Assessing Pine Processionary Moth Defoliation Using Unmanned Aerial Systems


Abstract
1. Introduction
2. Materials and Methods
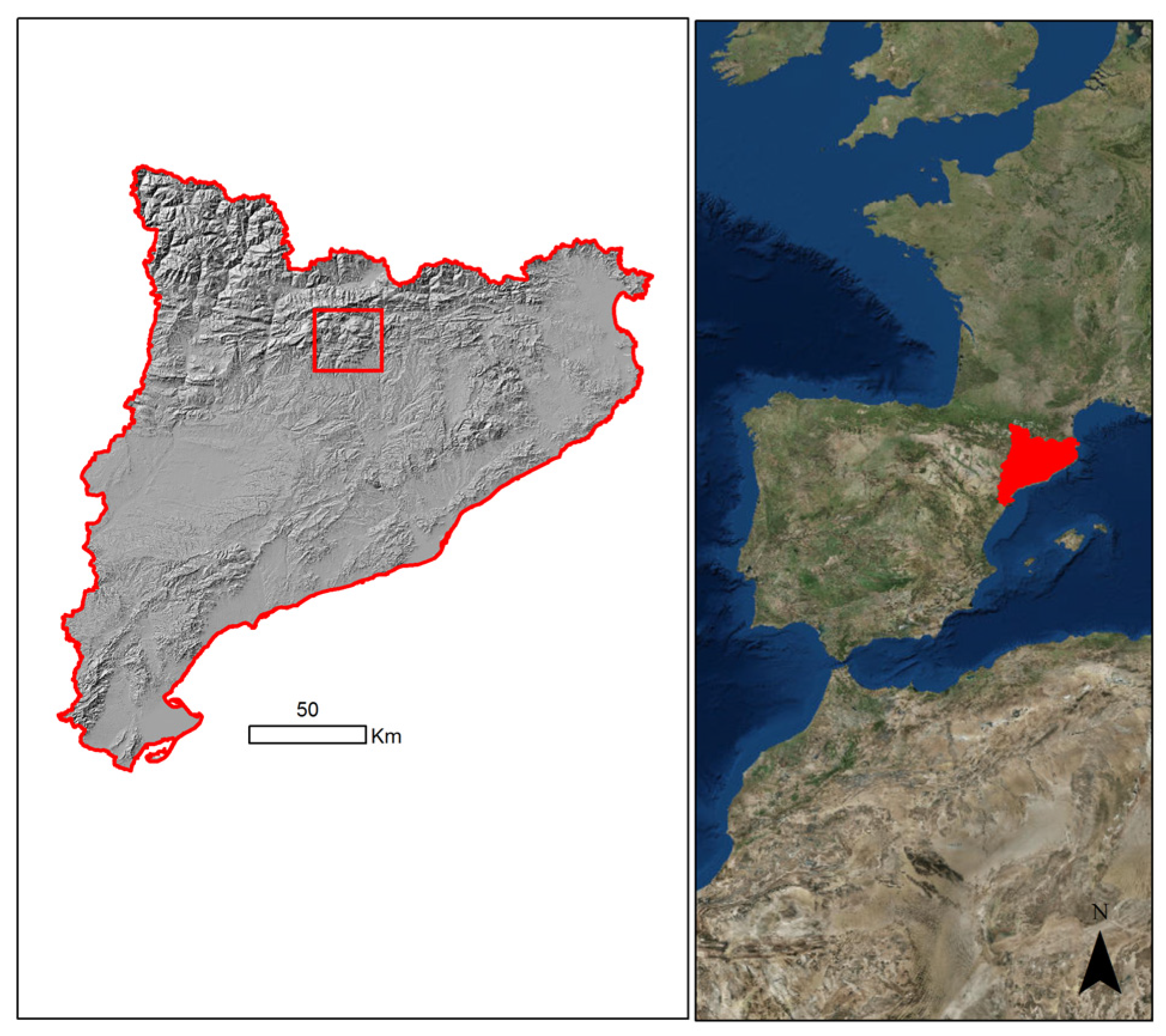
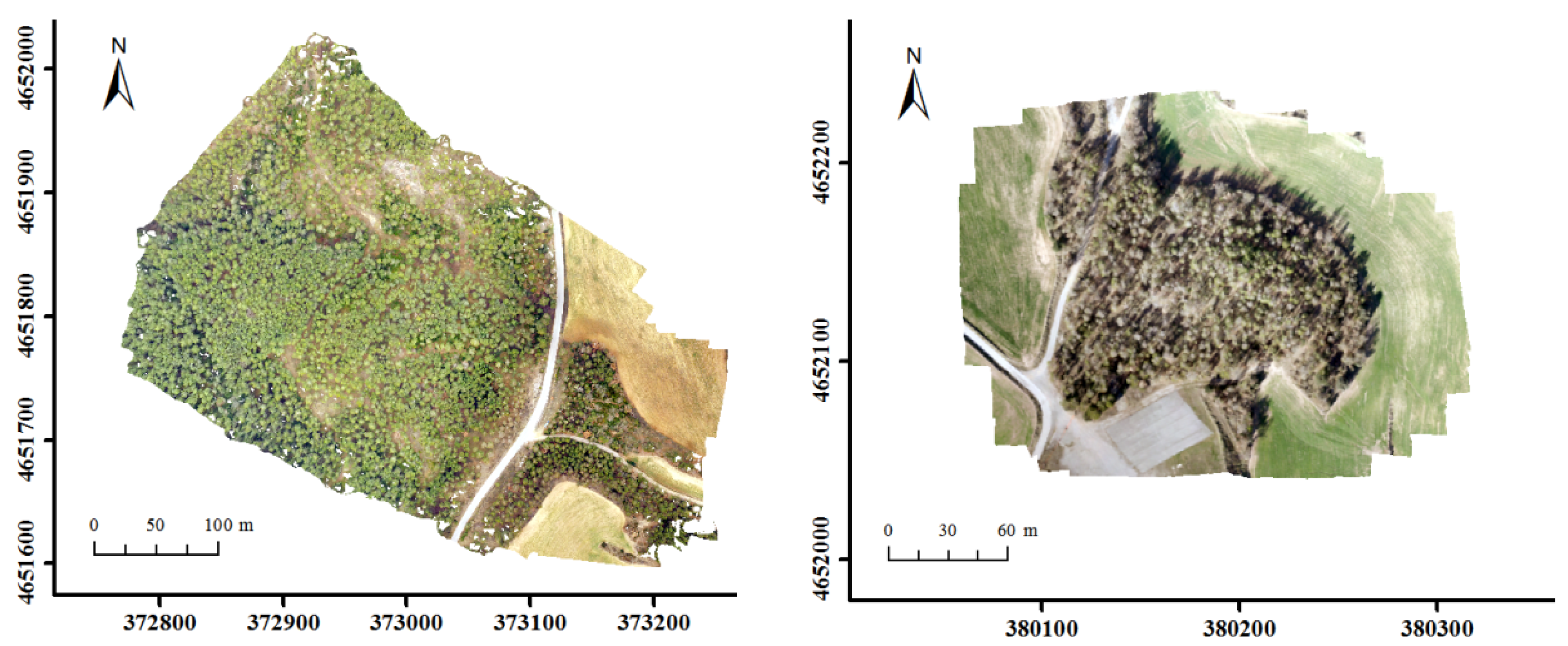
2.1. Study Area
2.2. PPM Life Cycle
2.3. UAS-Based Image Acquisition and Data Preprocessing
2.4. Field Validation Data
2.5. Data Analysis
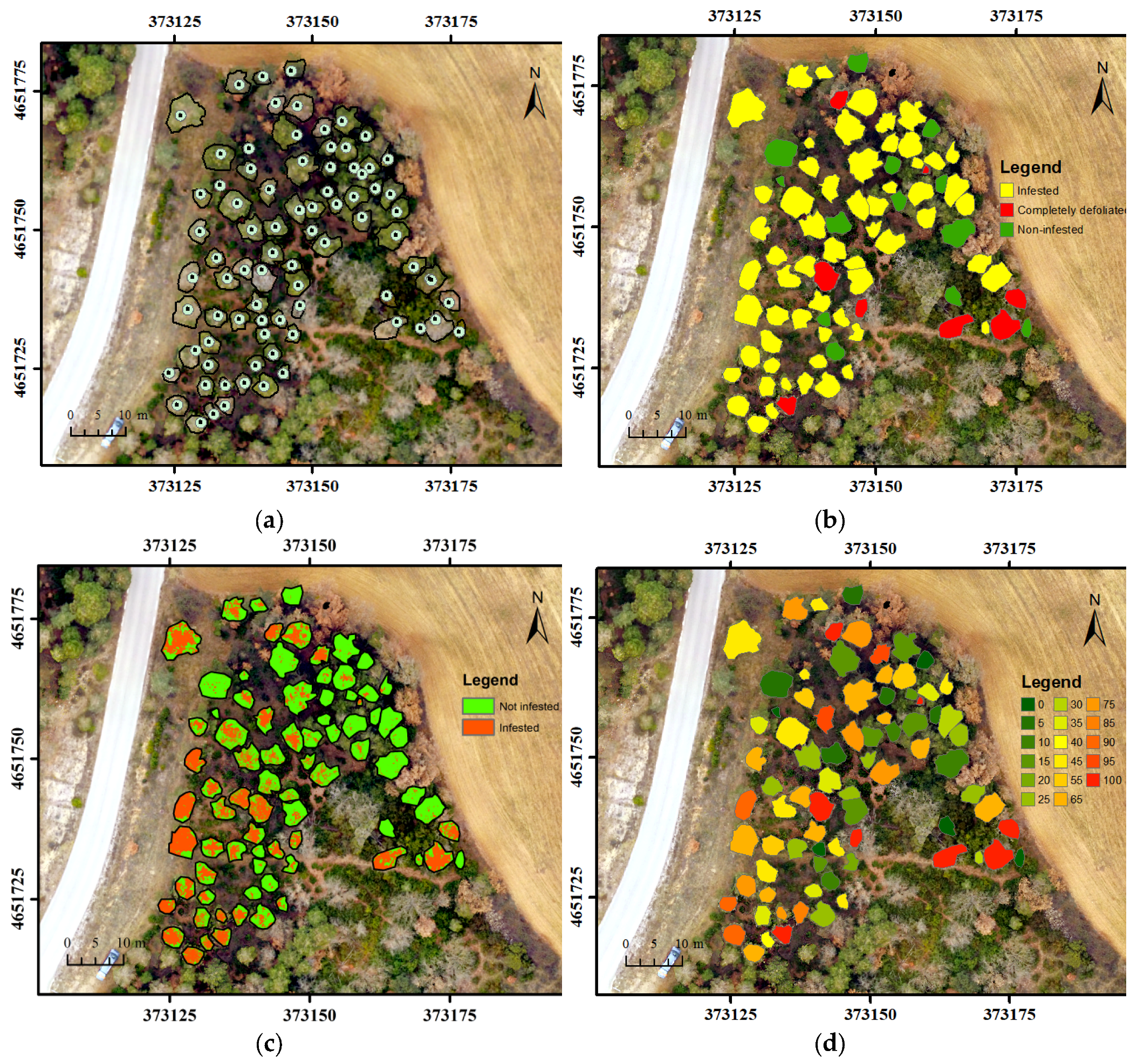
2.5.1. Image-Based Defoliation Assessment
2.5.2. Tree-Level Defoliation
2.5.3. Accuracy of Infested Tree Identification and Validation of Percent Defoliation
3. Results
3.1. Field Validation on PPM Defoliation Using RGB-UAS Images
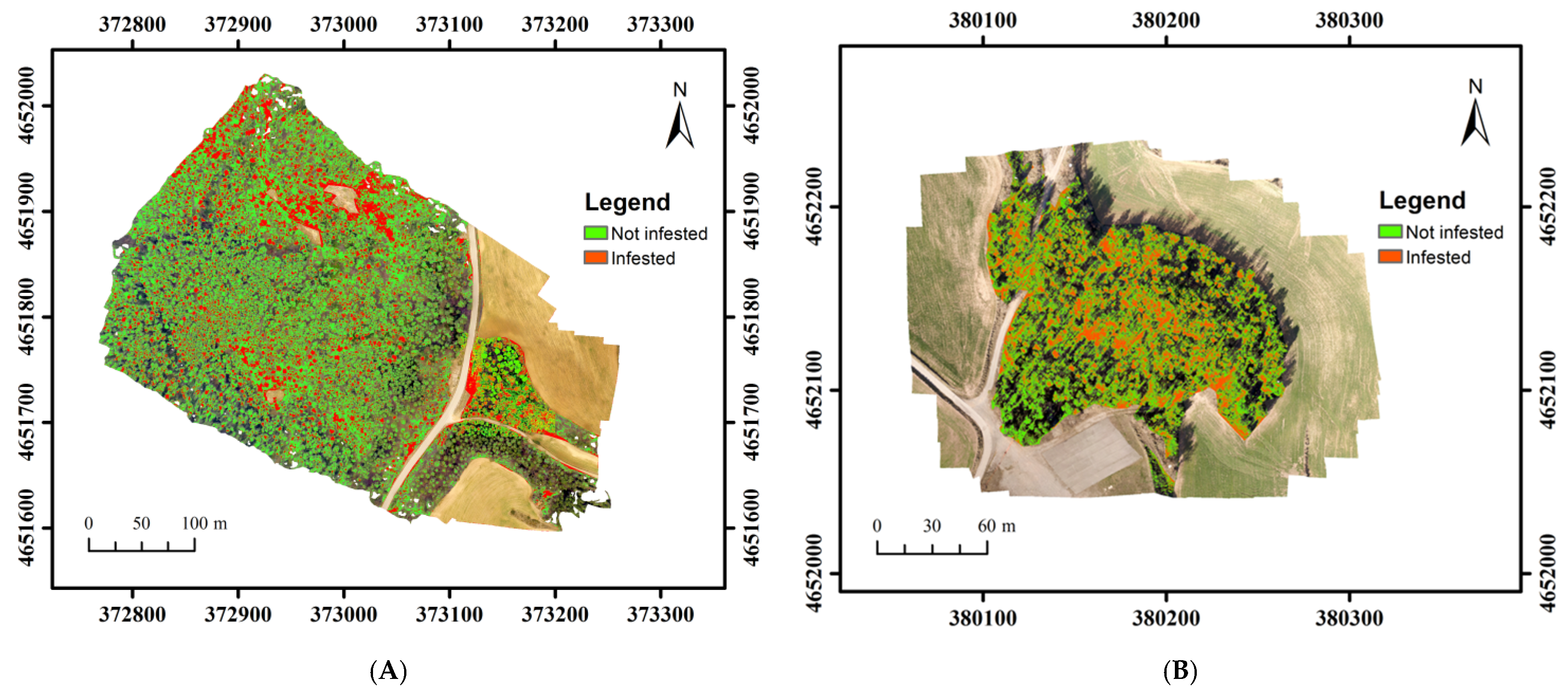
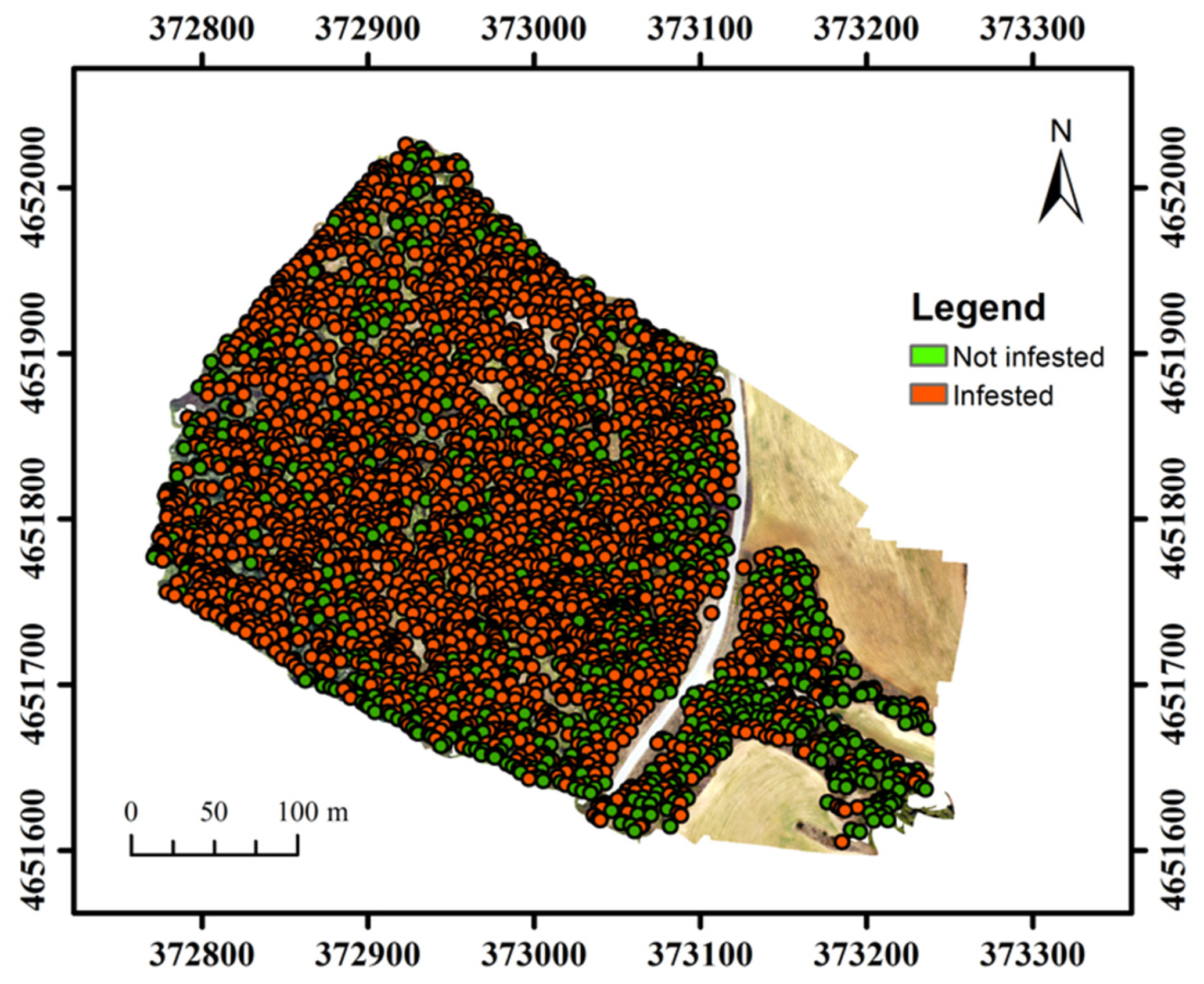
3.2. PPM Defoliation in Torregassa and Bosquet
3.3. PPM Defoliation Patterns at Tree-Level Scale and Nests
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cayuela, L.; Hódar, J.A.; Zamora, R. Is insecticide spraying a viable and cost-efficient management practice to control pine processionary moth in Mediterranean woodlands? For. Ecol. Manag. 2011, 261, 1732–1737. [Google Scholar] [CrossRef]
- Berryman, A. Population Cycles: The Case for Trophic Interactions; Oxford University Press: New York, NY, USA, 2002. [Google Scholar]
- Hódar, J.A.; Zamora, R.; Castro, J.; Baraza, E. Feast and famine: Previous defoliation limiting survival of pine processionary caterpillar Thaumetopoea pityocampa in Scots pine Pinus sylvestris. Acta Oecol. 2004, 26, 203–210. [Google Scholar] [CrossRef]
- Hódar, J.A.; Castro, J.; Zamora, R. Pine processionary caterpillar Thaumetopoea pityocampa as a new threat for relict Mediterranean Scots pine forests under climatic warming. Biol. Conserv. 2003, 110, 123–129. [Google Scholar] [CrossRef]
- Moore, B.; Allard, G. Climate Change Impacts on Forest Health; Working paper FBS/34E; FAO: Rome, Italy, 2008. [Google Scholar]
- Battisti, A.; Holm, G.; Fagrell, B.; Larsson, S. Urticating Hairs in Arthropods: Their Nature and Medical Significance. Annu. Rev. Entomol. 2011, 56, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Cayuela, L.; Hernández, R.; Hódar, J.A.; Sánchez, G.; Zamora, R. Tree damage and population density relationships for the pine processionary moth: Prospects for ecological research and pest management. For. Ecol. Manag. 2014, 328, 319–325. [Google Scholar] [CrossRef]
- Battisti, A. Forests and climate change-Lessons from insects. IForest 2008, 1, 1–5. [Google Scholar] [CrossRef]
- Jactel, H.; Menassieu, P.; Vétillard, F.; Barthélémy, B.; Piou, D.; Frérot, B.; Rousselet, J.; Goussard, F.; Branco, M.; Battisti, A. Population monitoring of the pine processionary moth (Lepidoptera: Thaumetopoeidae) with pheromone-baited traps. For. Ecol. Manag. 2006, 235, 96–106. [Google Scholar] [CrossRef]
- Rullan-Silva, C.D.; Olthoff, A.E.; Delgado de la Mata, J.A.; Pajares-Alonso, J.A. Remote Monitoring of Forest Insect Defoliation—A Review. For. Syst. 2013, 22, 377–391. [Google Scholar] [CrossRef]
- Silva, C.A.; Hudak, A.T.; Klauberg, C.; Vierling, L.A.; González-Benecke, C.; de Padua Chaves Carvalho, S.; Rodríguez, L.C.E.; Cardil, A. Combined effect of pulse density and grid cell size on predicting and mapping aboveground carbon in fast-growing Eucalyptus forest plantation using airborne LiDAR data. Carbon Balance Manag. 2017, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.A.; Klauberg, C.; Hudak, A.T.; Vierling, L.A. Predicting Stem Total and Assortment Volumes in an Industrial Pinus taeda L. Forest Plantation Using Airborne Laser Scanning Data and Random Forest. Forests 2017, 8, 254. [Google Scholar] [CrossRef]
- Kantola, T.; Vastaranta, M.; Lyytikäinen-Saarenmaa, P.; Holopainen, M.; Kankare, V.; Talvitie, M.; Hyyppä, J. Classification of needle loss of individual scots pine trees by means of airborne laser scanning. Forests 2013, 4, 386–403. [Google Scholar] [CrossRef]
- Ortiz, S.M.; Breidenbach, J.; Kändler, G. Early detection of bark beetle green attack using terraSAR-X and rapideye data. Remote Sens. 2013, 5, 1912–1931. [Google Scholar] [CrossRef]
- Wulder, M.A.; Dymond, C.C.; White, J.C.; Leckie, D.G.; Carroll, A.L. Surveying mountain pine beetle damage of forests: A review of remote sensing opportunities. For. Ecol. Manag. 2006, 221, 27–41. [Google Scholar] [CrossRef]
- Näsi, R.; Honkavaara, E.; Lyytikäinen-Saarenmaa, P.; Blomqvist, M.; Litkey, P.; Hakala, T.; Viljanen, N.; Kantola, T.; Tanhuanpää, T.; Holopainen, M. Using UAV-based photogrammetry and hyperspectral imaging for mapping bark beetle damage at tree-level. Remote Sens. 2015, 7, 15467–15493. [Google Scholar] [CrossRef]
- Lehmann, J.R.K.; Nieberding, F.; Prinz, T.; Knoth, C. Analysis of unmanned aerial system-based CIR images in forestry-a new perspective to monitor pest infestation levels. Forests 2015, 6, 594–612. [Google Scholar] [CrossRef]
- Puliti, S.; Ene, L.T.; Gobakken, T.; Næsset, E. Use of partial-coverage UAV data in sampling for large scale forest inventories. Remote Sens. Environ. 2017, 194, 115–126. [Google Scholar] [CrossRef]
- Puliti, S.; Olerka, H.; Gobakken, T.; Næsset, E. Inventory of Small Forest Areas Using an Unmanned Aerial System. Remote Sens. 2015, 7, 9632–9654. [Google Scholar] [CrossRef]
- Whitehead, K.; Hugenholtz, C.H. Remote sensing of the environment with small unmanned aircraft systems (UASs), part 1: A review of progress and challenges. J. Unmanned Veh. Syst. 2014, 2, 69–85. [Google Scholar] [CrossRef]
- Gao, M.; Xu, X.; Klinger, Y.; van der Woerd, J.; Tapponnier, P. High-resolution mapping based on an Unmanned Aerial Vehicle (UAV) to capture paleoseismic offsets along the Altyn-Tagh fault, China. Sci. Rep. 2017, 7, 8281. [Google Scholar] [CrossRef] [PubMed]
- Casbeer, D.W.; Kingston, D.B.; Beard, R.W.; McLain, T.W. Cooperative forest fire surveillance using a team of small unmanned air vehicles. Int. J. Syst. Sci. 2006, 37, 351–360. [Google Scholar] [CrossRef]
- Hung, C.; Bryson, M.; Sukkarieh, S. Multi-class predictive template for tree crown detection. ISPRS J. Photogramm. Remote Sens. 2012, 68, 170–183. [Google Scholar] [CrossRef]
- Dandois, J.P.; Ellis, E.C. High spatial resolution three-dimensional mapping of vegetation spectral dynamics using computer vision. Remote Sens. Environ. 2013, 136, 259–276. [Google Scholar] [CrossRef]
- Feng, Q.; Liu, J.; Gong, J. UAV Remote Sensing for Urban Vegetation Mapping Using Random Forest and Texture Analysis. Remote Sens. 2015, 7, 1074–1094. [Google Scholar] [CrossRef]
- Hung, C.; Xu, Z.; Sukkarieh, S. Feature learning based approach for weed classification using high resolution aerial images from a digital camera mounted on a UAV. Remote Sens. 2014, 6, 12037–12054. [Google Scholar] [CrossRef]
- Torresan, C.; Berton, A.; Carotenuto, F.; Di Gennaro, S.F.; Gioli, B.; Matese, A. Forestry applications of UAVs in Europe: A review. Int. J. Remote Sens. 2017, 38, 2427–2447. [Google Scholar]
- Regos, A.; Aquilué, N.; Retana, J.; De Cáceres, M.; Brotons, L. Using Unplanned Fires to Help Suppressing Future Large Fires in Mediterranean Forests. PLoS ONE 2014, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chenchouni, H.; Zanati, K.; Rezougui, A.; Briki, A.; Arar, A. Population Monitoring of Pine Processionary Moth (Thaumetopoea pityocampa) by Pheromone Trapping at the Southern Limit of Distribution of Pinus halepensis in. For. Sci. Technol. 2010, 6, 67–79. [Google Scholar]
- Tamburini, G.; Marini, L.; Hellrigl, K.; Salvadori, C.; Battisti, A. Effects of climate and density-dependent factors on population dynamics of the pine processionary moth in the Southern Alps. Clim. Chang. 2013, 121, 701–712. [Google Scholar] [CrossRef]
- Verhoeven, G. Taking computer vision aloft—archaeological three-dimensional reconstructions from aerial photographs with photoscan. Archaeol. Prospect. 2011, 18, 67–73. [Google Scholar] [CrossRef]
- Lucieera, A.; Turner, D.; Kingb, D.H.; Robinson, S.A. Using an Unmanned Aerial Vehicle ( UAV ) to capture micro-topography of Antarctic moss beds. Int. J. Appl. Earth Obs. Geoinf. 2014, 27, 52–62. [Google Scholar] [CrossRef]
- Mohan, M.; Silva, C.A.; Klauberg, C.; Jat, P.; Catts, G.; Cardil, A.; Hudak, A.T.; Dia, M. Individual Tree Detection from Unmanned Aerial Vehicle (UAV) Derived Canopy Height Model in an Open Canopy Mixed Conifer Forest. Forests 2017, 8, 340. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Cardil, A.; Molina, D.M.; Kobziar, L.N. Extreme temperature days and their potential impacts on southern Europe. Nat. Hazards Earth Syst. Sci. 2014, 14, 3005–3014. [Google Scholar] [CrossRef]
- Cardil, A.; Eastaugh, C.S.; Molina, D.M. Extreme temperature conditions and wildland fires in Spain. Theor. Appl. Climatol. 2015, 122, 219–228. [Google Scholar] [CrossRef]
- Molina-Terrén, D.M.; Cardil, A.; Kobziar, L.N. Practitioner perceptions of wildland fire management across South Europe and Latin America. Forests 2016, 7, 184. [Google Scholar] [CrossRef]
- Molina-Terrén, D.M.; Cardil, A. Temperature determining larger wildland fires in NE Spain. Theor. Appl. Climatol. 2016, 125, 295–302. [Google Scholar] [CrossRef]
- Hall, R.J.; Castilla, G.; White, J.C.; Cooke, B.J.; Skakun, R.S. Remote sensing of forest pest damage: A review and lessons learned from a Canadian perspective. Can. Entomol. 2016, 148, S296–S356. [Google Scholar] [CrossRef]
- Eklundh, L.; Johansson, T.; Solberg, S. Mapping insect defoliation in Scots pine with MODIS time-series data. Remote Sens. Environ. 2009, 113, 1566–1573. [Google Scholar] [CrossRef]
- Solberg, S. Mapping gap fraction, LAI and defoliation using various ALS penetration variables. Int. J. Remote Sens. 2010, 31, 1227–1244. [Google Scholar] [CrossRef]
- Townsend, P.A.; Singh, A.; Foster, J.R.; Rehberg, N.J.; Kingdon, C.C.; Eshleman, K.N.; Seagle, S.W. A general Landsat model to predict canopy defoliation in broadleaf deciduous forests. Remote Sens. Environ. 2012, 119, 255–265. [Google Scholar] [CrossRef]
- Dennison, P.E.; Nagler, P.L.; Hultine, K.R.; Glenn, E.P.; Ehleringer, J.R. Remote monitoring of tamarisk defoliation and evapotranspiration following saltcedar leaf beetle attack. Remote Sens. Environ. 2009, 113, 1462–1472. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field/RGB-UAS | Non-Infested | Infested | Total | Omission Error (%) |
|---|---|---|---|---|
| Non-infested | 12 | 1 | 13 | 7.7 |
| Infested | 16 | 52 | 68 | 23.5 |
| Total | 28 | 53 | 81 | |
| Commission error | 57.1 | 1.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardil, A.; Vepakomma, U.; Brotons, L. Assessing Pine Processionary Moth Defoliation Using Unmanned Aerial Systems. Forests 2017, 8, 402. https://doi.org/10.3390/f8100402
Cardil A, Vepakomma U, Brotons L. Assessing Pine Processionary Moth Defoliation Using Unmanned Aerial Systems. Forests. 2017; 8(10):402. https://doi.org/10.3390/f8100402
Chicago/Turabian StyleCardil, Adrián, Udayalakshmi Vepakomma, and Lluis Brotons. 2017. "Assessing Pine Processionary Moth Defoliation Using Unmanned Aerial Systems" Forests 8, no. 10: 402. https://doi.org/10.3390/f8100402
APA StyleCardil, A., Vepakomma, U., & Brotons, L. (2017). Assessing Pine Processionary Moth Defoliation Using Unmanned Aerial Systems. Forests, 8(10), 402. https://doi.org/10.3390/f8100402