 | 8.2 | 294.5 | |
| P. engelmannii (89) | Max | 51.8 | 27.8 | 1825.0 | 44.3 | 308.3 | 30.4 | 2024.7 |
| Min | 6.5 | 3.8 | 6.0 | 0.3 | 0.5 | 0.2 | 6.9 | |
| Mean | 27.7 | 15.0 | 507.5 | 18.5 | 107.0 | 11.8 | 644.7 | |
| SD | 9.4 | 5.4 | 421.7 | 11.5 | 58.9 | 6.9 | 479.5 | |
| P. leiophylla (84) | Max | 55.3 | 29.2 | 1129.8 | 89.7 | 392.0 | 21.8 | 1614.6 |
| Min | 8.4 | 5.4 | 5.7 | 0.5 | 1.1 | 0.4 | 7.7 | |
| Mean | 29.6 | 16.6 | 329.5 | 23.2 | 149.6 | 7.4 | 509.7 | |
| SD | 12.0 | 5.4 | 278.6 | 20.8 | 111.3 | 5.7 | 402.6 | |
| P. herrerae (97) | Max | 46.4 | 31.0 | 1056.9 | 72.8 | 139.4 | 31.8 | 1200.4 |
| Min | 5.0 | 5.2 | 4.0 | 0.3 | 0.8 | 0.4 | 5.4 | |
| Mean | 27.8 | 16.3 | 354.3 | 20.3 | 49.4 | 13.6 | 437.6 | |
| SD | 9.2 | 4.9 | 273.0 | 13.9 | 34.0 | 8.6 | 319.2 | |
| P. teocote (81) | Max | 45.0 | 24.7 | 789.3 | 47.9 | 161.3 | 34.8 | 975.0 |
| Min | 10.0 | 4.5 | 3.2 | 0.3 | 1.1 | 0.6 | 5.2 | |
| Mean | 29.6 | 15.6 | 288.9 | 16.6 | 50.5 | 12.2 | 368.2 | |
| SD | 9.4 | 4.0 | 207.6 | 12.2 | 38.3 | 7.9 | 261.7 | |
| P. lumholtzii (35) | Max | 42.0 | 24.9 | 832.2 | 37.6 | 86.9 | 37.0 | 981.5 |
| Min | 5.0 | 3.6 | 2.9 | 0.2 | 0.4 | 0.5 | 4.0 | |
| Mean | 22.4 | 14.7 | 236.3 | 10.7 | 32.0 | 11.7 | 290.7 | |
| SD | 8.6 | 4.3 | 209.7 | 8.5 | 27.1 | 10.2 | 252.7 | |
| P. strobiformis (98) | Max | 49.0 | 26.6 | 1240.1 | 37.6 | 211.3 | 40.4 | 1501.1 |
| Min | 5.0 | 6.3 | 1.8 | 0.3 | 1.5 | 0.4 | 4.0 | |
| Mean | 27.1 | 16.2 | 292.9 | 17.9 | 73.1 | 17.2 | 401.3 | |
| SD | 9.8 | 4.8 | 277.2 | 9.8 | 56.2 | 11.2 | 354.4 | |
| P. oocarpa (37) | Max | 35.7 | 18.7 | 448.9 | 49.5 | 173.1 | 38.0 | 647.5 |
| Min | 7.5 | 3.2 | 6.0 | 0.6 | 3.0 | 2.0 | 11.5 | |
| Mean | 21.6 | 12.9 | 153.4 | 19.6 | 53.4 | 16.8 | 243.3 | |
| SD | 6.3 | 3.6 | 109.6 | 12.4 | 42.0 | 9.8 | 167.3 | |
| P. douglasiana (30) | Max | 39.0 | 25.6 | 718.4 | 60.8 | 124.2 | 31.2 | 884.1 |
| Min | 8.9 | 5.6 | 2.7 | 0.5 | 1.1 | 0.3 | 4.6 | |
| Mean | 25.0 | 17.5 | 261.9 | 21.8 | 40.5 | 12.4 | 336.6 | |
| SD | 7.2 | 4.4 | 174.9 | 15.2 | 29.5 | 9.2 | 224.9 | |
| P. michoacana (32) | Max | 42.1 | 24.7 | 866.7 | 56.2 | 137.2 | 39.8 | 1073.5 |
| Min | 12.9 | 13.5 | 48.8 | 4.0 | 12.9 | 4.2 | 69.9 | |
| Mean | 31.3 | 20.3 | 416.6 | 27.4 | 64.6 | 21.4 | 530.0 | |
| SD | 8.2 | 3.0 | 232.5 | 13.5 | 36.6 | 9.2 | 285.6 | |
| Juniperus deppeana (48) | Max | 43.7 | 21.5 | 357.0 | 21.8 | 56.3 | 27.5 | 456.3 |
| Min | 10.0 | 4.5 | 3.7 | 0.5 | 1.2 | 0.8 | 6.2 | |
| Mean | 32.9 | 11.4 | 183.7 | 11.4 | 25.8 | 13.7 | 234.7 | |
| SD | 7.9 | 3.1 | 92.7 | 4.8 | 15.8 | 6.9 | 117.7 | |
| Arbutus bicolor (49) | Max | 44.8 | 2.5 | 236.1 | 9.6 | 231.1 | 14.5 | 375.6 |
| Min | 7.9 | 2.4 | 3.8 | 0.2 | 3.1 | 0.2 | 7.2 | |
| Mean | 22.8 | 8.9 | 83.3 | 3.4 | 48.5 | 5.4 | 140.6 | |
| SD | 7.8 | 2.9 | 53.9 | 2.4 | 46.8 | 3.7 | 97.6 | |
| Quercus sideroxyla (123) | Max | 57.0 | 24.8 | 1018.0 | 276.3 | 308.3 | 29.6 | 1559.0 |
| Min | 11.0 | 6.3 | 12.3 | 3.3 | 0.4 | 0.4 | 16.4 | |
| Mean | 30.7 | 14.6 | 290.9 | 87.3 | 75.6 | 8.5 | 462.2 | |
| SD | 9.7 | 3.7 | 213.4 | 90.7 | 62.2 | 6.7 | 331.9 | |
| Q. rugosa (61) | Max | 41.3 | 20.2 | 456.1 | 129.8 | 132.0 | 19.4 | 648.6 |
| Min | 9.3 | 3.3 | 4.2 | 1.6 | 3.2 | 1.5 | 14.3 | |
| Mean | 22.5 | 11.2 | 107.2 | 40.6 | 51.1 | 19.2 | 218.0 | |
| SD | 8.1 | 3.7 | 95.8 | 31.4 | 33.4 | 13.0 | 166.2 | |
| Q. durifolia (131) | Max | 45.5 | 22.1 | 930.9 | 118.4 | 421.9 | 63.9 | 1526.3 |
| Min | 7.0 | 5.4 | 6.4 | 1.3 | 2.1 | 0.3 | 10.2 | |
| Mean | 27.2 | 12.4 | 344.4 | 32.5 | 125.6 | 25.1 | 527.6 | |
| SD | 8.8 | 3.5 | 226.6 | 26.9 | 109.6 | 18.3 | 371.7 | |
| Q. crassifolia (108) | Max | 43.1 | 18.8 | 533.9 | 88.3 | 274.6 | 35.5 | 865.0 |
| Min | 8.5 | 6.0 | 14.3 | 1.3 | 11.0 | 1.0 | 33.4 | |
| Mean | 25.5 | 11.2 | 218.5 | 25.6 | 93.3 | 14.2 | 351.5 | |
| SD | 7.7 | 2.6 | 134.7 | 18.7 | 62.0 | 8.2 | 212.5 | |
| Tree Biomass Component Equation | Cross-Validation | Tree Biomass Component Equation | Cross-Validation | ||
|---|---|---|---|---|---|
| R2 | RMSE | R2 | RMSE | ||
| Pinus cooperi | P. douglasiana and P. michoacana | ||||
| 0.97 | 41.22 | 0.89 | 68.85 | ||
| 0.81 | 10.43 | 0.77 | 6.94 | ||
| 0.89 | 27.06 | 0.90 | 11.25 | ||
| 0.74 | 3.79 | 0.83 | 4.24 | ||
| 0.94 | 87.21 | 0.91 | 76.01 | ||
| P. durangensis | Juniperus depeanna | ||||
| 0.96 | 39.77 | 0.91 | 28.98 | ||
| 0.87 | 8.44 | 0.57 | 3.15 | ||
| 0.71 | 36.74 | 0.81 | 5.87 | ||
| 0.83 | 3.38 | 0.67 | 3.92 | ||
| 0.96 | 56.05 | 0.90 | 37.00 | ||
| P. engelmannii | Arbutus bicolor | ||||
| 0.91 | 127.64 | 0.93 | 15.11 | ||
| 0.98 | 1.64 | 0.87 | 0.85 | ||
| 0.59 | 37.66 | 0.95 | 6.99 | ||
| 0.70 | 3.77 | 0.79 | 1.72 | ||
| 0.92 | 140.91 | 0.95 | 22.81 | ||
| P. leiophylla | Quercus sideroxyla | ||||
| 0.94 | 68.35 | 0.90 | 65.75 | ||
| 0.89 | 6.92 | 0.79 | 27.87 | ||
| 0.63 | 67.38 | 0.80 | 27.31 | ||
| 0.81 | 2.53 | 0.74 | 3.45 | ||
| 0.92 | 110.21 | 0.92 | 96.83 | ||
| P. herrerae | Q. rugosa | ||||
| 0.82 | 114.23 | 0.90 | 29.42 | ||
| 0.72 | 7.37 | 0.77 | 15.14 | ||
| 0.72 | 18.01 | 0.86 | 12.56 | ||
| 0.60 | 5.41 | 0.84 | 5.21 | ||
| 0.85 | 124.12 | 0.94 | 41.8 | ||
| P. teocote | Q. durifolia | ||||
| 0.92 | 58.5 | 0.94 | 55.64 | ||
| 0.93 | 9.34 | 0.87 | 9.53 | ||
| 0.93 | 3.02 | 0.89 | 35.50 | ||
| 0.73 | 16.67 | 0.86 | 6.87 | ||
| 0.95 | 76.11 | 0.95 | 80.97 | ||
| P. lumholtzii | Q. crassifolia | ||||
| 0.91 | 65.02 | 0.83 | 56.45 | ||
| 0.76 | 16.98 | 0.81 | 8.12 | ||
| 0.83 | 4.21 | 0.82 | 25.96 | ||
| 0.82 | 4.14 | 0.64 | 4.92 | ||
| 0.90 | 83.32 | 0.91 | 64.58 | ||
| P. strobiformis | All pine species | ||||
| 0.90 | 88.01 | 0.92 | 72.12 | ||
| 0.88 | 6.34 | 0.63 | 11.65 | ||
| 0.93 | 15.24 | 0.61 | 31.05 | ||
| 0.72 | 5.55 | 0.52 | 5.98 | ||
| 0.93 | 93.02 | 0.93 | 83.04 | ||
| P. oocarpa | All oak species | ||||
| 0.93 | 27.12 | 0.78 | 94.25 | ||
| 0.87 | 4.44 | 0.65 | 27.68 | ||
| 0.87 | 15.32 | 0.57 | 53.64 | ||
| 0.83 | 3.96 | 0.29 | 11.95 | ||
| 0.94 | 39.66 | 0.82 | 134.12 | ||
| Species | Carbon Proportion | |||
|---|---|---|---|---|
| Wood | Bark | Leaves/Needles | Total (s.d) | |
| Pinus cooperi | 0.485 | 0.511 | 0.471 | 0.489 (0.0020) |
| Pinus durangensis | 0.489 | 0.531 | 0.487 | 0.505 (0.0030) |
| Pinus engelmannii | 0.497 | 0.531 | 0.494 | 0.507 (0.0020) |
| Pinus leiophylla | 0.495 | 0.540 | 0.515 | 0.516 (0.0023) |
| Pinus herrerae | 0.479 | 0.524 | 0.480 | 0.512 (0.0025) |
| Pinus teocote | 0.484 | 0.539 | 0.512 | 0.512 (0.0028) |
| Pinus lumholtzii | 0.485 | 0.532 | 0.492 | 0.501 (0.0027) |
| Pinus strobiformis | 0.494 | 0.533 | 0.492 | 0.506 (0.0023) |
| Pinus oocarpa | 0.469 | 0.528 | 0.472 | 0.490 (0.0033) |
| Pinus douglasiana | 0.483 | 0.525 | 0.494 | 0.500 (0.0021) |
| Pinus michoacana | 0.48 | 0.508 | 0.486 | 0.491 (0.0015) |
| Quercus sideroxyla | 0.462 | 0.462 | 0.477 | 0.467 (0.0009) |
| Quercus rugosa | 0.455 | 0.466 | 0.459 | 0.438 (0.0034) |
| Quercus durifolia | 0.463 | 0.428 | 0.451 | 0.448 (0.0018) |
| Quercus crassifolia | 0.460 | 0.416 | 0.459 | 0.445 (0.0025) |
| Juniperus depeanna | 0.527 | 0.438 | 0.496 | 0.487 (0.0045) |
| Arbutus bicolor | 0.467 | 0.378 | 0.479 | 0.441 (0.0055) |
| AGB | Carbon | |||||
|---|---|---|---|---|---|---|
| MLR | DR-DF | SVM | MLR | RD-RF | SVM | |
| R2 | 0.465 | 0.488 | 0.512 | 0.463 | 0.491 | 0.510 |
| RMSE | 51.044 | 49.971 | 49.771 | 25.282 | 24.630 | 24.727 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Larreta, B.; López-Sánchez, C.A.; Corral-Rivas, J.J.; López-Martínez, J.O.; Aguirre-Calderón, C.G.; Álvarez-González, J.G. Allometric Equations for Estimating Biomass and Carbon Stocks in the Temperate Forests of North-Western Mexico. Forests 2017, 8, 269. https://doi.org/10.3390/f8080269
Vargas-Larreta B, López-Sánchez CA, Corral-Rivas JJ, López-Martínez JO, Aguirre-Calderón CG, Álvarez-González JG. Allometric Equations for Estimating Biomass and Carbon Stocks in the Temperate Forests of North-Western Mexico. Forests. 2017; 8(8):269. https://doi.org/10.3390/f8080269
Chicago/Turabian StyleVargas-Larreta, Benedicto, Carlos Antonio López-Sánchez, José Javier Corral-Rivas, Jorge Omar López-Martínez, Cristóbal Gerardo Aguirre-Calderón, and Juan Gabriel Álvarez-González. 2017. "Allometric Equations for Estimating Biomass and Carbon Stocks in the Temperate Forests of North-Western Mexico" Forests 8, no. 8: 269. https://doi.org/10.3390/f8080269
APA StyleVargas-Larreta, B., López-Sánchez, C. A., Corral-Rivas, J. J., López-Martínez, J. O., Aguirre-Calderón, C. G., & Álvarez-González, J. G. (2017). Allometric Equations for Estimating Biomass and Carbon Stocks in the Temperate Forests of North-Western Mexico. Forests, 8(8), 269. https://doi.org/10.3390/f8080269