
Hydrology of a Water‐Limited Forest under Climate Change Scenarios: The Case of the Caatinga Biome, Brazil

Abstract
:1. Introduction
2. Materials and Methods
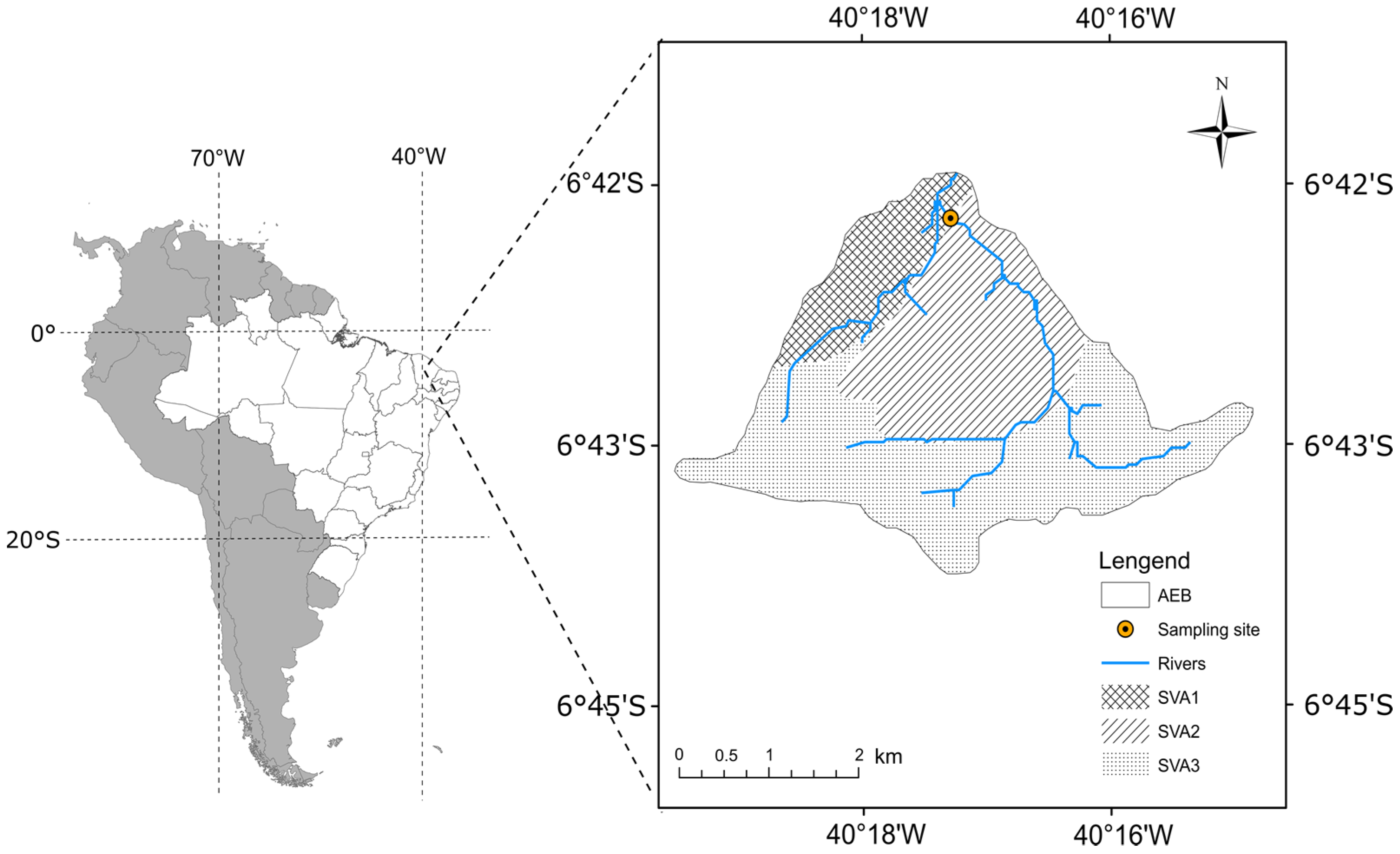
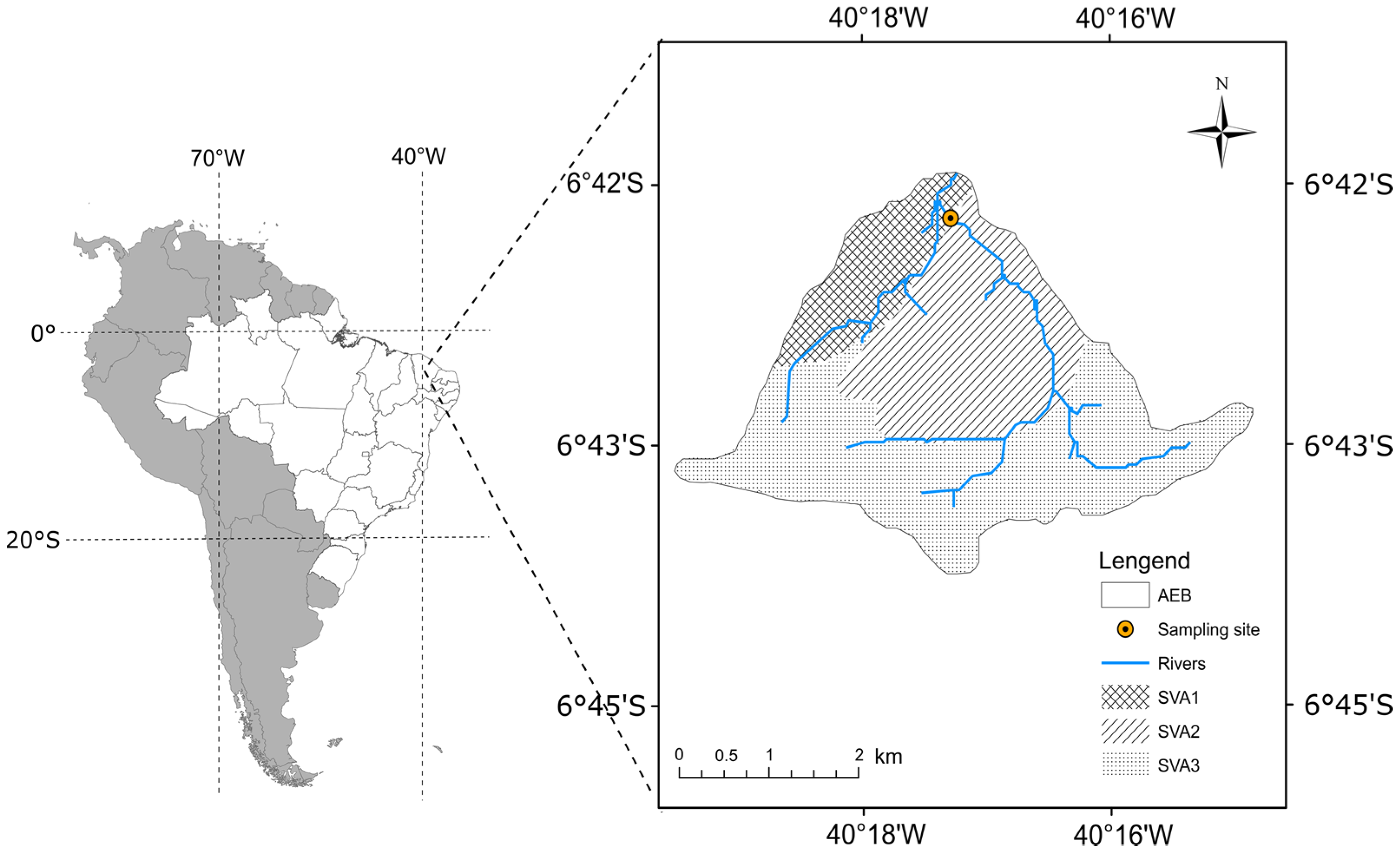
2.1. Study Area
2.2. Climatic Data Generator
2.3. Climate Change Scenario
2.4 Modeling
3. Results
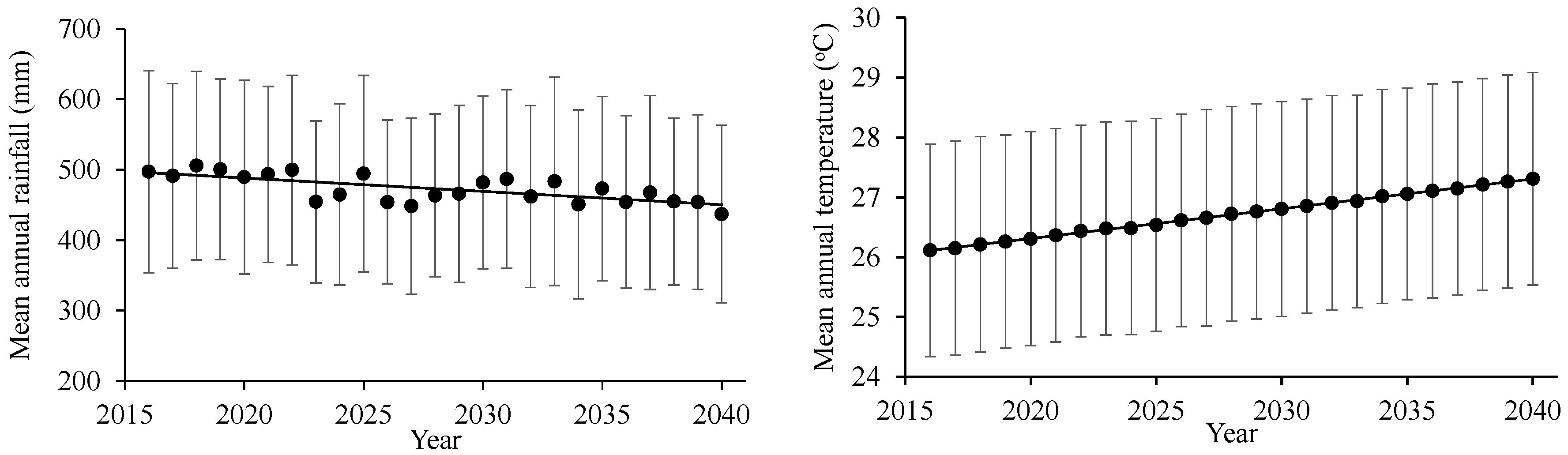
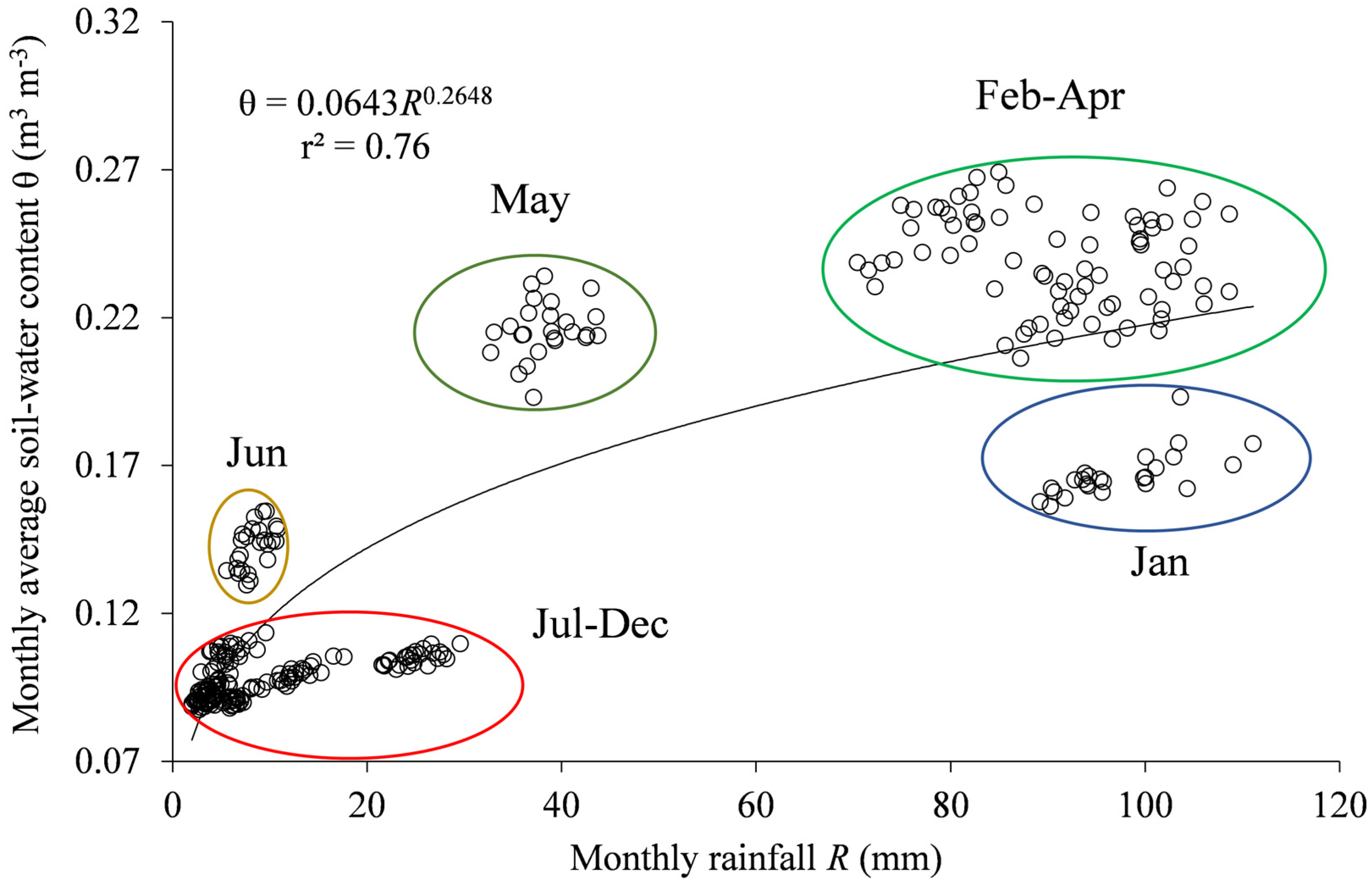
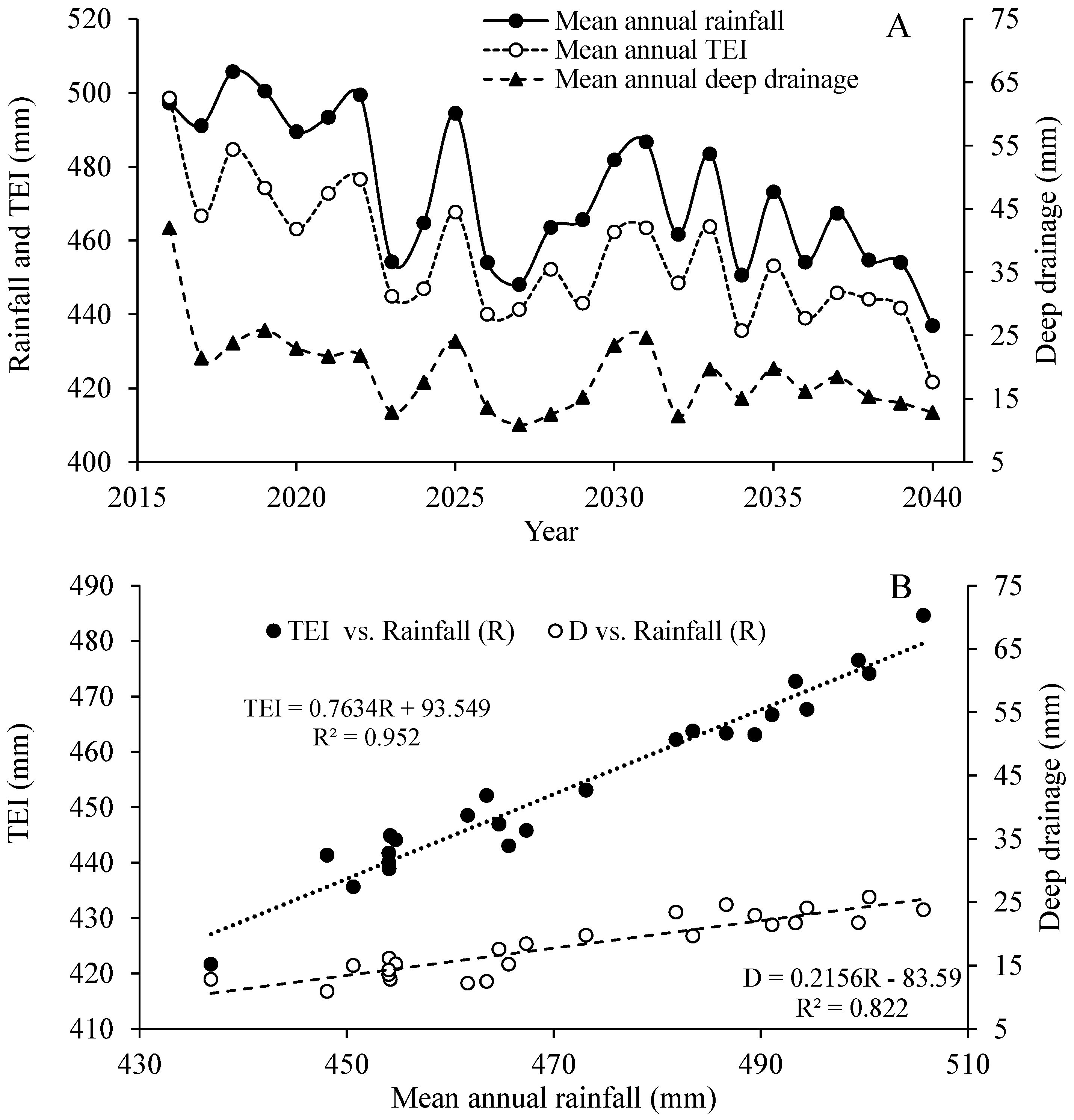
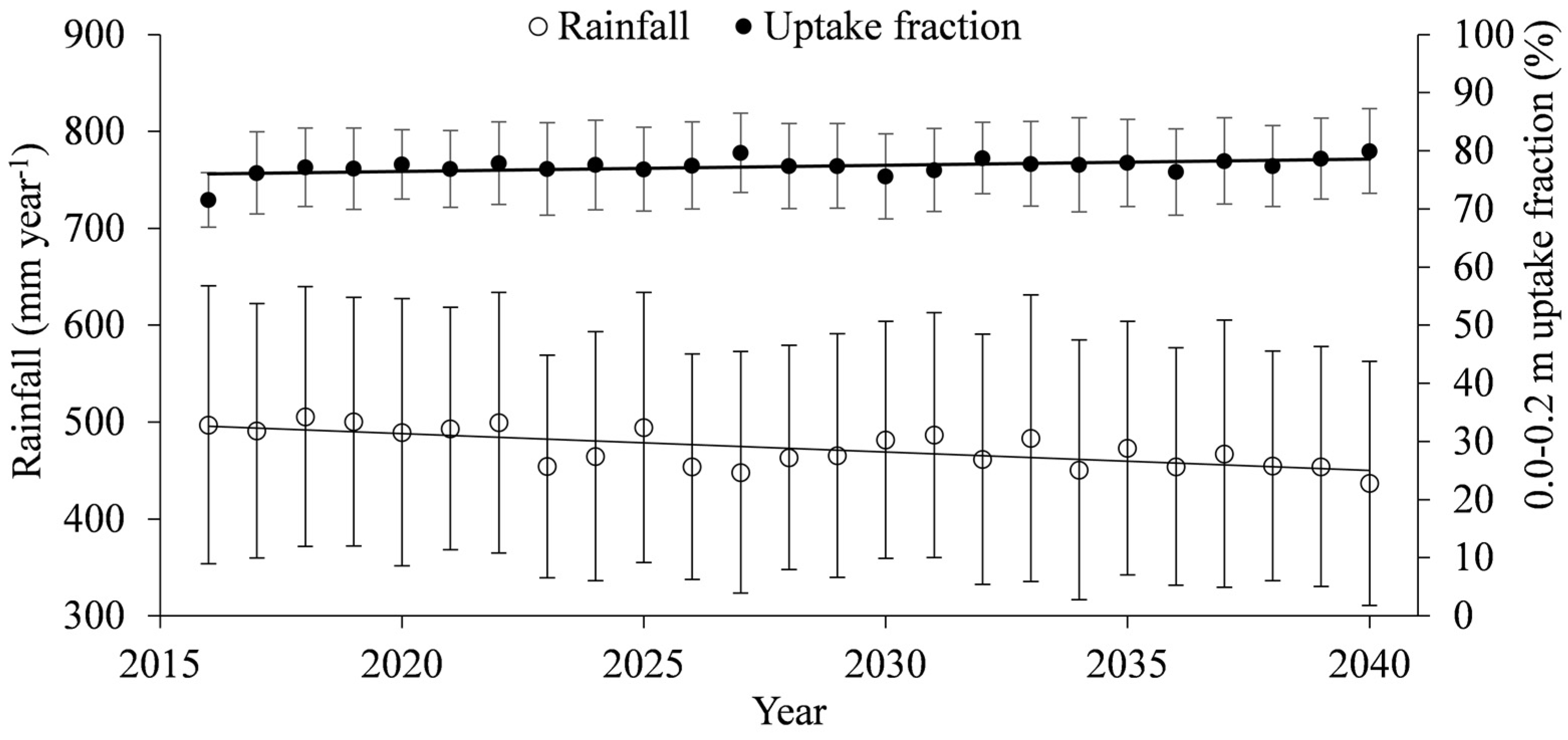
3.1. Generated Climatic Data
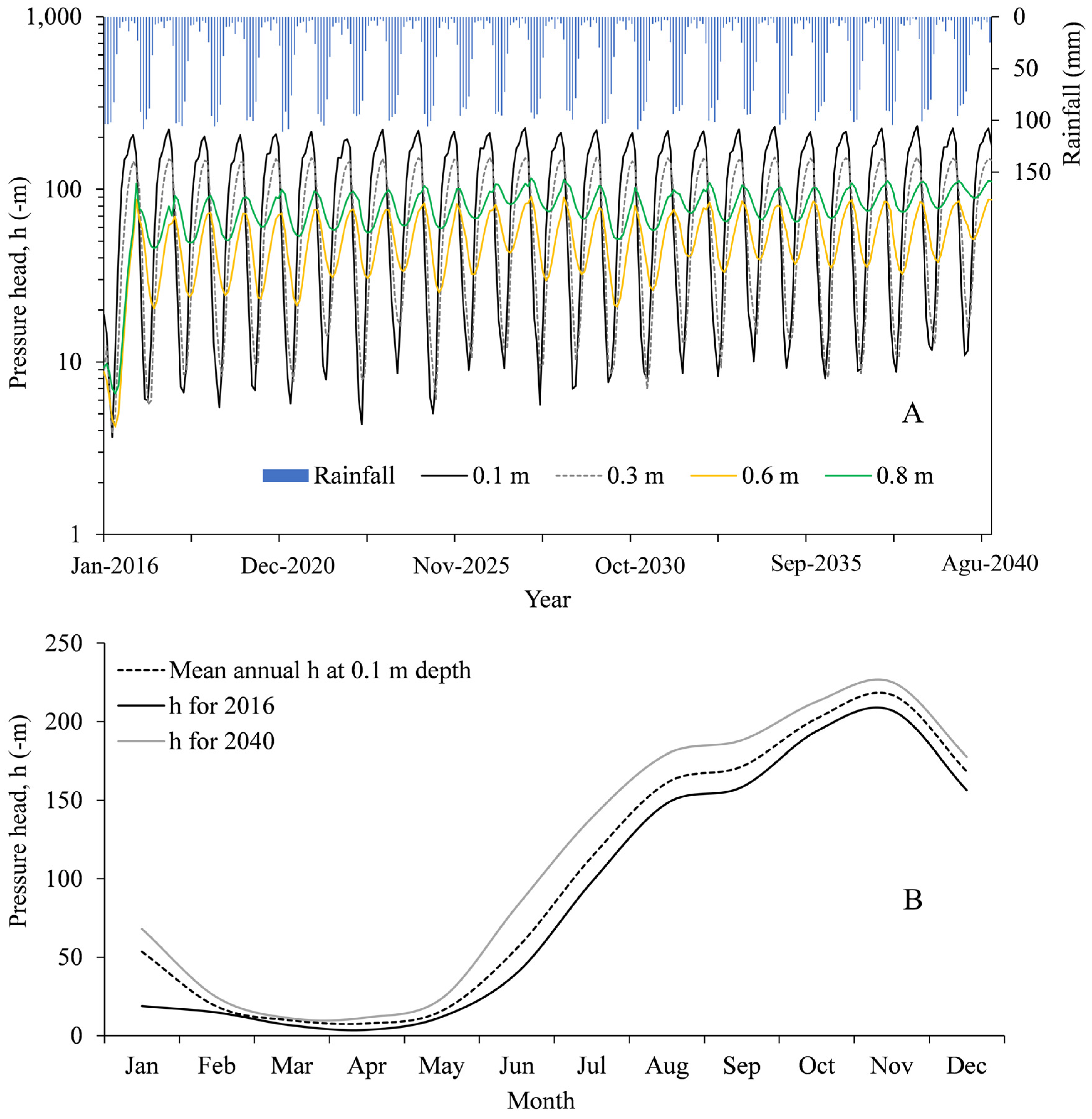
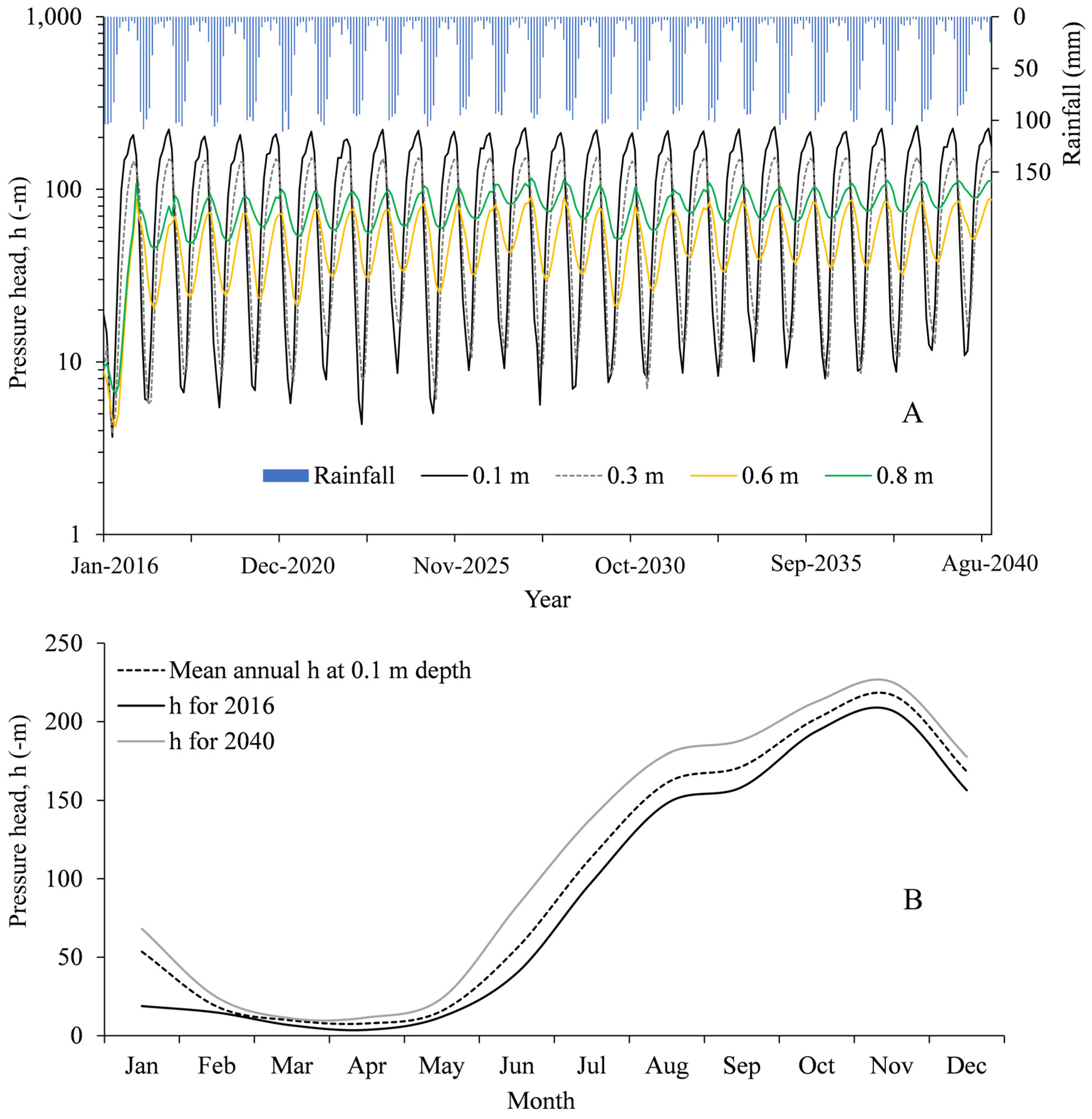
3.2. Hydrological Simulations
4. Discussion
4.1. General Hydrological Features
4.2. Final Remarks
5. Conclusions
- Based on 95% levels of confidence, in order to obtain a positive year-based water budget, annual rainfall should be higher than 443 mm.
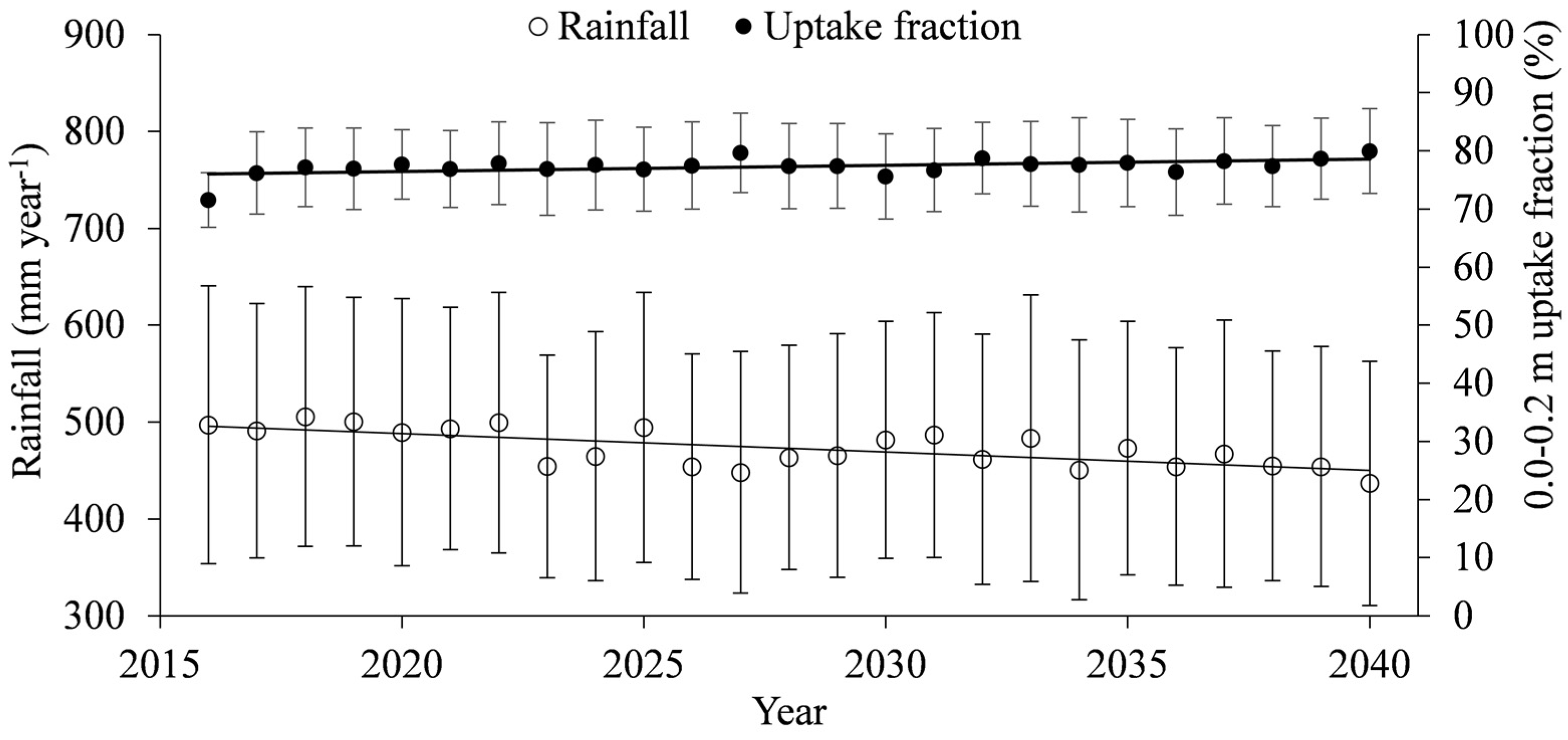
- Compared to simulations with the current climate, climate change would lead to an increase of actual transpiration of 36%, and a decrease of 16% and 34% for soil evaporation and interception losses, respectively, whereas the amount of precipitation returned to the atmosphere as evapotranspiration was predicted to reach, on average, 98%.
- The greatest change in water balance components under the simulated climate change was predicted for deep drainage, accounting only for 2% of the annual rainfall, followed by a soil-water reduction of 38%.
- Regarding soil water availability, the soil-plant-atmosphere fluxes seem to be controlled by the top layer (0.0–0.2 m), which provides, on average, 80% of the total transpiration, suggesting that the Caatinga biome may become completely soil-water pulse dominated under scenarios of reduced water availability.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Salazar, L.F.; Nobre, C.A.; Oyama, M.D. Climate change consequences on the biome distribution in tropical South America. Geophys. Res. Lett. 2007, 34, 1–6. [Google Scholar] [CrossRef]
- Jasechko, S.; Sharp, Z.D.; Gibson, J.J.; Birks, S.J.; Yi, Y.; Fawcett, P.J. Terrestrial water fluxes dominated by transpiration. Nature 2013, 496, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Sterling, S.M.; Ducharne, A.; Polcher, J. The impact of global land-cover change on the terrestrial water cycle. Nat. Clim. Chang. 2012, 3, 385–390. [Google Scholar] [CrossRef]
- Seneviratne, S.L.; Corti, T.; Davin, E.L.; Hirschi, M.; Jaeger, E.B.; Lehner, I.; Orlowsky, B.; Teuling, A.J. Investigating soil moisture-climate interactions in a changing climate: A review. Earth Sci. Rev. 2010, 99, 125–161. [Google Scholar] [CrossRef]
- Marengo, J.A.; Chou, S.C.; Kay, G.; Alves, L.M.; Pesquero, J.F.; Soares, W.R.; Santos, D.C.; Lyra, A.A.; Sueiro, G.; Betts, R.; et al. Development of regional future climate change scenarios in South America using the Eta CPTEC/HadCM3 climate change projections: Climatology and regional analyses for the Amazon, São Francisco and the Paraná River basins. Clim. Dyn. 2012, 38, 1829–1848. [Google Scholar] [CrossRef]
- Seneviratne, S.I.; Nicholls, N.; Easterling, D.; Goodess, C.M.; Kanae, S.; Kossin, J.; Luo, Y.; Marengo, J.; Mcinnes, K.; Rahimi, M.; et al. Changes in climate extremes and their impacts on the natural physical environment. In Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation: A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC); Field, C.B., Barros, V., Stocker, T.F., Qin, D., Dokken, D.J., Ebi, K.L., Mastrandrea, M.D., Mach, K.J., Planttner, G.K., Allen, S.K., et al., Eds.; Cambridge University Press: Cambridge, UK and New York, NY, USA, 2012; pp. 109–230. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, B.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Wang, L.; D’odorico, P.; Evans, J.P.; Eldridge, D.J.; McCabe, M.F.; Caylor, K.K.; King, E.G. Dryland ecohydrology and climate change: Critical issues and technical advances. Hydrol. Earth Syst. 2012, 16, 2585–2603. [Google Scholar] [CrossRef]
- Klein, T.; Yakir, D.; Buchmann, N.; Grünzweig, J.M. Towards an advanced assessment of the hydrological vulnerability of forests to climate change-induced drought. New Phytol. 2014, 3, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, E.A.R.; Metselaar, K.; De Jong van Lier, Q.; de Araújo, J.C. Importance of soil water to the Caatinga biome, Brazil. Ecohydrology 2016, 9, 1313–1327. [Google Scholar] [CrossRef]
- Pinheiro, E.A.R.; Costa, C.A.G.; de Araújo, J.C. Effective root depth of the Caatinga Biome. J. Arid Environ. 2013, 89, 1–4. [Google Scholar] [CrossRef]
- Marengo, J.A.; Torres, R.R.; Alves, L.M. Drought in Northeast Brazil—Past, present, and future. Theor. Appl. Climatol. 2016. [Google Scholar] [CrossRef]
- Leal, I.R.; Da Silva, J.M.C.; Tabarelli, M.; Lacher, T.E., Jr. Changing the course of biodiversity conservation in the Caatinga of Northeastern Brazil. Conserv. Biol. 2005, 19, 701–706. [Google Scholar] [CrossRef]
- Peter, A.J.; de Araújo, J.C.; Araújo, N.A.M.; Hermann, H.J. Flood avalanches in a semiarid basin with a dense reservoir network. J. Hydrol. 2014, 512, 408–420. [Google Scholar] [CrossRef]
- D’odorico, P.; Bhattachan, A.; Davis, K.F.; Ravi, S.; Runyan, C.W. Global desertification: Drivers and feedbacks. Adv. Water Resour. 2013, 51, 326–344. [Google Scholar] [CrossRef]
- De Figueiredo, J.V.; de Araújo, J.C.; Medeiros, P.H.A.; Costa, A.C. Runoff initiation in a preserved semiarid Caatinga small watershed, Northeastern Brazil. Hydrol. Process. 2016, 30, 2390–2400. [Google Scholar] [CrossRef]
- Medeiros, P.H.A.; de Araújo, J.C. Temporal variability of rainfall in a semiarid environment in Brazil and its effect on sediment transport processes. J. Soils Sediments 2014, 14, 1216–1223. [Google Scholar] [CrossRef]
- Costa, C.A.G.; Lopes, J.W.B.; Pinheiro, E.A.R.; de Araújo, J.C.; Gomes Filho, R.R. Spatial behaviour of soil moisture in the root zone of the Caatinga biome. Rev. Ciên. Agron. 2013, 44, 685–694. [Google Scholar] [CrossRef]
- Güntner, A.; Krol, M.; de Araújo, J.C.; Bronstert, A. Simple water balance modeling of surface reservoir systems in a large data-scarce semiarid region. Hydrol. Sci. J. 2004, 49, 901–918. [Google Scholar] [CrossRef]
- Stöckle, C.O.; Campbell, G.S.; Nelson, R. ClimGen Manual; Biological Systems Engineering Department, Washington State University: Pullman, WA, USA, 1999; p. 28. [Google Scholar]
- Kroes, J.G.; van Dam, J.C.; Groenendijk, P.; Hendriks, R.F.A.; Jacobs, C.M.J. SWAP Version 3.2. Theory Description and User Manual; Wageningen: Alterra, The Netherlands, 2008; p. 262. [Google Scholar]
- Mualem, Y. A New model for predicting the hydraulic conductivity of unsaturated porous media. Water Resour. Res. 1976, 12, 513–522. [Google Scholar] [CrossRef]
- Van Genuchten, M.T. A closed-form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- De Jong van Lier, Q.; van Dam, J.C.; Metselaar, K.; de Jong, R.; Duijnisveld, W.H.M. Macroscopic Root Water Uptake Distribution Using a Matric Flux Potential Approach. Vadose Zone J. 2008, 7, 1065–1078. [Google Scholar] [CrossRef]
- De Jong van Lier, Q.; van Dam, J.C.; Durigon, A.; dos Santos, M.A.; Metselaar, K. Modeling water potentials and flows in the soil-plant system comparing hydraulic resistances and transpiration reduction functions. Vadose Zone J. 2013, 12, 1–20. [Google Scholar] [CrossRef]
- Gash, J.H.C. An analytical model of rainfall interception by forests. Q. J. R. Meteorol. Soc. 1979, 105, 43–55. [Google Scholar] [CrossRef]
- Medeiros, P.H.A.; de Araújo, J.C.; Bronstert, A. Interception measurement and assessment of Gash model performance for a tropical semi-arid region. Rev. Ciênc. Agron. 2009, 40, 165–174. [Google Scholar]
- Gourdiaan, J.; van Laar, H.H. Modelling Potential Crop Growth Processes, 2nd ed.; Kluwer Academic Publishers: Wageningen, The Netherlands, 1994; p. 238. [Google Scholar]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Raz-Yaseef, N.; Yakir, D.; Rotenberg, E.; Schiller, G.; Cohen, S. Ecohydrology of a semi-arid forest: Partitioning among water balance and its components implications for predicted precipitation changes. Ecohydrology 2010, 3, 143–154. [Google Scholar]
- Ohmura, A.; Wild, M. Is the hydrological cycle accelerating? Science 2002, 298, 1345–1346. [Google Scholar] [CrossRef] [PubMed]
- Peterson, T.C.; Golubev, V.S.; Groisman, P.Y. Evaporation losing its strength. Nature 1995, 377, 687–688. [Google Scholar] [CrossRef]
- Wild, M.; Ohmura, A.; Cubasch, U. GCM simulated surface energy fluxes in climate change experiments. J. Clim. 1997, 10, 3093–3110. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Schlaepfer, D.R.; Bradford, J.B. Ecohydrology of Dry Regions: Storage versus Pulse Soil Water Dynamics. Ecosystems 2014, 17, 1469–1479. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Jasechko, S. Transpiration in the global water cycle. Agric. For. Meteorol. 2014, 189–190, 115–117. [Google Scholar] [CrossRef]
- Gedney, N.; Cox, P.M.; Betts, R.A.; Boucher, O.; Huntingford, C.; Stott, A. Detection of a direct carbon dioxide effect in continental river runoff records. Nature 2006, 439, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Jackson, R.B.; Mooney, H.A. Stomatal responses to increased CO2: Implications from the plant to the global-scale. Plant Cell Environ. 1995, 18, 1214–1225. [Google Scholar] [CrossRef]
- Nowak, R.S.; Zitzer, S.F.; Babcock, D.; Smith-Longozo, V.; Charlet, T.N.; Coleman, J.S.; Seemann, J.R.; Smith, S.D. Elevated atmospheric CO2 does not conserve soil water in the Mojave desert. Ecology 2004, 85, 93–99. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Z.; Duffy, R.; Chen, W.; An, S.; Liu, S.; Liu, F. Analyzing relationships among water uptake patterns, rootlet biomass distribution and soil water content profile in a subalpine shrubland using water isotopes. Eur. J. Soil Biol. 2011, 47, 380–386. [Google Scholar] [CrossRef]
- Raz-Yaseef, N.; Yakir, D.; Schiller, G.; Cohen, S. Dynamics of evapotranspiration partitioning in a semi-arid forest as affected by temporal rainfall patterns. Agric. For. Meteorol. 2012, 157, 77–85. [Google Scholar] [CrossRef]
- Gaines, K.P.; Stanley, J.W.; Meinzer, F.C.; McCulloh, K.A.; Woodruff, D.R.; Chen, W.; Adams, T.S.; Lin, H.; Eissenstat, D.M. Reliance on shallow soil water in a mixed-hardwood forest in central Pennsylvania. Tree Physiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Adiku, S.G.K.; Rose, C.W.; Braddock, R.D.; Ozier-Lafontaine, H. On the simulation of root water extraction: Examination of a minimum energy hypothesis. Soil Sci. 2000, 165, 226–236. [Google Scholar] [CrossRef]
- Jackson, R.B.; Sperry, J.S.; Dawson, T.E. Root water uptake and transport: Using physiological processes in global predictions. Trends Plant Sci. 2000, 5, 482–488. [Google Scholar] [CrossRef]
- Bucci, S.J.; Scholz, F.G.; Goldstein, G.; Meinzer, F.C.; Arce, M.E. Soil water availability and rooting depth as determinants of hydraulic architecture of Patagonian woody species. Oecologia 2009, 160, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.K.; Chase, M.W.; de Bruijn, A.; Fay, M.F.; Woodward, F.I. Temperature-based population segregation in birch. Ecol. Lett. 2003, 6, 1–3. [Google Scholar] [CrossRef]
- Jump, A.; Peñuelas, J. Running to stand still: Adaptation and response of plants to rapid climate change. Ecol. Lett. 2005, 8, 1010–1020. [Google Scholar] [CrossRef]
- Challinor, A.J.; Koehler, A.K.; Ramirez-Villegas, J.; Whitfield, S.; Das, B. Current warming will reduce yields unless maize breeding and seed systems adapt immediately. Nat. Clim. Chang. 2016. [Google Scholar] [CrossRef]
- Waring, R.H.; Coops, N.C. Predicting large wildfires across western North America by modeling seasonal variation in soil water balance. Clim. Chang. 2016, 135, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Krueger, E.S.; Ochsner, T.E.; Engle, D.M.; Carlson, J.D.; Twidwell, D.; Fuhlendorf, S.D. Soil moisture affects growing-season wildfire size in the Southern Great Plains. Soil Sci. Soc. Am. J. 2015, 79, 1567–1576. [Google Scholar] [CrossRef]
- Running, S.W. Is global warming causing more, larger wildfires? Science 2006, 313, 927–928. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Parameterization | ||||
|---|---|---|---|---|
| Parameter | Value | Source | ||
| hroot (m) | −160 | [10] | ||
| Rood radius (m) | 4.3 × 10−4 | [10] | ||
| Canopy albedo (–) | 0.16 | [10] | ||
| Soil albedo (–) | 0.24 | [10] | ||
| LAI range (–) | 0.04–1.3 | According to Equations (3) and (4) | ||
| k (–) | 0.75 | [25] | ||
| Kc range (–) | 0.98–1.1 | According to Equation (2) | ||
| Ref (m) | 0.78 | [11] | ||
| Soil-depth dependent parameterization | ||||
| Parameter | Soil-layer thickness (m) | |||
| 0.0–0.2 | 0.2–0.4 | 0.4–0.6 | 0.6–0.8 | |
| θr (m3·m−3) | 0.000 | 0.000 | 0.000 | 0.000 |
| θs (m3·m−3) | 0.418 | 0.440 | 0.433 | 0.415 |
| α·(m−1) | 6.8 | 22.5 | 24.9 | 13.9 |
| n (–) | 1.21 | 1.86 | 1.162 | 1.170 |
| Ks (m·day−1) | 2.22 | 2.5 | 0.36 | 0.16 |
| RLD (m·m−3) | 3700 | 2800 | 2000 | 1600 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, E.A.R.; Van Lier, Q.D.J.; Bezerra, A.H.F. Hydrology of a Water‐Limited Forest under Climate Change Scenarios: The Case of the Caatinga Biome, Brazil. Forests 2017, 8, 62. https://doi.org/10.3390/f8030062
Pinheiro EAR, Van Lier QDJ, Bezerra AHF. Hydrology of a Water‐Limited Forest under Climate Change Scenarios: The Case of the Caatinga Biome, Brazil. Forests. 2017; 8(3):62. https://doi.org/10.3390/f8030062
Chicago/Turabian StylePinheiro, Everton Alves Rodrigues, Quirijn De Jong Van Lier, and Andre Herman Freire Bezerra. 2017. "Hydrology of a Water‐Limited Forest under Climate Change Scenarios: The Case of the Caatinga Biome, Brazil" Forests 8, no. 3: 62. https://doi.org/10.3390/f8030062
APA StylePinheiro, E. A. R., Van Lier, Q. D. J., & Bezerra, A. H. F. (2017). Hydrology of a Water‐Limited Forest under Climate Change Scenarios: The Case of the Caatinga Biome, Brazil. Forests, 8(3), 62. https://doi.org/10.3390/f8030062