
Riparian Partial Harvesting and Upland Clear Cutting Alter Bird Communities in a Boreal Mixedwood Forest

Abstract
:1. Introduction
2. Materials and Methods
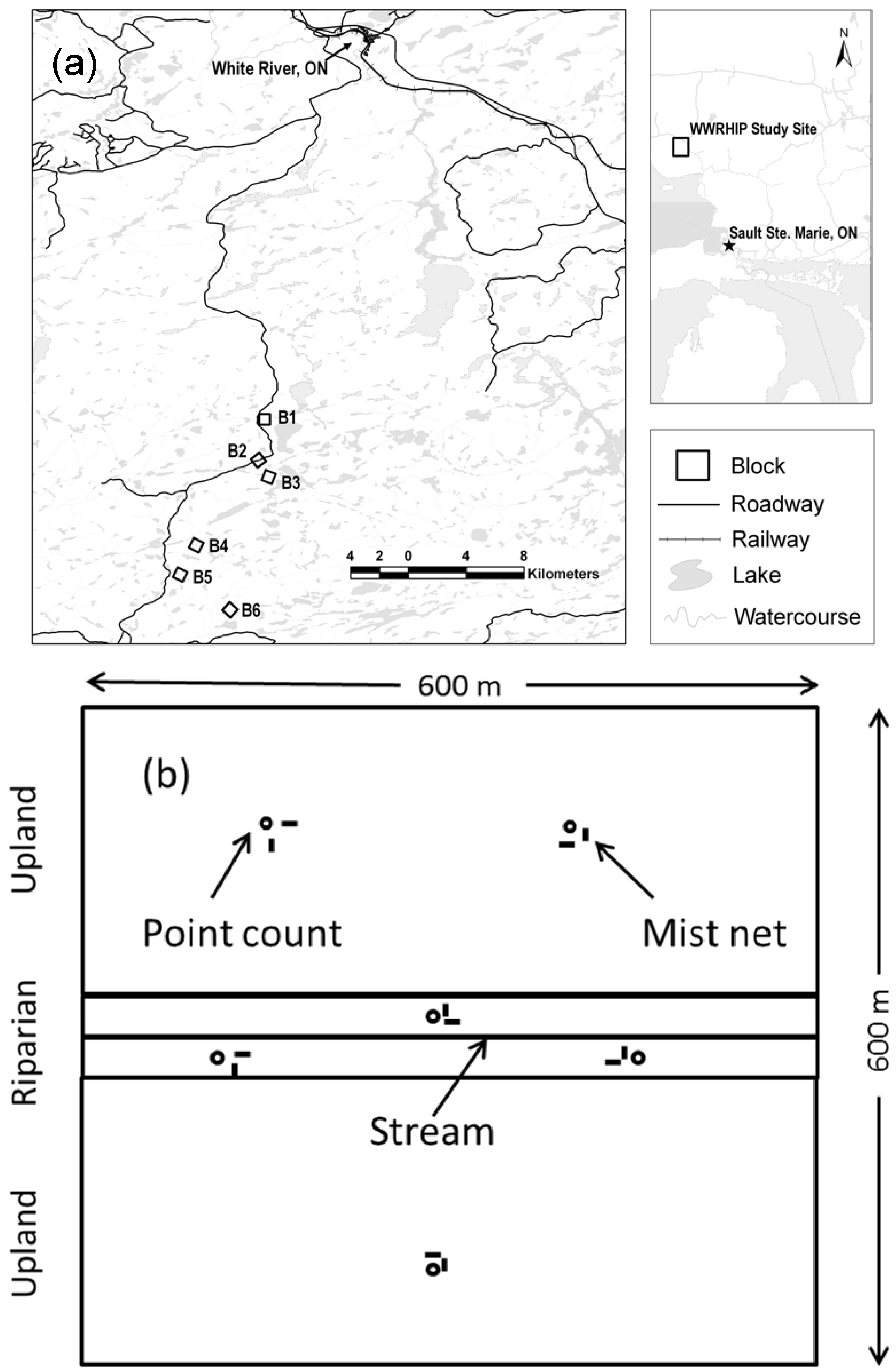
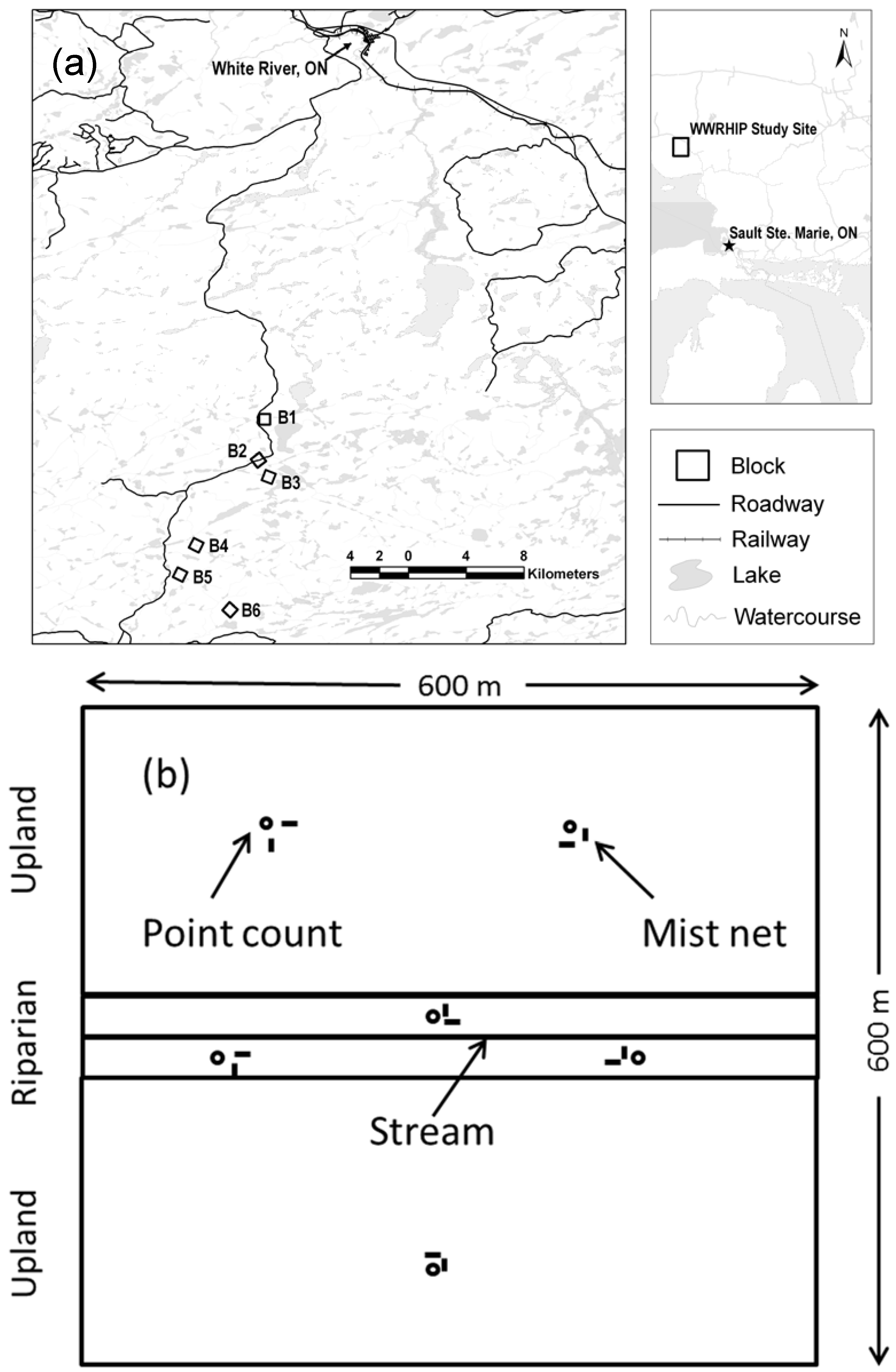
2.1. The Study Area and Plot Design
2.2. Bird Sampling
2.3. Data Analysis
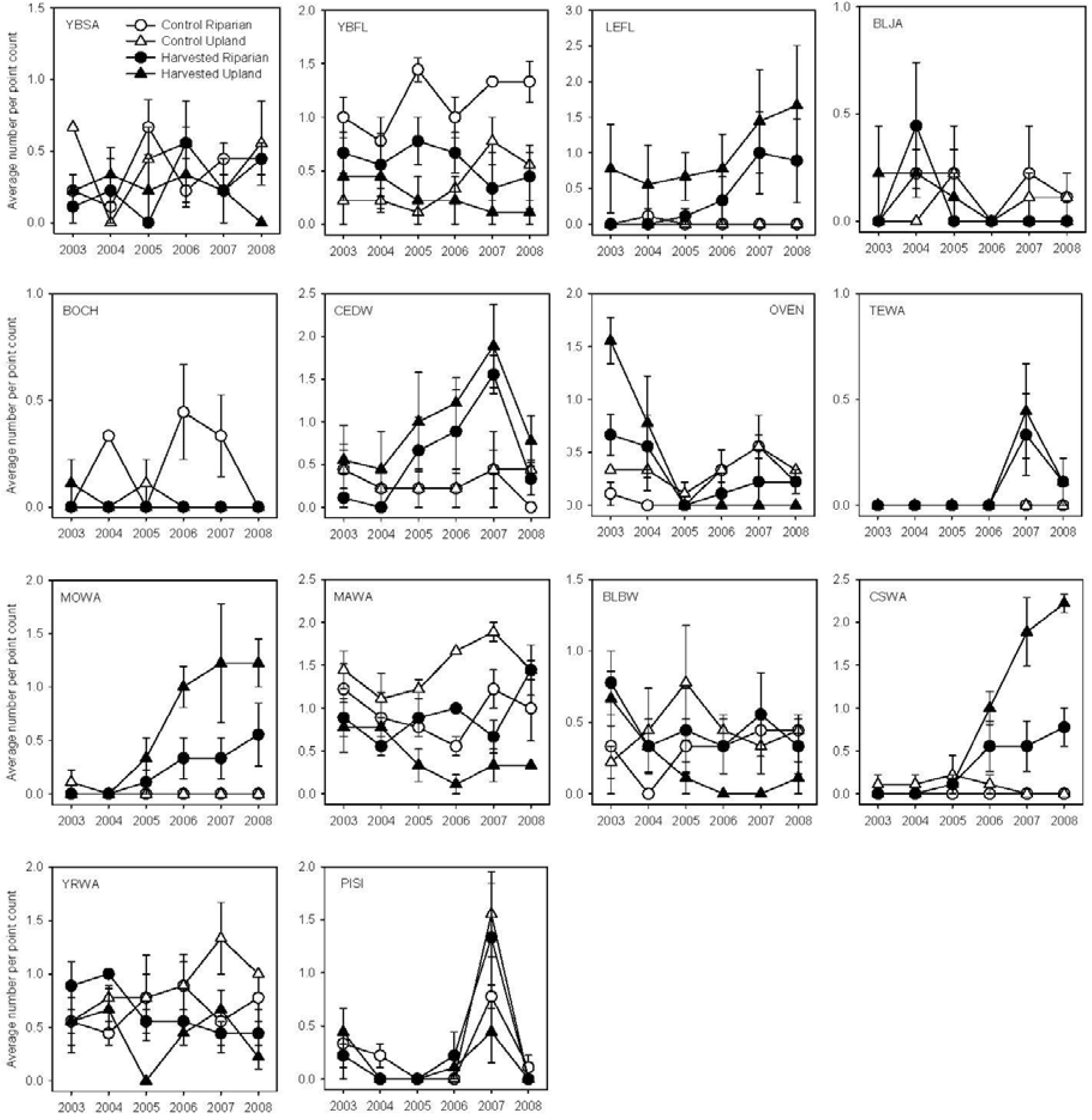
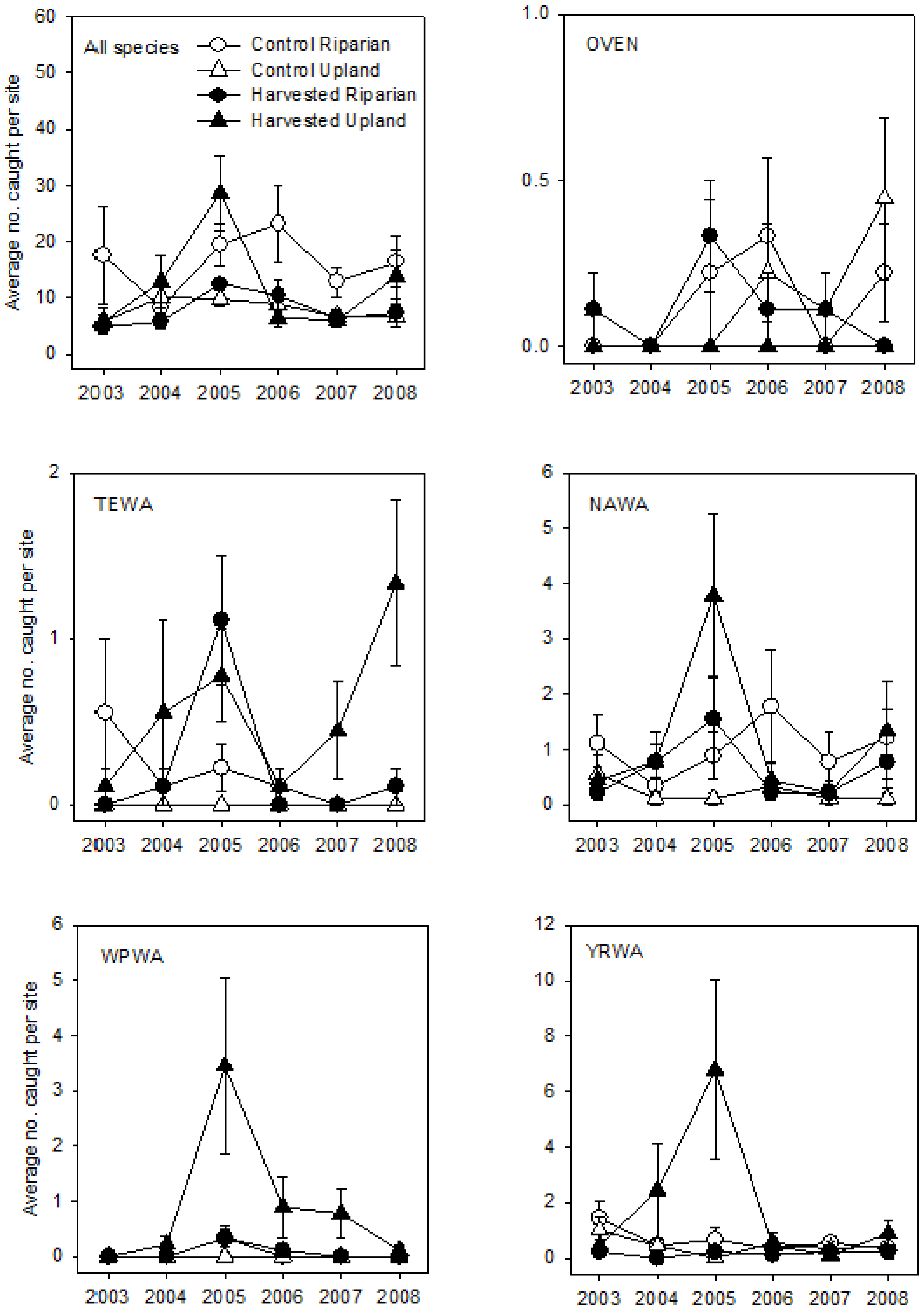
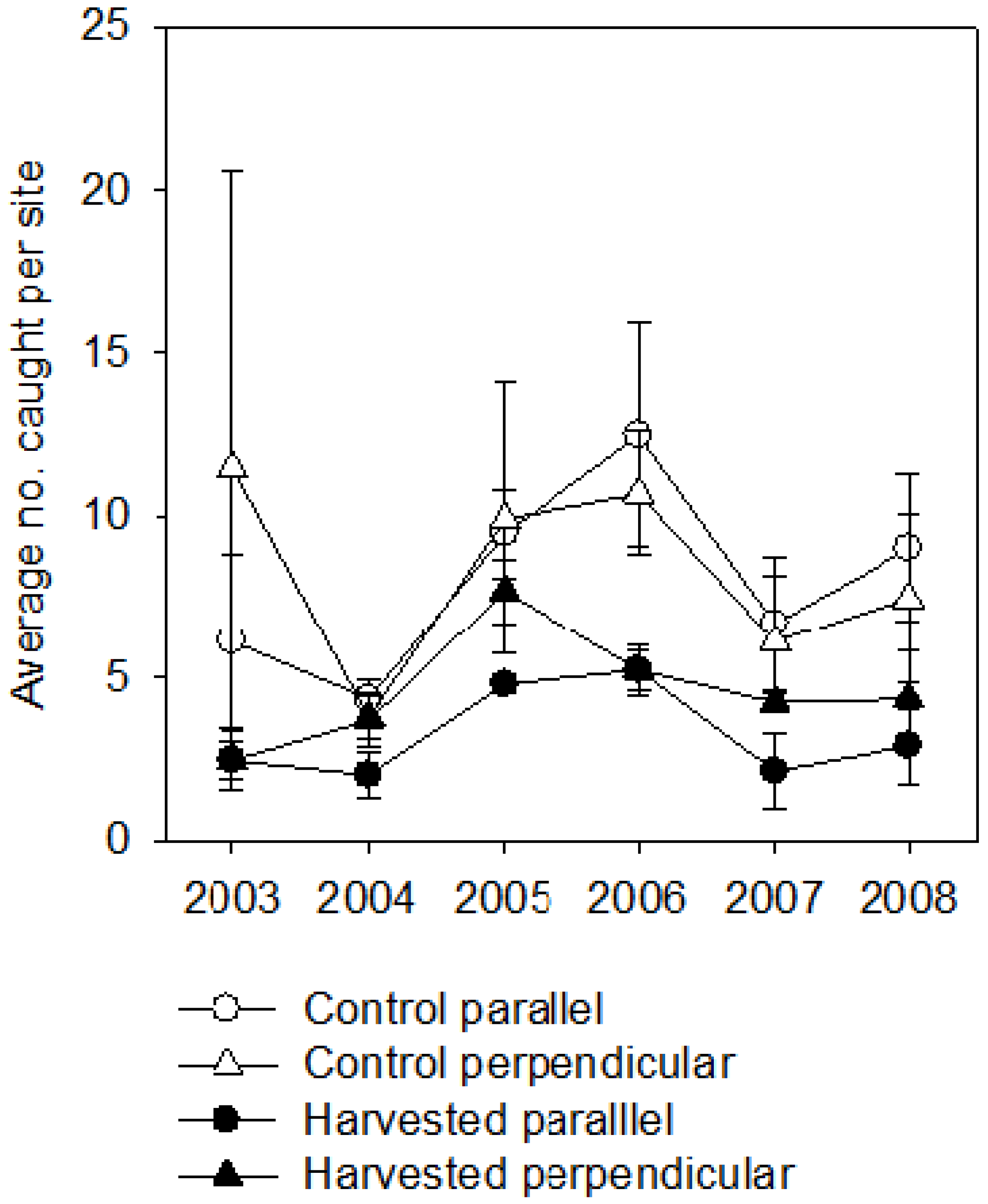
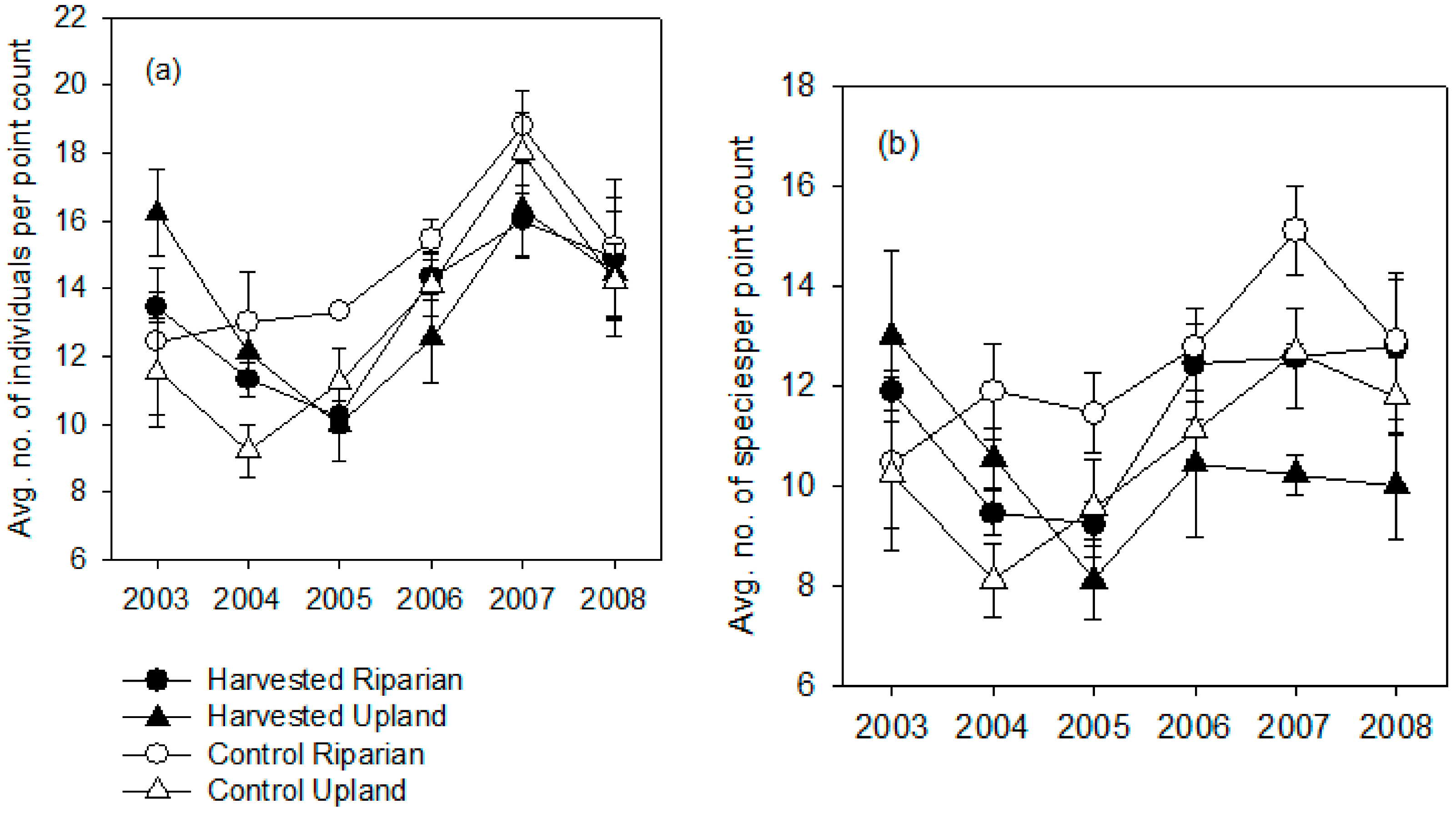
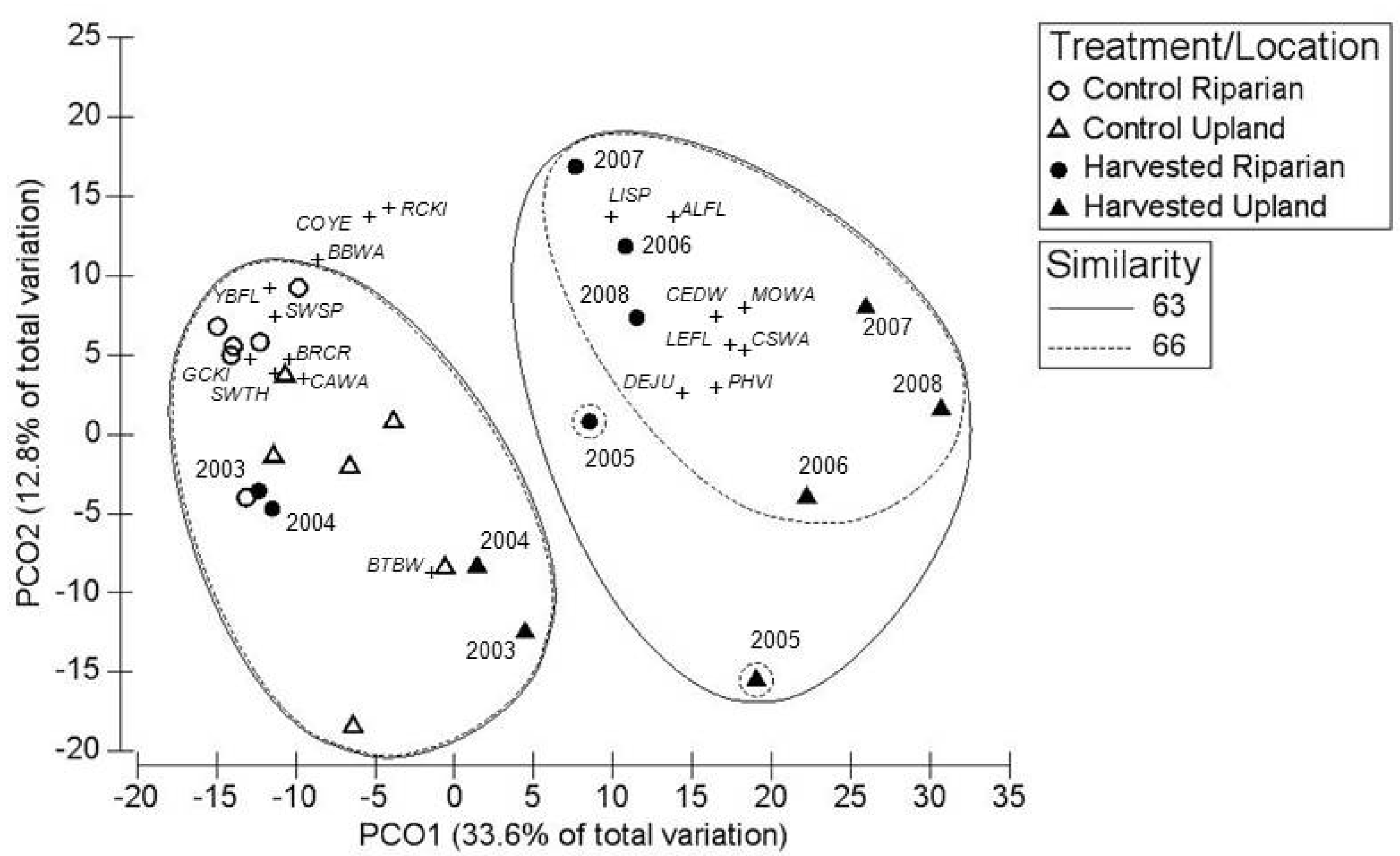
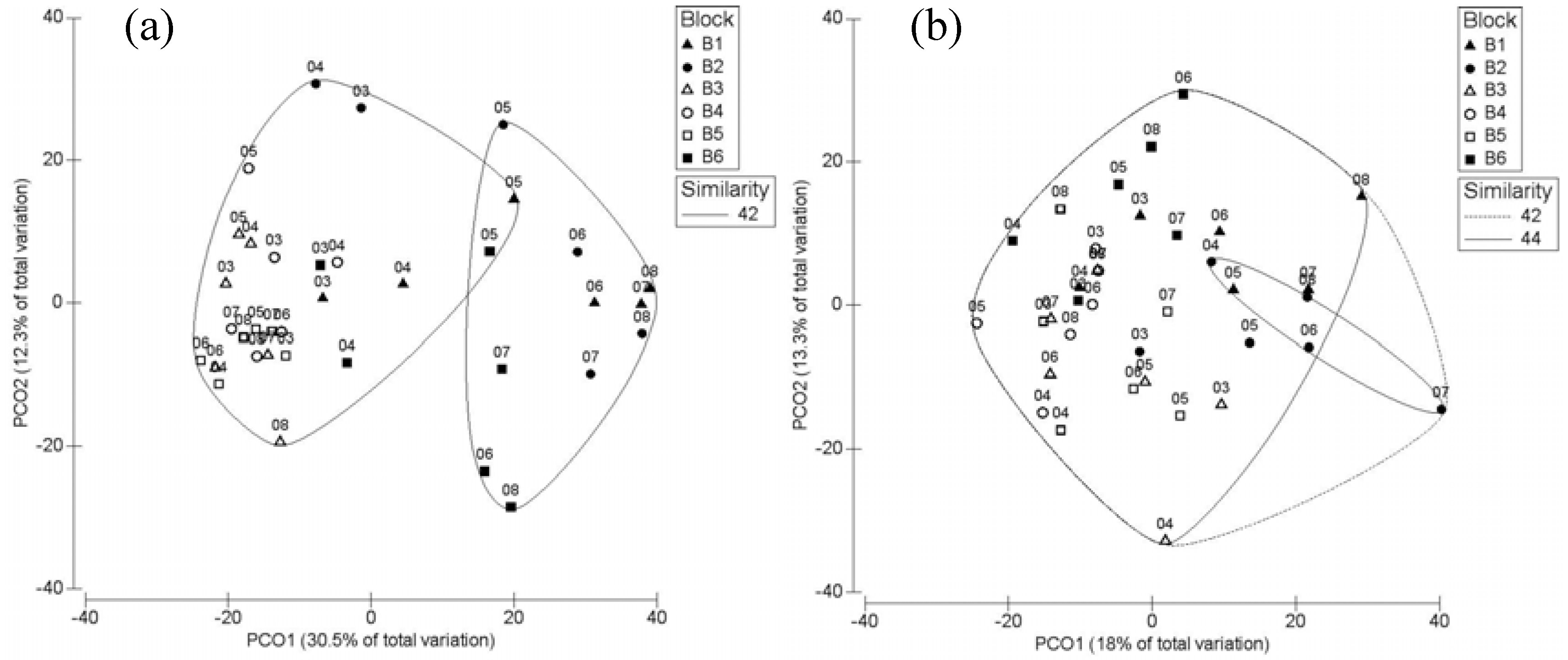
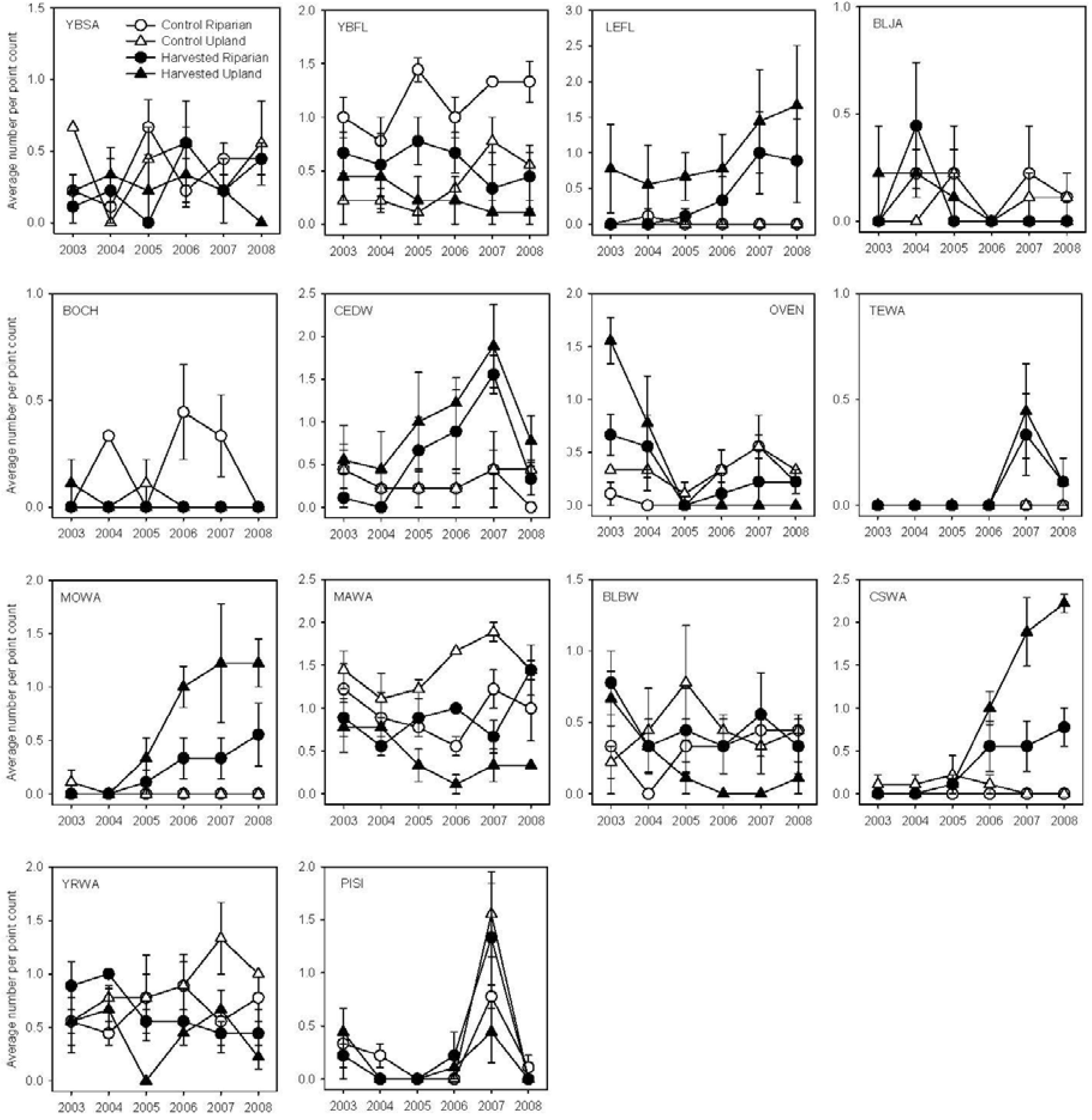
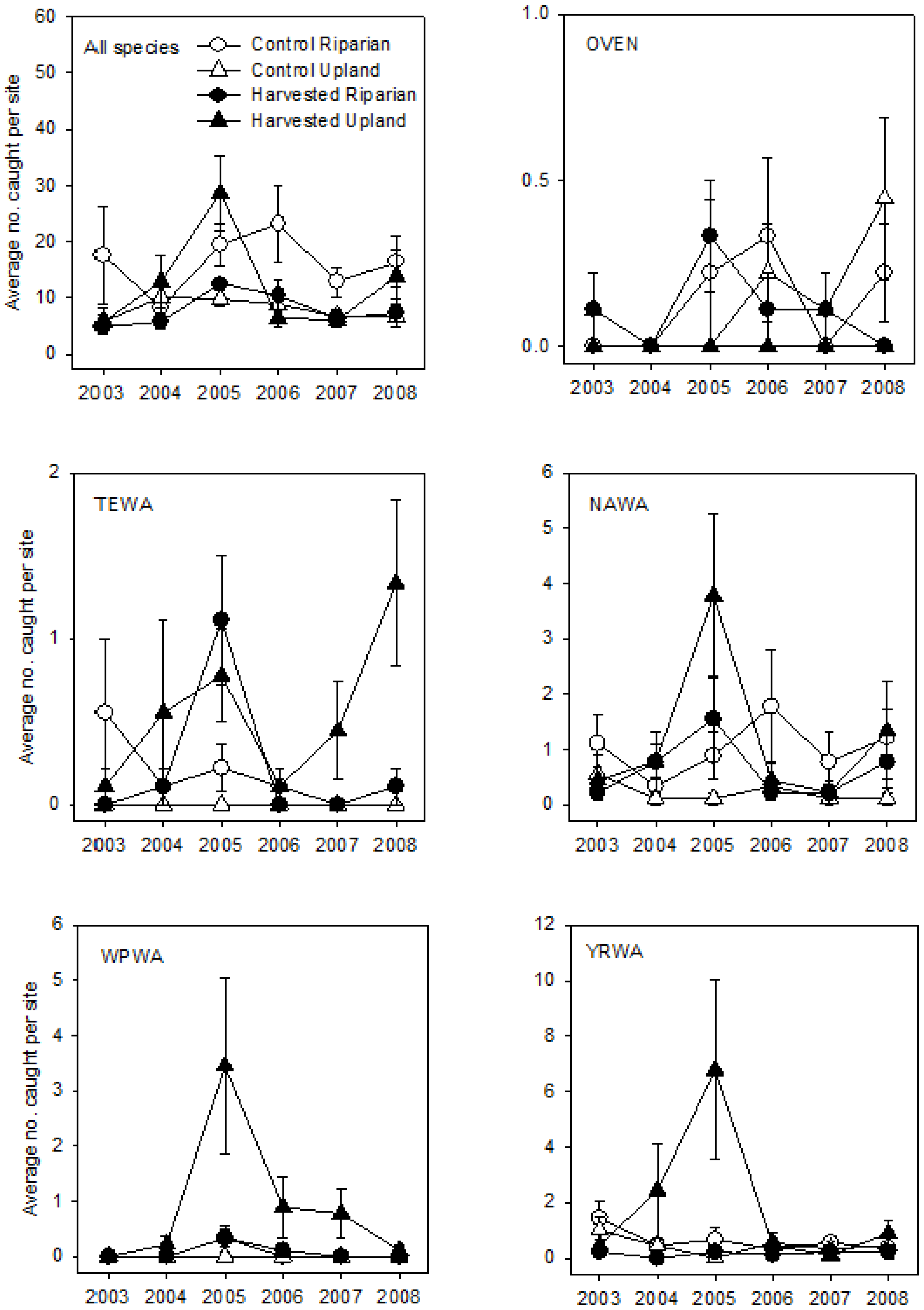
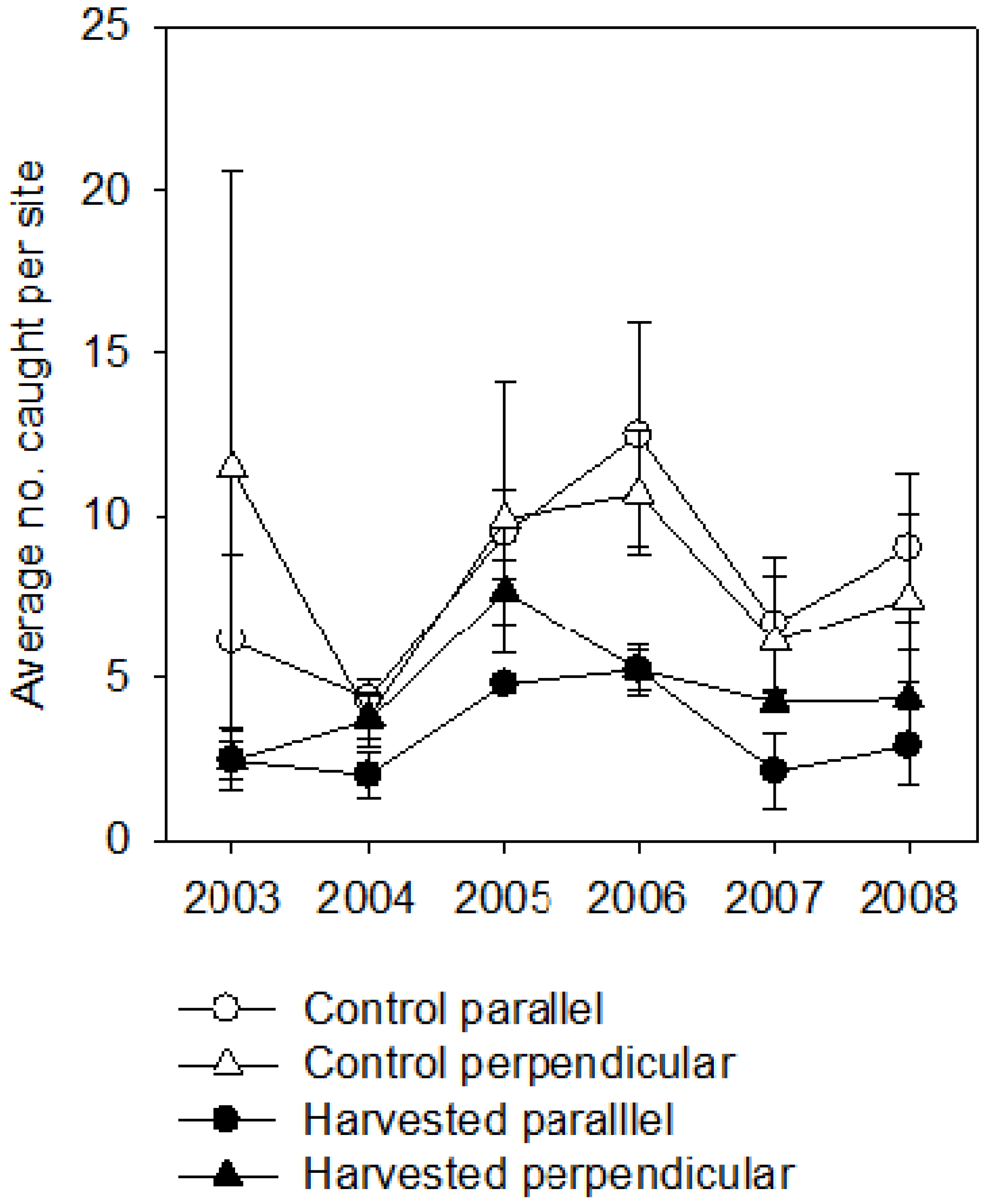
3. Results
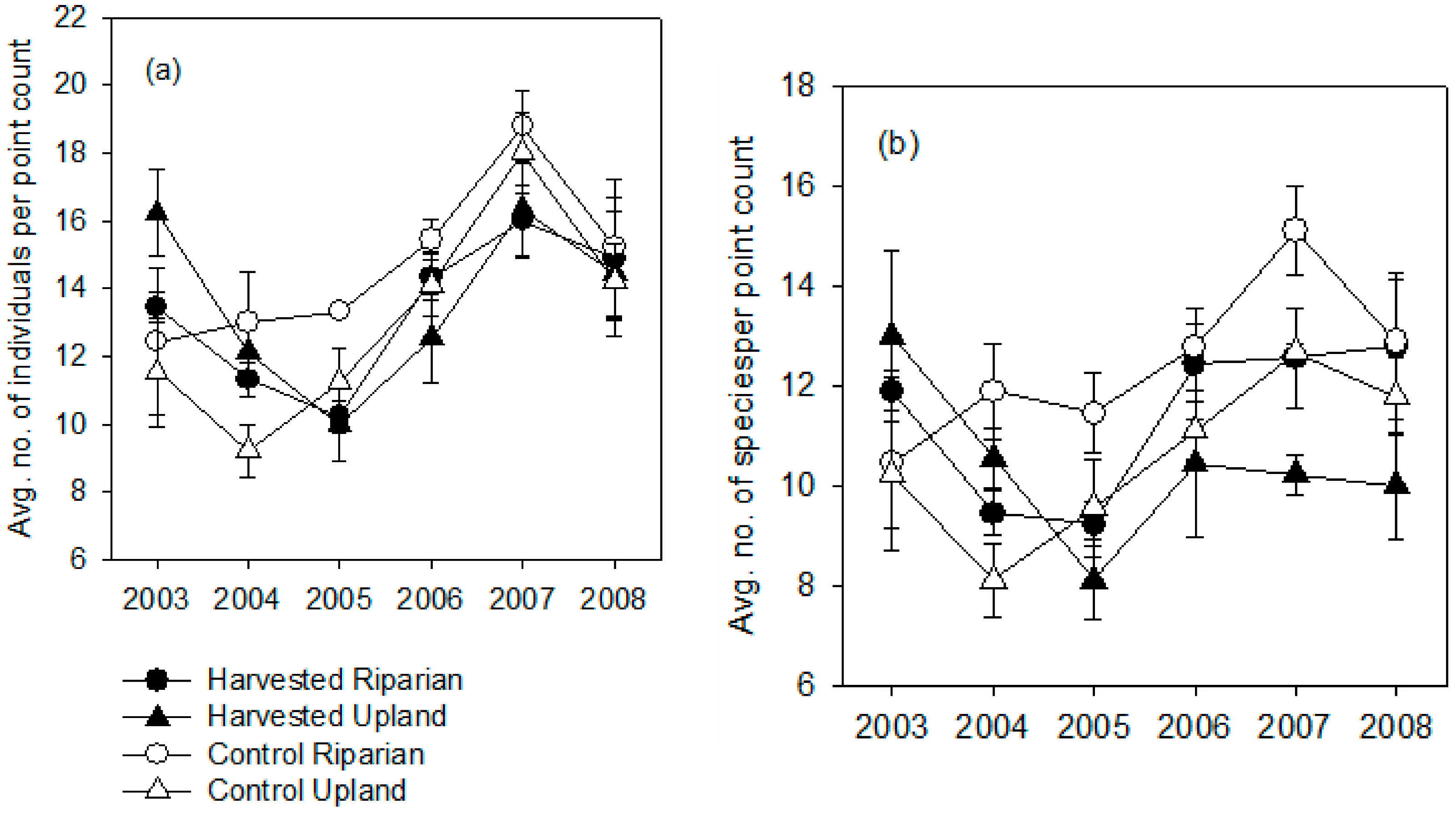
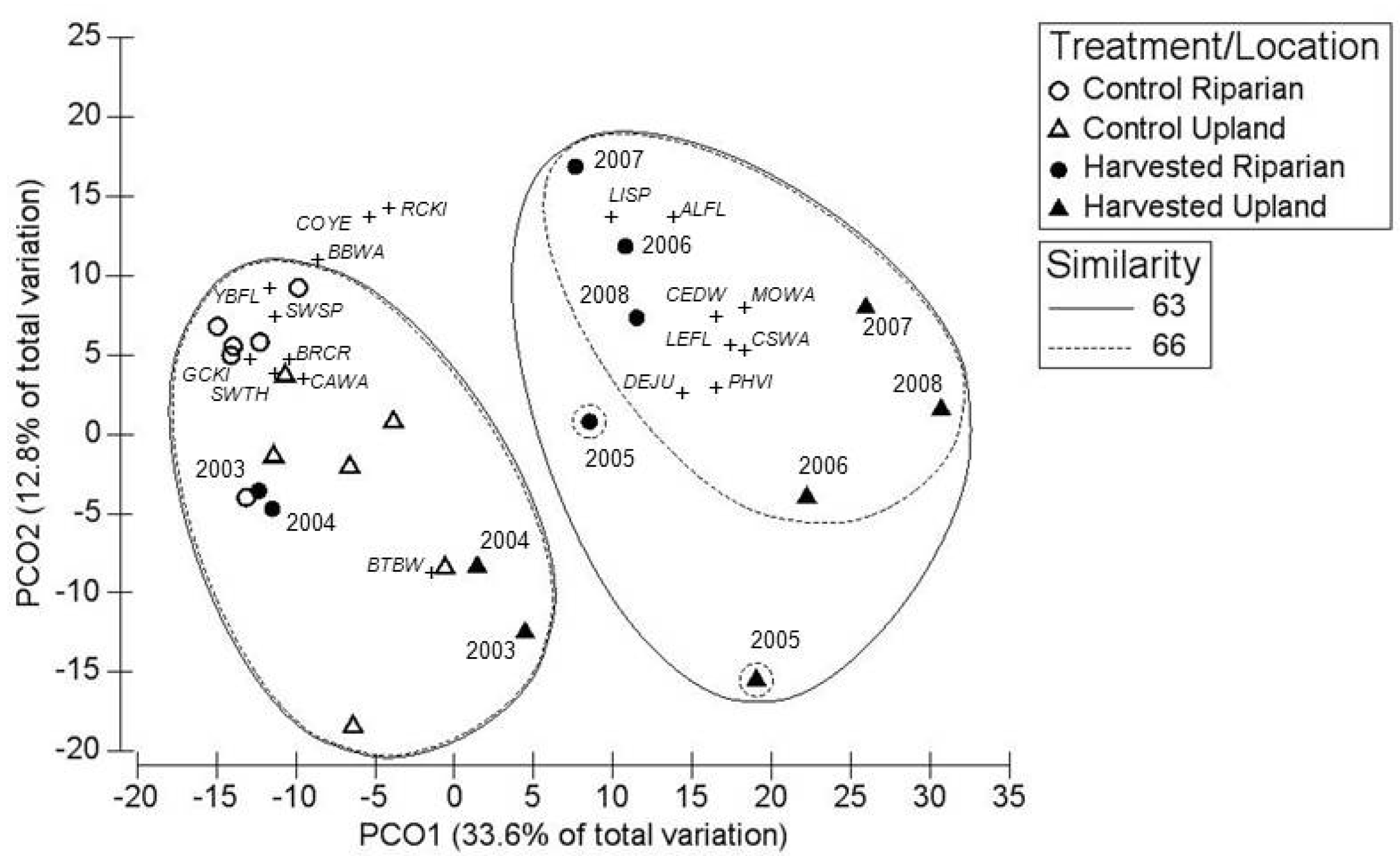
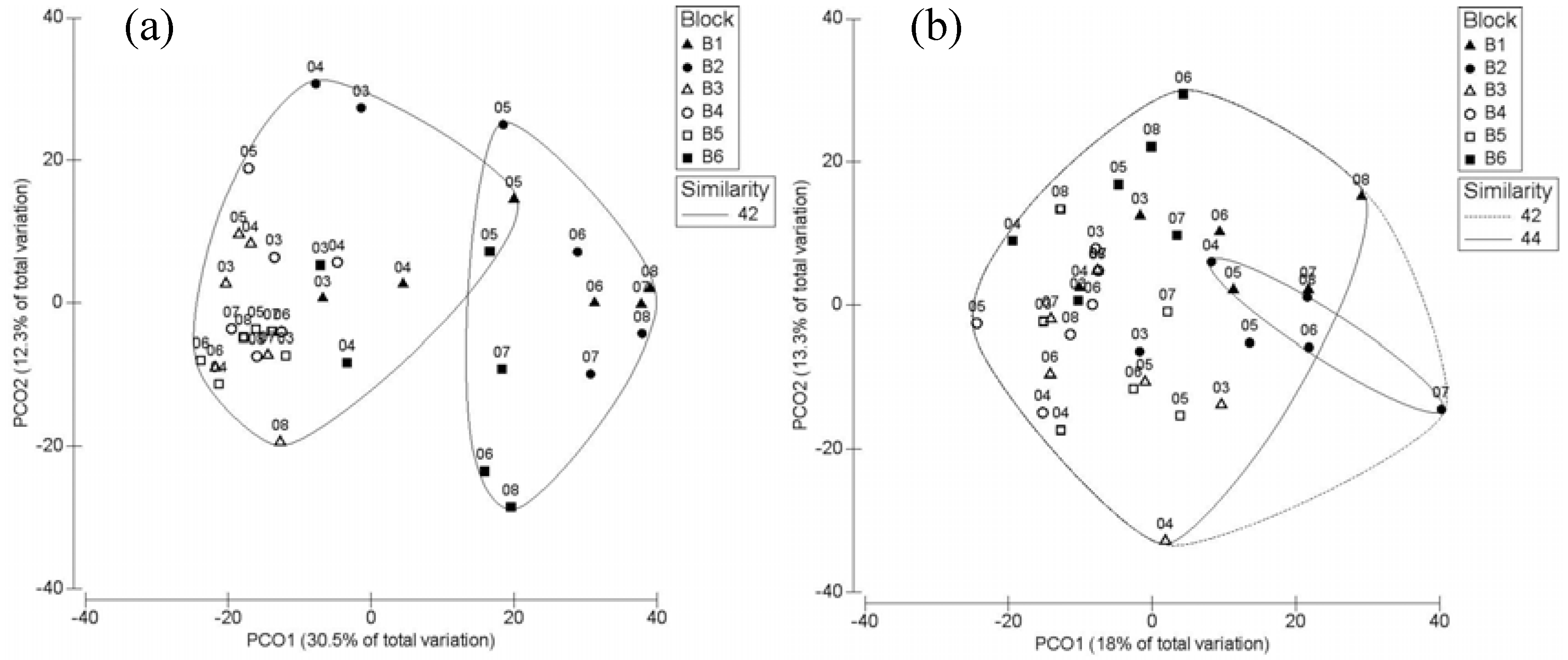
3.1. Breeding Season
3.2. Post-Breeding/Fall Migration
4. Discussion
4.1. Breeding Bird Response to Partial Harvesting
4.2. Caveats to Our Approach
4.3. Buffers as Travel Corridors
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Naiman, R.J.; Decamps, H.; Pollock, M. The role of riparian corridors in maintaining regional biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Pearson, S.F.; Giovanini, J.; Jones, J.E.; Kroll, A.J. Breeding bird community continues to colonize riparian buffers ten years after harvest. PLoS ONE 2015, 10, e0143241. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.F.; Nimmo, D.G.; Radford, J.Q. Riparian vegetation has disproportionate benefits for landscape-scale conservation of woodland birds in highly modified environments. J. Appl. Ecol. 2014, 51, 514–523. [Google Scholar] [CrossRef]
- Perry, R.W.; Wigley, T.B.; Melchiors, M.A.; Thill, R.E.; Tappe, P.A.; Miller, D.A. Width of riparian buffer and structure of adjacent plantations influence occupancy of conservation priority birds. Biodivers. Conserv. 2011, 20, 625–642. [Google Scholar] [CrossRef]
- LaRue, P.; Belanger, L.; Huot, J. Riparian edge effects on boreal balsam fir bird communities. Can. J. For. Res. 1995, 25, 555–566. [Google Scholar] [CrossRef]
- Mosley, E.; Holmes, S.B.; Nol, E. Songbird diversity and movement in upland and riparian habitats in the boreal mixedwood forest of northeastern Ontario. Can. J. For. Res. 2006, 36, 1149–1164. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Eaton, B.; Machtans, C.S.; Paszkowski, C.A.; Hannon, S.J.; Boutin, S. Is forest close to lakes ecologically unique? Analysis of vegetation, small mammals, amphibians and songbirds. For. Ecol. Manag. 2006, 223, 1–17. [Google Scholar] [CrossRef]
- Whitaker, D.M.; Montevecchi, W.A. Breeding bird assemblages associated with riparian, interior forest, and nonriparian edge habitats in a balsam fir ecosystem. Can. J. For. Res. 1997, 27, 1159–1167. [Google Scholar] [CrossRef]
- Whitaker, D.M.; Carroll, A.L.; Montevecchi, W.A. Elevated numbers of flying insects and insectivorous birds in riparian buffer strips. Can. J. Zool. 2000, 78, 740–747. [Google Scholar] [CrossRef]
- Hannon, S.J.; Paszkowski, C.A.; Boutin, S.; DeGroot, J.; Macdonald, S.E.; Wheatley, M.; Eaton, B.R. Abundance and species composition of amphibians, small mammals, and songbirds in riparian forest buffer strips of varying widths in the boreal mixedwood of Alberta. Can. J. For. Res. 2002, 32, 1784–1800. [Google Scholar] [CrossRef]
- Lee, P.; Smyth, C.; Boutin, S. Quantitative review of riparian buffer width guidelines from Canada and the United States. J. Environ. Manag. 2004, 70, 165–180. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H.; McClain, M.E. Riparia: Ecology, Conservation, and Management of Streamside Communities; Elsevier Academic Press: San Diego, CA, USA, 2005. [Google Scholar]
- Kardynal, K.J.; Morissette, J.L.; Van Wilgenburg, S.L.; Bayne, E.M.; Hobson, K.A. Avian responses to experimental harvest in southern boreal mixedwood shoreline forests: Implications for riparian buffer management. Can. J. For. Res. 2011, 41, 2375–2388. [Google Scholar] [CrossRef]
- Meiklejohn, B.A.; Hughes, J.W. Bird communities in riparian buffer strips of industrial forests. Am. Midl. Nat. 1999, 141, 172–184. [Google Scholar] [CrossRef]
- Whitaker, D.M.; Montevecchi, W.A. Breeding bird assemblages inhabiting riparian buffer strips in Newfoundland, Canada. J. Wildl. Manag. 1999, 63, 167–179. [Google Scholar] [CrossRef]
- Lambert, J.D.; Hannon, S.J. Short-term effects of timber harvest on abundance, territory characteristics, and pairing success of ovenbirds in riparian buffer strips. Auk 2000, 117, 687–698. [Google Scholar] [CrossRef]
- Darveau, M.; Beauchesne, P.; Belanger, L.; Huot, J.; LaRue, P. Riparian forest strips as habitat for breeding birds in boreal forest. J. Wildl. Manag. 1995, 59, 67–78. [Google Scholar] [CrossRef]
- Pearson, S.F.; Manuwal, D.A. Breeding bird response to riparian buffer width in managed Pacific northwest douglas-fir forests. Ecol. Appl. 2001, 11, 840–853. [Google Scholar] [CrossRef]
- Machtans, C.S.; Villard, M.A.; Hannon, S.J. Use of riparian buffer strips as movement corridors by forest birds. Conserv. Biol. 1996, 10, 1366–1379. [Google Scholar] [CrossRef]
- Robichaud, I.; Villard, M.A.; Machtans, C.S. Effects of forest regeneration on songbird movements in a managed forest landscape of Alberta, Canada. Landsc. Ecol. 2002, 17, 247–262. [Google Scholar] [CrossRef]
- Hannon, S.J.; Schmiegelow, F.K.A. Corridors may not improve the conservation value of small reserves for most boreal birds. Ecol. Appl. 2002, 12, 1457–1468. [Google Scholar] [CrossRef]
- Schmiegelow, F.K.A.; Machtans, C.S.; Hannon, S.J. Are boreal birds resilient to forest fragmentation? An experimental study of short-term community responses. Ecology 1997, 78, 1914–1932. [Google Scholar] [CrossRef]
- Buttle, J.M. Rethinking the donut: The case for hydrologically relevant buffer zones. Hydrol. Process. 2002, 16, 3093–3096. [Google Scholar] [CrossRef]
- Dragotescu, I.; Kneeshaw, D.D. A comparison of residual forest following fires and harvesting in boreal forests in Quebec, Canada. Silva Fenn. 2012, 46, 365–376. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Burgess, C.J.; Scimgeour, G.J.; Boutin, S.; Reedyk, S.; Kotak, B. Should riparian buffers be part of forest management based on emulation of natural disturbances? For. Ecol. Manag. 2004, 187, 185–196. [Google Scholar] [CrossRef]
- Johnson, E.A. Fire and Vegetation Dynamics: Studies from the North American Boreal Forest; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Andison, D.W.; McCleary, K. Disturbance in Riparian Zones on Foothills and Mountain Landscapes of Alberta; Foothills Model Forest: Hinton, AB, Canada, 2002; p. 40. [Google Scholar]
- Nitschke, C.R. Does forest harvesting emulate fire disturbance? A comparison of effects on selected attributes in coniferous-dominated headwater systems. For. Ecol. Manag. 2005, 214, 305–319. [Google Scholar] [CrossRef]
- Kardynal, K.J.; Hobson, K.A.; Van Wilgenburg, S.L.; Morissette, J.L. Moving riparian management guidelines towards a natural disturbance model: An example using boreal riparian and shoreline forest bird communities. For. Ecol. Manag. 2009, 257, 54–65. [Google Scholar] [CrossRef]
- Holmes, S.B.; Kreutzweiser, D.P.; Hamilton, P.S. Operational and economic feasibility of logging within forested riparian zones. For. Chron. 2010, 86, 601–607. [Google Scholar] [CrossRef]
- Rowe, J.S. Forest Regions of Canada; Canadian Forestry Service: Ottawa, ON, Canada, 1972. [Google Scholar]
- Ralph, C.J.; Droege, S.; Sauer, J.R. Managing and monitoring birds using point counts: Standards and applications. In Monitoring Bird Populations by Point Counts; Ralph, C.J., Sauer, J.R., Droege, S., Eds.; United States Department of Agriculture Forest Service: Albany, CA, USA, 1995; pp. 171–175. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Science B.V.: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zer-adjusted bray-curtis coefficient for denuded assemblages. J. Exp. Mar. Biol. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–281. [Google Scholar] [CrossRef]
- Cormack, R.M. A review of classification. J. R. Stat. Soc. Ser. A 1971, 134, 321–367. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric mutlivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. Permanova + for Primer: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymount, UK, 2008; p. 214. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef]
- Moran, M.D. Arguments for rejecting the sequential bonferroni in ecological studies. Oikos 2003, 100, 403–405. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; Lawrence Earlbaum Associates Inc.: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Chizinski, C.J.; Peterson, A.; Hanowski, J.; Blinn, C.; Vondracek, B.; Niemi, G. Breeding bird response to partially harvested riparian management zones. For. Ecol. Manag. 2011, 261, 1892–1900. [Google Scholar] [CrossRef]
- Johnson, W.N., Jr.; Brown, P.W. Avian use of a lakeshore buffer strip and an undisturbed lakeshore in maine. North. J. Appl. For. 1990, 7, 114–117. [Google Scholar]
- Hanowski, J.; Danz, N.; Lind, J.; Niemi, G. Breeding bird response to riparian harvest and harvest equipment. For. Ecol. Manag. 2003, 174, 315–328. [Google Scholar] [CrossRef]
- Hanowski, J.; Danz, N.; Lind, J. Breeding bird response to riparian forest managment: 9 years post-harvest. For. Ecol. Manag. 2007, 241, 272–277. [Google Scholar] [CrossRef]
- Hanowski, J.; Danz, N.; Lind, J.; Niemi, G. Breeding bird response to varying amounts of basal area retention in riparian buffers. J. Wildl. Manag. 2005, 69, 689–698. [Google Scholar] [CrossRef]
- Marczak, L.B.; Sakamaki, T.; Turvey, S.L.; Deguise, I.; Wood, S.L.R.; Richardson, J.S. Are forest buffers an effective conservation strategy for riparian fauna? An assessment using meta-analysis. Ecol. Appl. 2010, 20, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Celis-Murillo, A.; Deppe, J.L.; Allen, M.F. Using soundscape recordings to estimate bird species abundance, richness, and composition. J. Field Ornithol. 2009, 80, 64–78. [Google Scholar] [CrossRef]
- Anderson, M.J. The need to get the basics right in wildlife fiedl studies. Wildl. Soc. Bull. 2001, 29, 1294–1297. [Google Scholar]
- Rempel, R.S.; Hobson, K.A.; Holborn, G.; Van Wilgenburg, S.L.; Elliott, J. Bioacoustic monitoring of forest songbirds: Interpreter variability and effects of configuration and digital processing methods in the laboratory. J. Field Ornithol. 2005, 76, 1–11. [Google Scholar] [CrossRef]
- Hutto, R.L.; Stutzman, R.J. Human versus autonomous recording units: A comparison of point-count results. J. Field Ornithol. 2009, 80, 387–398. [Google Scholar] [CrossRef]
- Campbell, M.; Frnacis, C.M. Using stereo-microphones to evaluate observer variation in North American breeding bird survey point counts. Auk 2011, 128, 303–312. [Google Scholar] [CrossRef]
- Smith, E.P.; Orvos, D.R.; Cairns, J., Jr. Impact assessment using the Before-After-Control-Impact (BACI) model: Concerns and comments. Can. J. Fish. Aquat. Sci. 1993, 50, 627–637. [Google Scholar] [CrossRef]
- Shirley, S.M.; Smith, J.N.M. Bird community structure across riparian buffer strips of varying width in a coastal temperate forest. Biol. Conserv. 2005, 125, 475–489. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zone | Riparian Buffer Width (m) | Pre-Harvest Basal Area (m2/ha) | Residual Basal Area (m2/ha) | Basal Area Removed (%) |
|---|---|---|---|---|
| B1-North | 40 (30–70) a | 30.9 | 24.4 | 21 |
| B1-South | 30 (30–50) | 34.3 | 27.5 | 20 |
| B2-North | 30 (30–90) | 34.6 | 26.3 | 24 |
| B2-South | 30 (30–50) | 29.0 | 19.3 | 33 |
| B3 | NA b | NA | NA | 0 |
| B4 | NA | NA | NA | 0 |
| B5 | NA | NA | NA | 0 |
| B6-East | 50 (50–70) | 29.9 | 23.4 | 22 |
| B6-West | NA | NA | NA | 0 |
| p-Value | Cohen’s d a | ||||||
|---|---|---|---|---|---|---|---|
| Riparian/Upland | Riparian | Upland | Riparian | Upland | |||
| Species b | n c | Tr × Yr | Lo × Tr × Yr d | Tr × Yr | Tr × Yr | ||
| YBSA | 104 | 0.495 | 0.030 | 0.020 | 0.129 | −0.4 | −0.6 |
| YBFL | 176 | 0.001 | 0.926 | - | - | −1.2 | −0.6 |
| LEFL | 101 | 0.023 | 0.988 | - | - | 0.8 | 1.1 |
| BLJA | 31 | 0.003 | 0.980 | - | - | −0.4 | −0.3 |
| BOCH | 18 | 0.534 | 0.042 | 0.040 | 0.324 | −0.3 | −0.1 |
| CEDW | 142 | 0.001 | 0.991 | - | - | 0.8 | 1.0 |
| OVEN | 77 | 0.001 | 0.259 | - | - | −0.3 | −1.0 |
| TEWA | 12 | 0.012 | 0.469 | - | - | 0.4 | 0.3 |
| MOWA | 55 | 0.053 | 0.024 | 0.188 | 0.003 | 0.9 | 1.7 |
| MAWA | 295 | 0.884 | 0.008 | 0.160 | 0.011 | 0.2 | −2.1 |
| BLBW | 98 | 0.030 | 0.667 | - | - | 0.1 | −1.0 |
| CSWA | 98 | 0.007 | 0.006 | 0.014 | <0.001 | 1.2 | 1.9 |
| YRWA | 209 | 0.019 | 0.870 | - | - | −0.5 | −1.2 |
| PISI | 68 | 0.967 | 0.022 | 0.356 | 0.004 | 0.1 | −0.2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holmes, S.B.; McIlwrick, K.A.; Kreutzweiser, D.P.; Venier, L.A. Riparian Partial Harvesting and Upland Clear Cutting Alter Bird Communities in a Boreal Mixedwood Forest. Forests 2017, 8, 141. https://doi.org/10.3390/f8050141
Holmes SB, McIlwrick KA, Kreutzweiser DP, Venier LA. Riparian Partial Harvesting and Upland Clear Cutting Alter Bird Communities in a Boreal Mixedwood Forest. Forests. 2017; 8(5):141. https://doi.org/10.3390/f8050141
Chicago/Turabian StyleHolmes, Stephen B., Ken A. McIlwrick, David P. Kreutzweiser, and Lisa A. Venier. 2017. "Riparian Partial Harvesting and Upland Clear Cutting Alter Bird Communities in a Boreal Mixedwood Forest" Forests 8, no. 5: 141. https://doi.org/10.3390/f8050141
APA StyleHolmes, S. B., McIlwrick, K. A., Kreutzweiser, D. P., & Venier, L. A. (2017). Riparian Partial Harvesting and Upland Clear Cutting Alter Bird Communities in a Boreal Mixedwood Forest. Forests, 8(5), 141. https://doi.org/10.3390/f8050141