
Effects of Repeated Growing Season Prescribed Fire on the Structure and Composition of Pine–Hardwood Forests in the Southeastern Piedmont, USA

Abstract
:
1. Introduction
- (1)
- How do the density and composition of saplings in the midstory change following repeated growing season burns?
- (2)
- How do the structure (i.e., basal area, density, and quadratic mean diameter) and composition of overstory trees change following repeated growing season burns?
- (3)
- How do the effects of repeated growing season burns on overstory and midstory structure and composition differ when fires are applied at different times within the growing season?
2. Materials and Methods
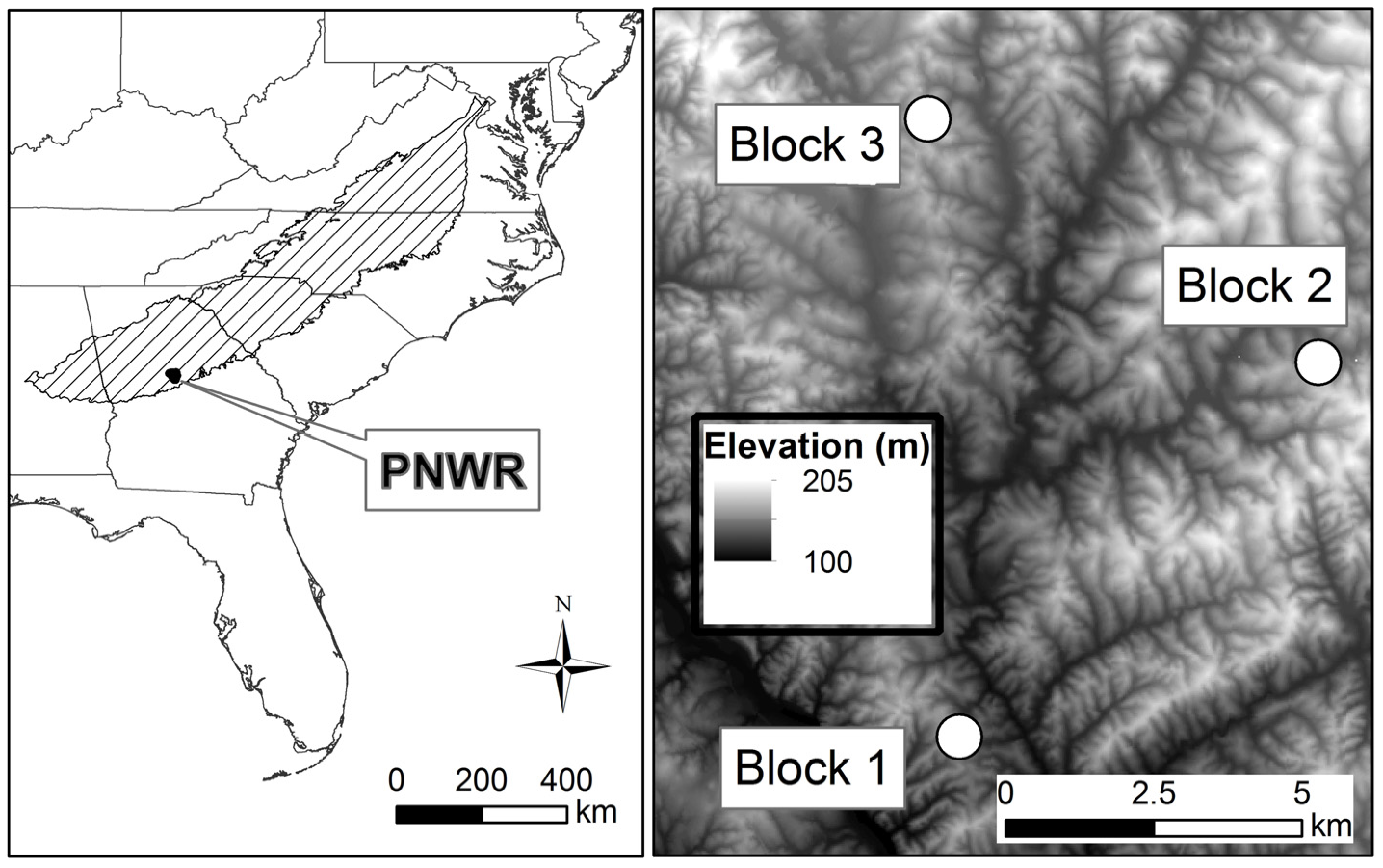
2.1. Study Site
2.2. Study Design
2.3. Field Sampling
2.4. Data Analysis
3. Results
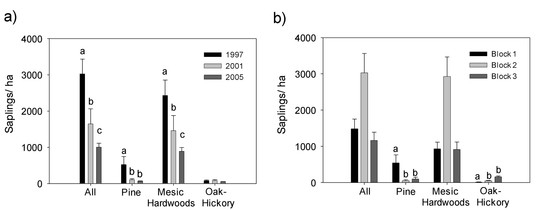
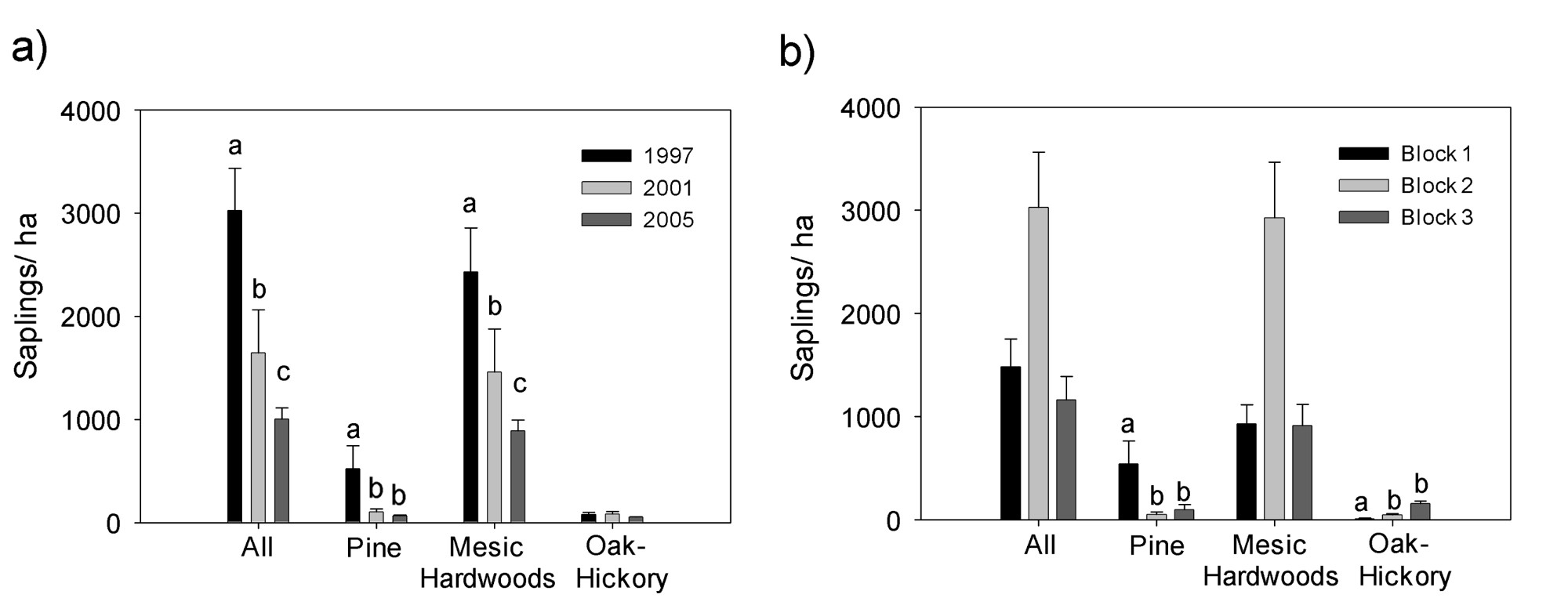
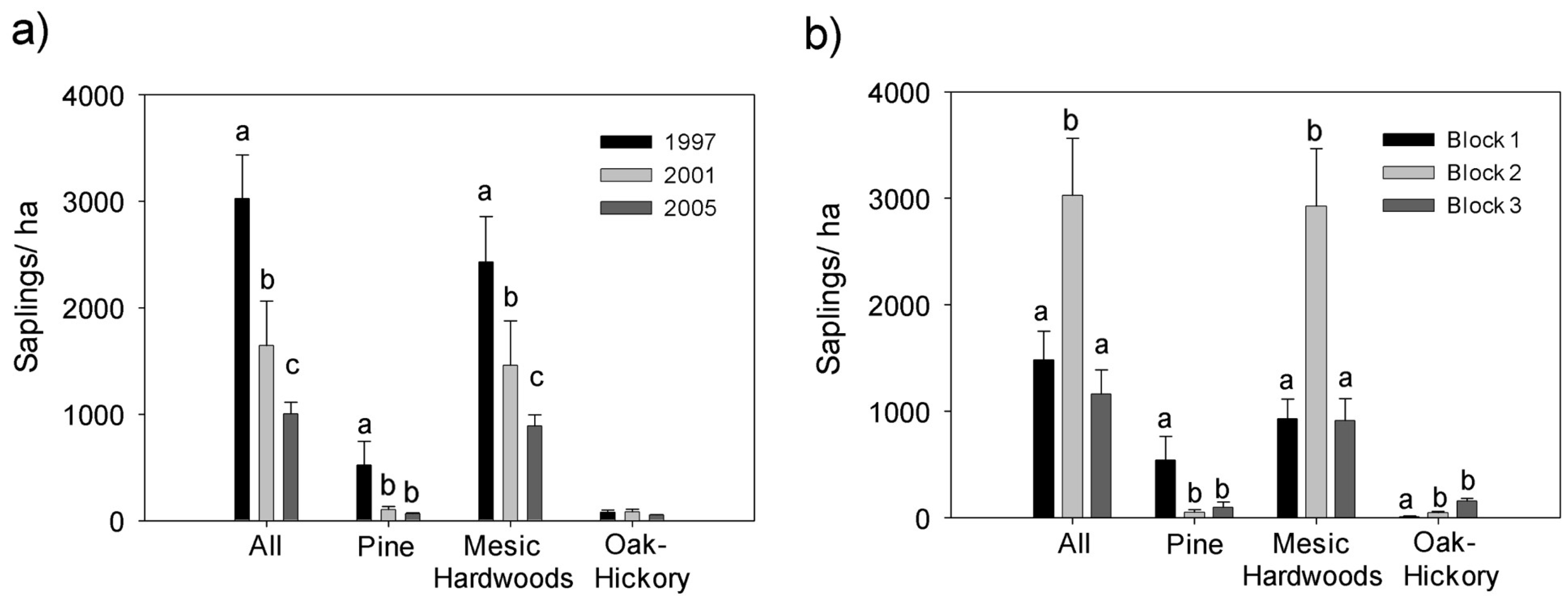
3.1. Saplings
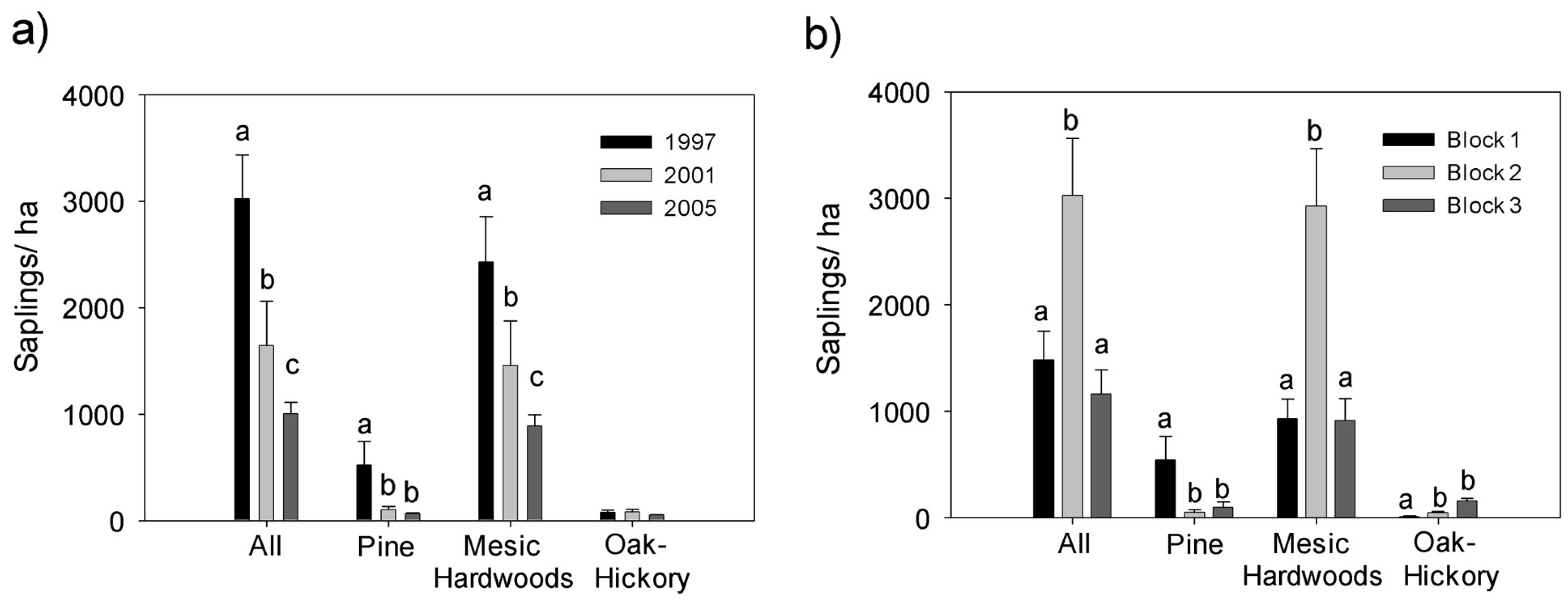
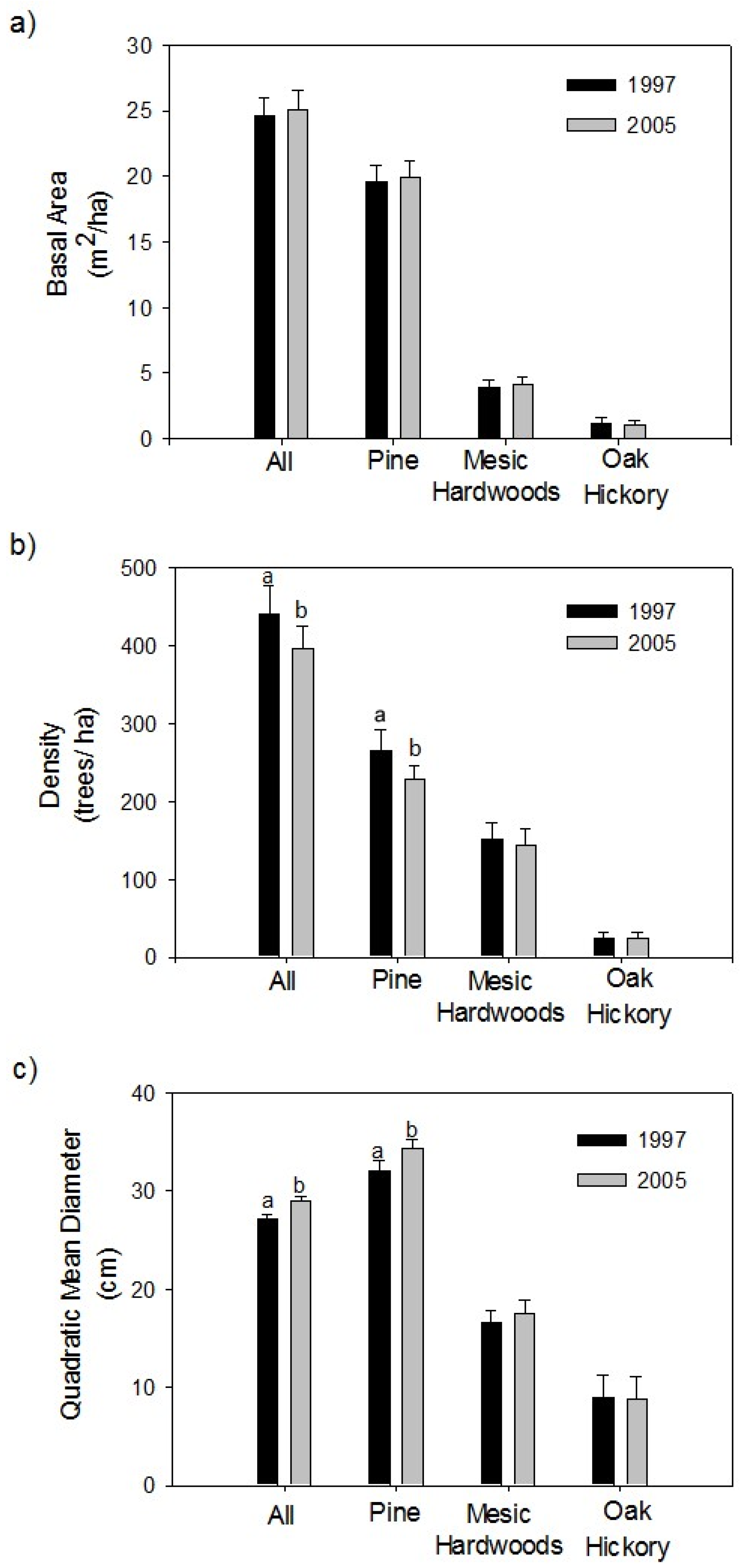
3.2. Overstory Trees
4. Discussion
4.1. Effects on Midstory Saplings
4.2. Effects on Overstory Trees
4.3. Timing of Growing Season Burn
4.4. Management Considerations
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moore, M.M.; Wallace Covington, W.; Fule, P.Z. Reference conditions and ecological restoration: A southwestern ponderosa pine perspective. Ecol. Appl. 1999, 9, 1266–1277. [Google Scholar] [CrossRef]
- Peterson, D.W.; Reich, P.B. Prescribed fire in an oak savanna: Fire frequency effects on stand structure and dynamics. Ecol. Appl. 2001, 11, 914–927. [Google Scholar] [CrossRef]
- Sparks, J.C.; Masters, R.E.; Engle, D.M.; Bukenhofer, G.A. Season of burn influences fire behavior and fuel consumption in restored shortleaf pine-grassland communities. Rest. Ecol. 2002, 10, 714–722. [Google Scholar] [CrossRef]
- Wade, D.D.; Brock, B.L.; Brose, P.H.; Grace, J.B.; Hoch, G.A.; Patterson, W.A. Fire in eastern ecosystems. In Wildland Fire in Ecosystems: Effects of Fire on Flora; GTR RMRS-42; Brown, J.K., Smith, J.K., Eds.; USDA forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000. [Google Scholar]
- McCaw, W.L. Managing forest fuels using prescribed fire-A perspective from southern Australia. For. Ecol. Manag. 2013, 294, 217–224. [Google Scholar] [CrossRef]
- Govender, N.; Trollope, W.S.W.; Van Wilgen, B.W. The effect of fire season, fire frequency, rainfall and management on fire intensity in savannah vegetation in South Africa. J. Appl. Ecol. 2006, 43, 748–758. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Botehlo, H.S. A review of prescribed burning effectiveness in fire hazard reduction. Int. J. Wildland Fire 2003, 12, 117–128. [Google Scholar] [CrossRef]
- Brockway, D.G.; Lewis, C.E. Long-term effects of dormant-season prescribed fire on plant community diversity, structure and productivity in a longleaf pine wiregrass ecosystem. For. Ecol. Manag. 1997, 96, 167–183. [Google Scholar] [CrossRef]
- Elliott, K.J.; Hendrick, R.L.; Major, A.E.; Vose, J.M.; Swank, W.T. Vegetation dynamics after a prescribed fire in the southern Appalachians. For. Ecol. Manag. 1999, 114, 199–213. [Google Scholar] [CrossRef]
- Ryan, K.C.; Knapp, E.E.; Varner, J.M. Prescribed fire in North American forests and woodlands: History, current practice, and challenges. Front. Ecol. Environ. 2013, 11, e15–e24. [Google Scholar] [CrossRef]
- Knapp, E.E.; Estes, B.L.; Skinner, C.N. Ecological Effects of Prescribed Fire Season; a Literature Review and Synthesis for Managers; USDA Forest Service General Technical Report PSW-GTR-224; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2009.
- Streng, D.R.; Glitzenstein, J.S.; Platt, W.J. Evaluating effects of season of burn in longleaf pine forests: A acritical literature and some results from an ongoing long-term study. In Proceedings of the Tall Timbers Fire Ecol. Conf. No. 18, Tallahassee, FL, USA, 30 May–2 June 1991; Tall Timbers Research Station: Tallahassee, FL, USA, 1993. [Google Scholar]
- Glitzenstein, J.S.; Streng, D.R.; Platt, W.J. Evaluating Effects of Season of Burn on Vegetation in Longleaf Pine Savannahs; Final Report; Florida Game and Freshwater Fish Committee: Tallahassee, FL, USA, 1990. [Google Scholar]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of fire regime and habitat on tree dynamics in north Florida longleaf pine savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar] [CrossRef]
- De Groot, W.J.; Wein, R.W. Effects of fire severity and season of burn on Betula glandulosa growth dynamics. Int. J. Wildland Fire 2004, 13, 287–295. [Google Scholar] [CrossRef]
- Kaufman, J.B.; Marin, R.E. Sprouting shrub response to different season and fuel consumption levels of prescribed fire in Sierra Nevada mixed conifer ecosystems. For. Sci. 1990, 36, 748–764. [Google Scholar]
- Williams, R.J.; Cook, G.D.; Gill, A.M.; Moore, P.H.R. Fire regime, fire intensity and tree survival in a tropical savanna in northern Australia. Aust. J. Ecol. 1999, 24, 50–59. [Google Scholar]
- Robbins, L.E.; Myers, R.L. Seasonal Effects of Prescribed Burning in Florida; Miscellaneous publication/Tall Timbers Research, Inc.: Tallahassee, FL, USA, 1992; Volume 8, pp. 1–97. [Google Scholar]
- Stoddard, H.L. Use of controlled fire in southeastern upland game management. J. For. 1935, 33, 346–351. [Google Scholar]
- Waldrop, T.A.; Van Lear, D.H.; Lloyd, F.T.; Harms, W.R. Long-Term Studies of Prescribed Burning in Loblolly Pine Forests of the Southeastern Coastal Plain; USDA Forest Service General Technical Report SE-45; USDA Forest Service, Southeastern Forest Experiment Station: Asheville, NC, USA, 1987.
- Reilly, M.J.; Waldrop, T.A.; O’Brien, J.J. Fuels management in the southern appalachian mountains, hot continental division. In Cumulative Watershed Effects of Fuel Management in the Eastern United States; USDA Forest Service General USDA Forest Service General Technical Report SRS-161; Southern Research Station: Asheville, NC, USA, 2012. [Google Scholar]
- Conner, R.N.; Snow, A.E.; O’Halloran, K.A. Red-cockaded woodpecker use of seedtree/shelterwood cuts in eastern Texas. Wildlife Soc. Bull. 1991, 19, 67–73. [Google Scholar]
- Conner, R.N.; O’Halloran, K.A. Cavity-tree selection by red-cockaded woodpeckers as related to growth dynamics of southern pines. Wilson. Bull. 1987, 99, 398–412. [Google Scholar]
- Conner, R.N.; Rudolph, D.C. Red-Cockaded Woodpecker Colony Status and Trends on the Angelina, Davy Crockett, and Sabine National Forests; USDA Forest Service Research Paper SO-250; Southern Forest Experimental Station: New Orleans, LA, USA, 1989.
- Loeb, S.C.; Pepper, W.D.; Doyle, A.T. Habitat characteristics of active and abandoned Red-cockaded woodpecker colonies. South J. Appl. For. 1992, 16, 120–125. [Google Scholar]
- Chaiken, L.E. The Behavior and Control of Understory Hardwoods in Loblolly Pine Stands; USDA Forest Service Research Tech. Note 72; Southern Forest Experimental Station: New Orleans, LA, USA, 1949.
- Harrington, T.A.; Stevenson, G.K. Repeat burns reduce small stems in Texas Big Thicket. J. For. 1955, 53, 847. [Google Scholar]
- Hodgkins, E.J. Effects of fire on undergrowth vegetation in upland southern pine forests. Ecology 1958, 39, 36–46. [Google Scholar] [CrossRef]
- Brender, E.V.; Cooper, R.W. Prescribed burning in Georgia’s Piedmont loblolly pine stands. J. For. 1968, 66, 31–36. [Google Scholar]
- Oosting, H.J. The comparative effect of surface and crown fire on the composition of a loblolly pine community. Ecology 1944, 25, 61–69. [Google Scholar] [CrossRef]
- Oosting, H.J.; Livingston, R.B. A resurvey of a loblolly pine community twenty-nine years after ground and crown fire. Bull. Torrey Bot. Club 1964, 91, 387–395. [Google Scholar] [CrossRef]
- Wade, D.D.; Weise, D.; Shell, R. Some Effects of Periodic Winter Fire on Communities on the Georgia Piedmont, Proceedings of the 5th Biennial Southern Silvicultural Research Conference, Memphis, TN, USA, 1–3 November 1988; General Tech. Rep. SO-74; Dept. of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1989.
- Cook, E.R.; Meko, D.M.; Stahle, D.W.; Cleaveland, M.K. Drought reconstructions for the continental United States. J. Clim. 1999, 12, 1145–1162. [Google Scholar] [CrossRef]
- USDA Soil Conservation Service and Forest Service. Soil Survey of Baldwin, Jones, and Putnam Counties, Georgia; USDA Soil Conservation Service and Forest Service: Washington, DC, USA, 1976.
- Cowell, C.M. Presettlement Piedmont forests: Patterns in composition and disturbance in central Georgia. Ann. Assoc. Am. Geogr. 1995, 85, 65–83. [Google Scholar]
- Guyette, R.P.; Stambaugh, M.C.; Dey, D.C.; Muzika, R.-M. Predicting fire frequency with chemistry and climate. Ecosystems 2012, 15, 322–335. [Google Scholar] [CrossRef]
- Radford, A.E.; Ahles, H.E.; Bell, C.R. Manual of the Vascular Flora of the Carolinas; The University of North Carolina Press: Chapel Hill, NC, USA, 1968. [Google Scholar]
- Scheiner, S.M. MANOVA: Multiple response variables and multispecies interactions. In The Design and Analysis of Ecological Experiments; Scheiner, S.M., Gurevitch, J., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 134–157. [Google Scholar]
- Von Ende, C.N. Repeated-measures analysis: Growth and other time dependent measures. In The Design and Analysis of Ecological Experiments; Scheiner, S.M., Gurevitch, J., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 134–157. [Google Scholar]
- Dey, D.C.; Hartman, G. Returning fire to Ozark Highland forest ecosystems: Effects on advance regeneration. For. Ecol. Manag. 2005, 217, 37–53. [Google Scholar] [CrossRef]
- McNab, W.H. An overcrowded loblolly pine stand thinned with fire. South. J. Appl. For. 1977, 1, 24–26. [Google Scholar]
- Waldrop, T.A.; Lloyd, F.T. Precommercial thinning a sapling-sized loblolly pine stand with prescribed fire. South. J. Appl. For. 1988, 12, 203–207. [Google Scholar]
- Wade, D.D. Thinning young loblolly pine stands with fire. Int. J. Wildland Fire 1993, 3, 169–178. [Google Scholar] [CrossRef]
- Peet, R.K.; Christensen, N.L. Competition and tree death. Bioscience 1987, 37, 586–595. [Google Scholar] [CrossRef]
- Lotti, T. Eliminating understory hardwoods with summer prescribed fires in Coastal Plain loblolly pine stands. J. For. 1956, 54, 191–192. [Google Scholar]
- Burns, R.M.; Honkala, B.H. Silvics of North America: 2. Hardwoods; Agricultural Handbook; USDA: Washington, DC, USA, 1990.
- Kreye, J.K.; Varner, J.M.; Hiers, J.K.; Mola, J. Toward a mechanism for eastern North American forest mesophication: Differential litter drying across 17 species. Ecol. Appl. 2013, 23, 1976–1986. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Block | Late Spring (4/1 to 5/12) | Early Summer (5/13 to 6/25) | Mid Summer (6/26 to 8/6) | Late Summer (8/7 to 9/18) | |
|---|---|---|---|---|---|
| Block 1 | 1998 | 4/2, 4/27 | 5/13, 6/9 | 7/29, 7/29 | 8/24, 9/9 |
| 2000 | 4/5, 4/26 | 7/261 | 7/26 | ||
| 2001 | 6/7 | 7/12 | 9/18, 9/18 | ||
| 2002 | 4/16, 5/7 | 5/15, 6/5 | 8/1 | ||
| 2003 | 6/26 | 9/10 | |||
| 2004 | 4/15, 4/29 | 5/24 | 8/4 | ||
| Block 2 | 1998 | 4/2, 4/27 | 5/13, 6/8 | 7/21, 7/21 | 8/11, 9/8 |
| 2000 | 4/5, 4/26 | 7/251 | 7/25 | ||
| 2001 | 6/6 | 7/12 | 9/18, 9/18 | ||
| 2002 | 4/16, 5/7 | 5/15, 6/5 | 8/1 | ||
| 2003 | 6/26 | 9/10, 9/10 | |||
| 2004 | 4/19, 4/29 | 5/24 | 8/4 | ||
| Block 3 | 1998 | 4/2, 4/25 | 5/13, 6/8 | 7/22, 8/4 | 8/24, 9/8 |
| 2000 | 4/16, 5/8 | 5/15 | 7/23 | ||
| 2001 | 6/5 | 7/9 | 9/17, 9/17 | ||
| 2002 | 4/16, 5/8 | 5/15, 6/4 | 8/1 | ||
| 2003 | 6/26 | 8/28, 8/28 | |||
| 2004 | 4/19, 4/29 | 5/24 | 8/4 |
| Sapling Density | Time (df = 2) | Block (df = 2) | Time*Block (df = 4) | Win (df = 3) | Time*Win (df = 6) | Block*Win (df = 6) | Time*Block*Win (df = 12) |
|---|---|---|---|---|---|---|---|
| All | F = 49.5 GG = 0.0000* | F = 4.44 p = 0.036* | F = 1.9 GG = 0.152 | F = 1.82 p = 0.197 | F = 1.04 GG = 0.423 | F = 1.53 p = 0.49 | F = 2.63 GG = 0.021* |
| Pine | F = 7.00 GG = 0.021* | F = 7.81 p = 0.0067* | F = 5.16 GG = 0.024* | F = 2.15 p = 0.147 | F = 1.81 GG = 0.198 | F = 2.68 p = 0.069 | F = 2.32 GG = 0.038* |
| Oak Hickory | F = 3.1 GG = 0.1 | F = 8.73 p = 0.005* | F = 1.75 GG = 0.2 | F = 1.88 p = 0.19 | F = 0.70 GG = 0.59 | F = 1.77 p = 0.19 | F = 0.95 GG = 0.5 |
| Mesic Hardwood | F = 36.35 p = 0.0000* | F = 6.25 p = 0.014* | F = 2.67 p = 0.06 | F = 2.32 p = 0.13 | F = 2.02 p = 0.1 | F = 1.56 p = 0.24 | F = 1.62 p = 0.15 |
| Structure/Composition | Time (df = 1) | Block (df = 2) | Time*Block (df = 2) | Win (df = 3) | Time*Win (df = 3) | Block*Win (df = 6) | Time*Block*Win (df = 6) |
|---|---|---|---|---|---|---|---|
| Basal Area | F = 0.57 p = 0.46 | F = 4.06 p = 0.05* | F = 0.1 p = 0.9 | F = 1.07 p = 0.4 | F = 0.77 p = 0.53 | F = 0.71 p = 0.65 | F = 0.45 p = 0.83 |
| PineBasal Area | F = 0.32 p = 0.58 | F = 1.16 p = 0.35 | F = 0.24 p = 0.79 | F = 1.00 p = 0.43 | F = 0.27 p = 0.84 | F = 0.21 p = 0.97 | F = 0.56 p = 0.76 |
| Oak-Hickory Basal Area | F = 0.28 p = 0.61 | F = 0.93 p = 0.42 | F = 1.05 p = 0.38 | F = 1.43 p = 0.28 | F = 0.63 p = 0.61 | F = 0.98 p = 0.48 | F = 2.13 p = 0.12 |
| Mesic Hardwood Basal Area | F = 3.01 p = 0.36 | F = 5.54 p = 0.02* | F = 0.54 p = 0.6 | F = 1.81 p = 0.2 | F = 3.44 p = 0.05* | F = 4.54 p = 0.013* | F = 0.42 p = 0.85 |
| Density | F = 6.45 p = 0.03* | F = 4.47 p = 0.04* | F = 0.29 p = 0.76 | F = 1.79 p = 0.20 | F = 0.43 p = 0.73 | F = 0.64 p = 0.70 | F = 0.72 p = 0.64 |
| Pine Density | F = 8.72 p = 0.012* | F = 0.96 p = 0.41 | F = 1.12 p = 0.36 | F = 1.36 p = 0.30 | F = 0.81 p = 0.51 | F = 0.36 p = 0.89 | F = 1.02 p = 0.46 |
| Oak-Hickory Density | F = 0.05 p = 0.82 | F = 1.22 p = 0.33 | F = 1.32 p = 0.30 | F = 1.65 p = 0.23 | F = 0.05 p = 0.98 | F = 0.64 p = 0.69 | F = 1.11 p = 0.41 |
| Mesic Hardwood Density | F = 2.25 p = 0.16 | F = 4.79 p = 0.03* | F = 0.06 p = 0.94 | F = 2.84 p = 0.08 | F = 2.46 p = 0.11 | F = 3.47 p = 0.03* | F = 1.22 p = 0.36 |
| QMD | F = 24.02 p < 0.001* | F = 0.70 p = 0.52 | F = 0.92 p = 0.43 | F = 0.41 p = 0.75 | F = 0.51 p = 0.68 | F = 0.40 p = 0.87 | F = 0.55 p = 0.76 |
| Pine QMD | F = 36.1 p < 0.001* | F = 0.50 p = 0.62 | F = 1.72 p = 0.22 | F = 0.45 p = 0.72 | F = 0.37 p = 0.78 | F = 0.36 p = 0.89 | F = 0.47 p = 0.82 |
| Oak-Hickory QMD | F = 0.06 p = 0.82 | F = 3.13 p = 0.08 | F = 0.04 p = 0.96 | F = 1.56 p = 0.25 | F = 1.39 p = 0.29 | F = 0.78 p = 0.60 | F = 3.81 p = 0.023* |
| Mesic Hardwood QMD | F = 3.13 p = 0.102 | F = 5.57 p = 0.02* | F = 0.01 p = 0.99 | F = 31.9 p < 0.001* | F = 0.33 p = 0.80 | F = 27.71 p < 0.001* | F = 0.47 p = 0.82 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reilly, M.J.; Outcalt, K.; O’Brien, J.J.; Wade, D. Effects of Repeated Growing Season Prescribed Fire on the Structure and Composition of Pine–Hardwood Forests in the Southeastern Piedmont, USA. Forests 2017, 8, 8. https://doi.org/10.3390/f8010008
Reilly MJ, Outcalt K, O’Brien JJ, Wade D. Effects of Repeated Growing Season Prescribed Fire on the Structure and Composition of Pine–Hardwood Forests in the Southeastern Piedmont, USA. Forests. 2017; 8(1):8. https://doi.org/10.3390/f8010008
Chicago/Turabian StyleReilly, Matthew J., Kenneth Outcalt, Joseph J. O’Brien, and Dale Wade. 2017. "Effects of Repeated Growing Season Prescribed Fire on the Structure and Composition of Pine–Hardwood Forests in the Southeastern Piedmont, USA" Forests 8, no. 1: 8. https://doi.org/10.3390/f8010008
APA StyleReilly, M. J., Outcalt, K., O’Brien, J. J., & Wade, D. (2017). Effects of Repeated Growing Season Prescribed Fire on the Structure and Composition of Pine–Hardwood Forests in the Southeastern Piedmont, USA. Forests, 8(1), 8. https://doi.org/10.3390/f8010008