
A Basin-Scale Estimation of Carbon Stocks of a Forest Ecosystem Characterized by Spatial Distribution and Contributive Features in the Liuxihe River Basin of Pearl River Delta

Abstract
:1. Introduction
2. Materials and Methods
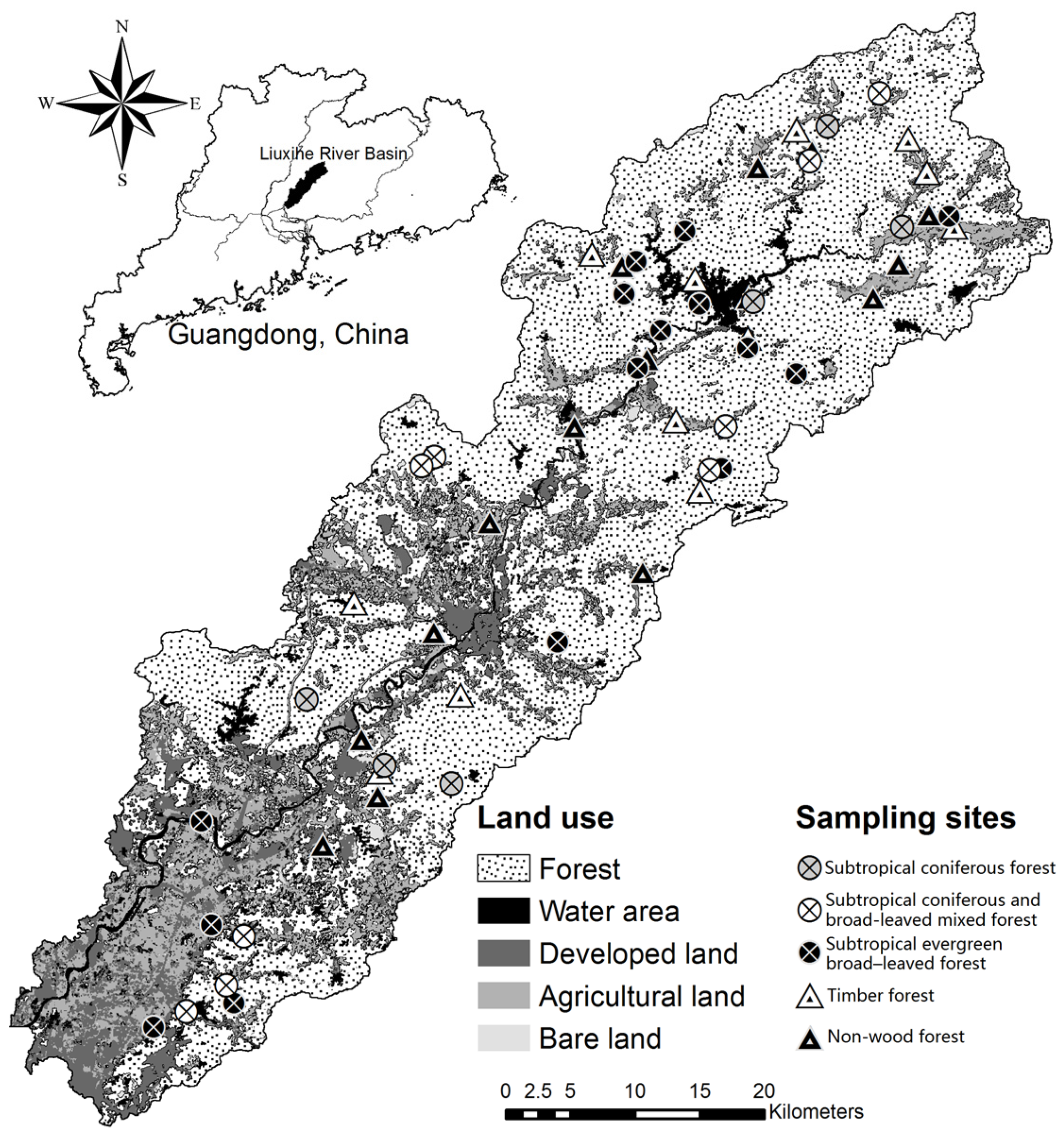
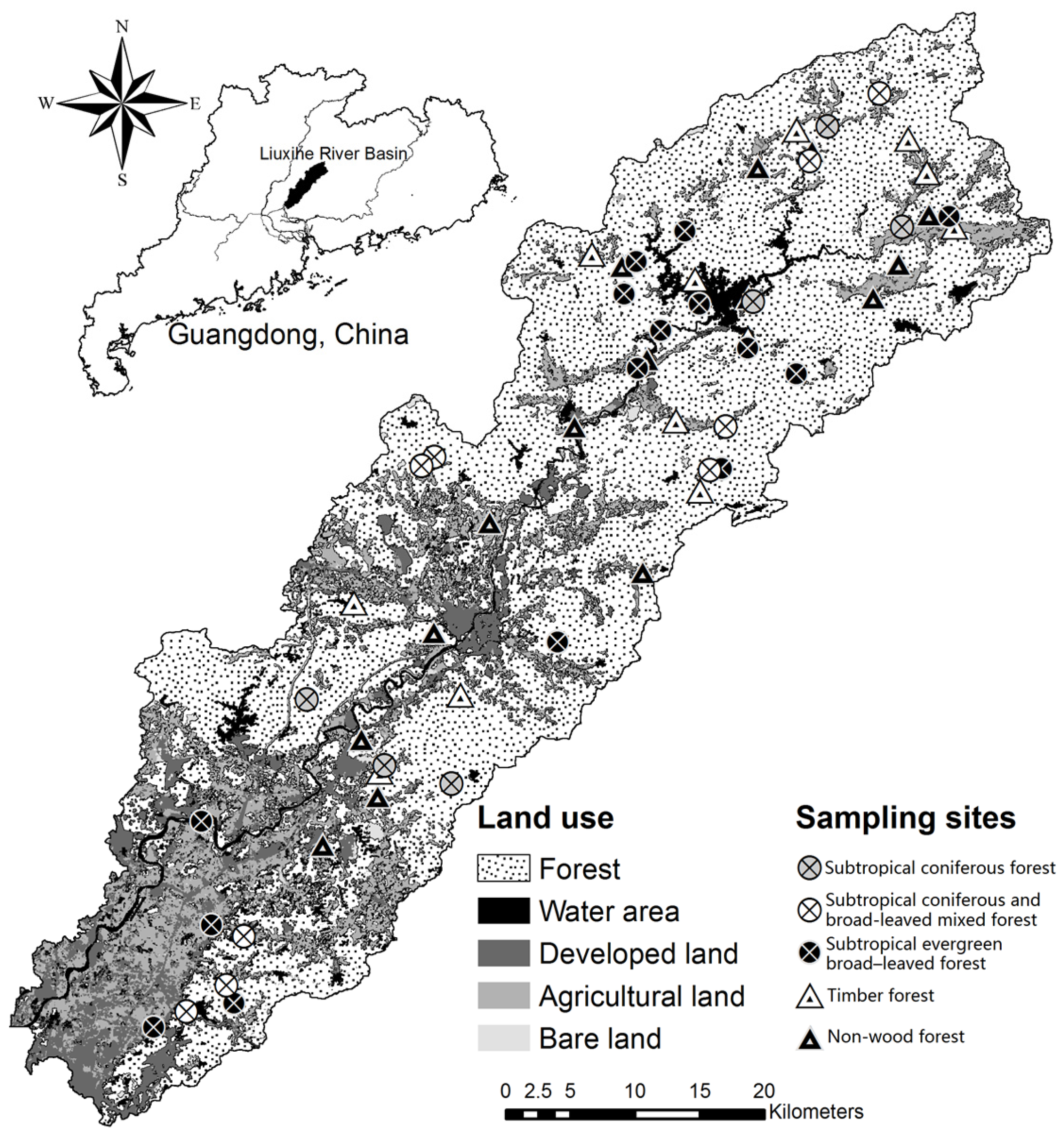
2.1. Study Area and Sample Site
2.2. Sampling and Measurements
2.3. Statistical Analysis
3. Results
3.1. Plant Communities
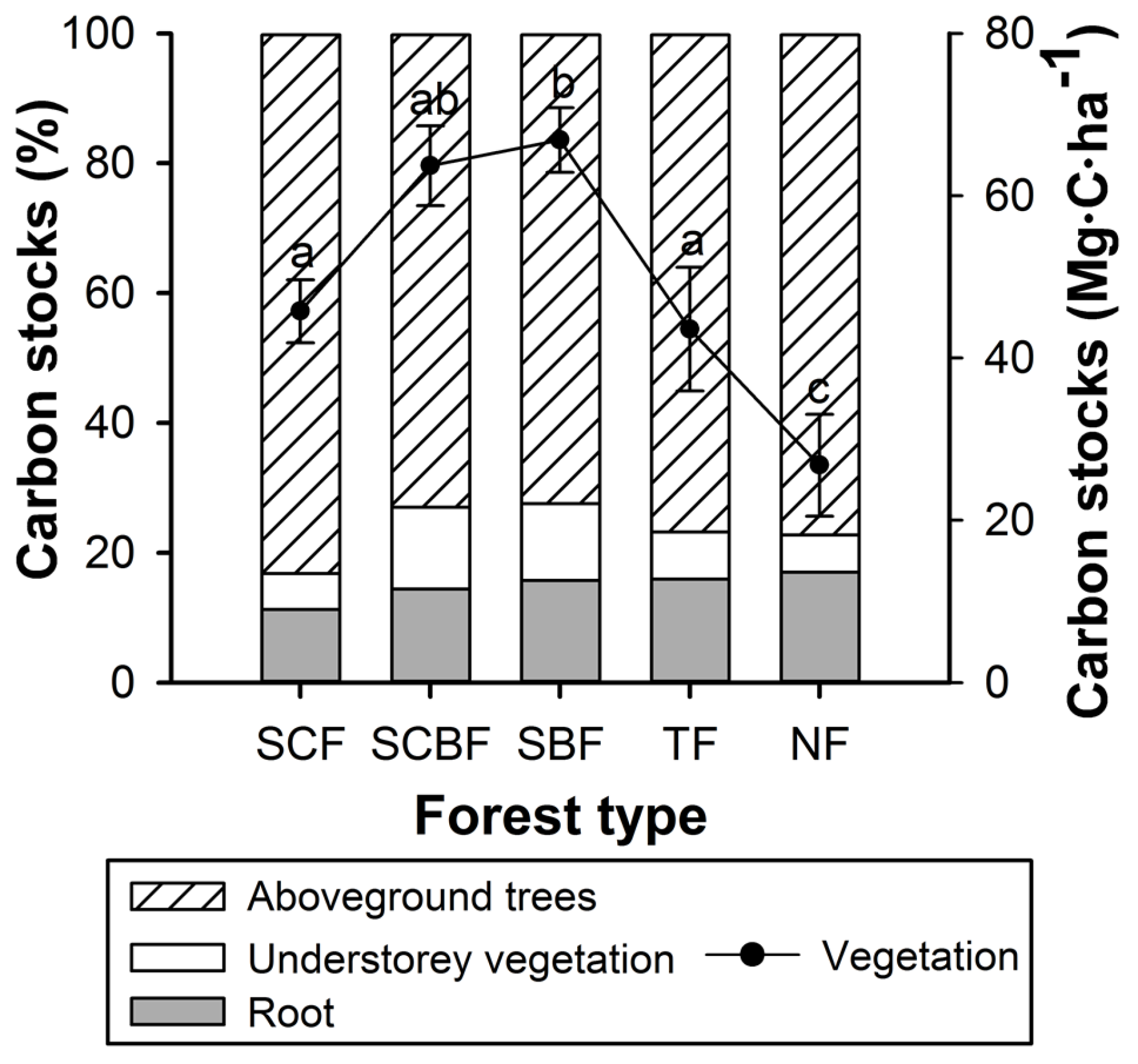
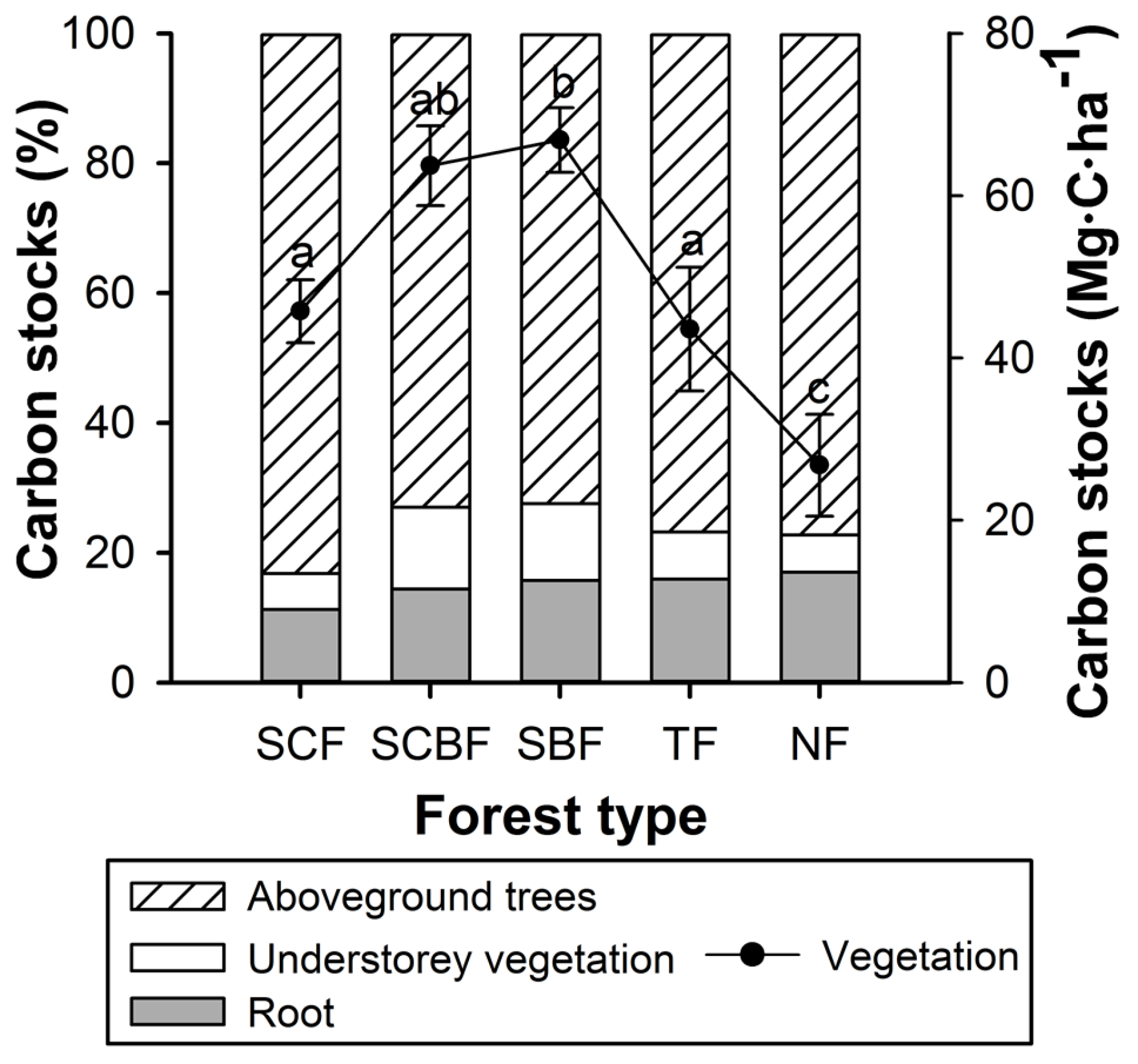
3.2. Vegetation Carbon
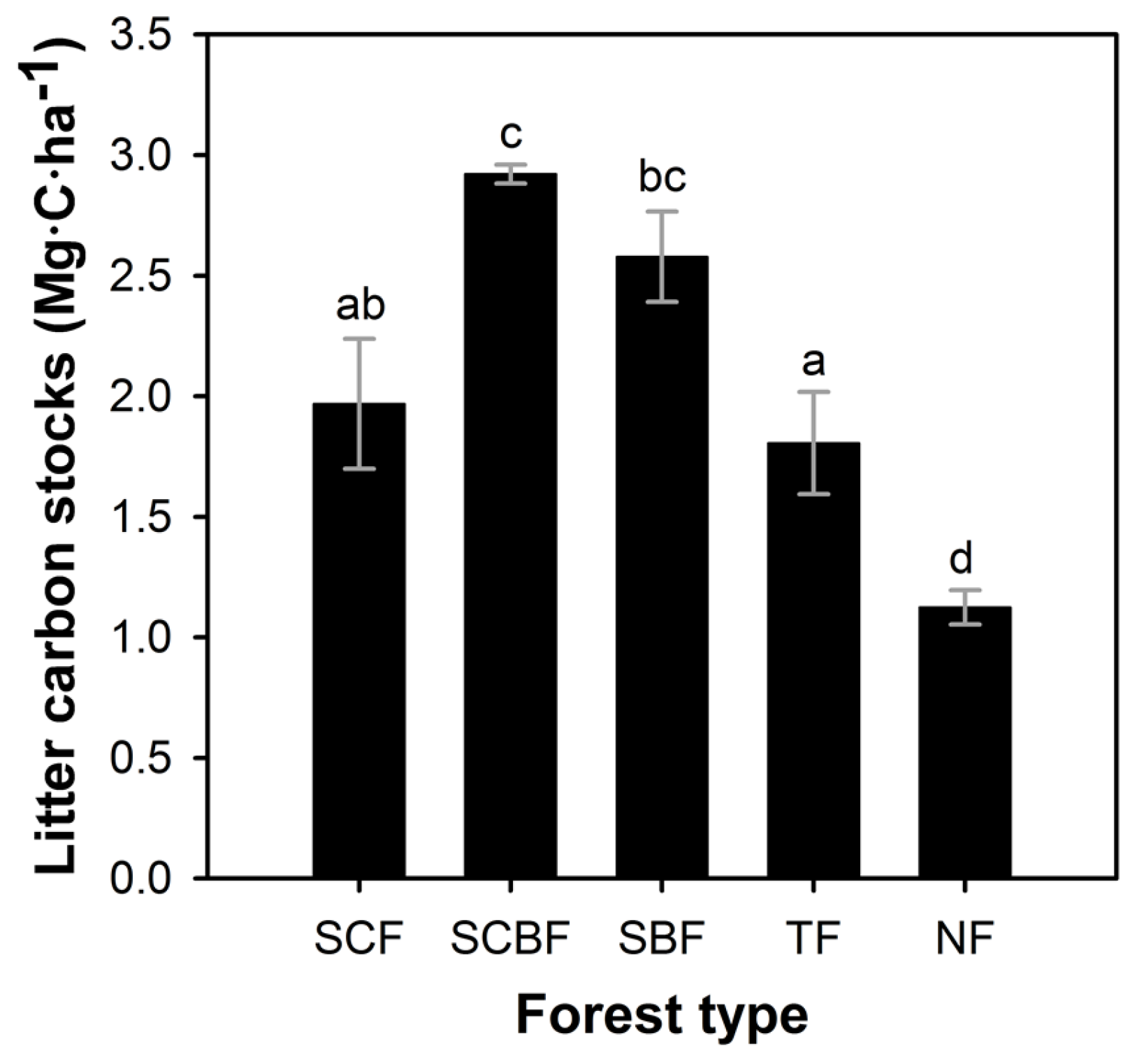
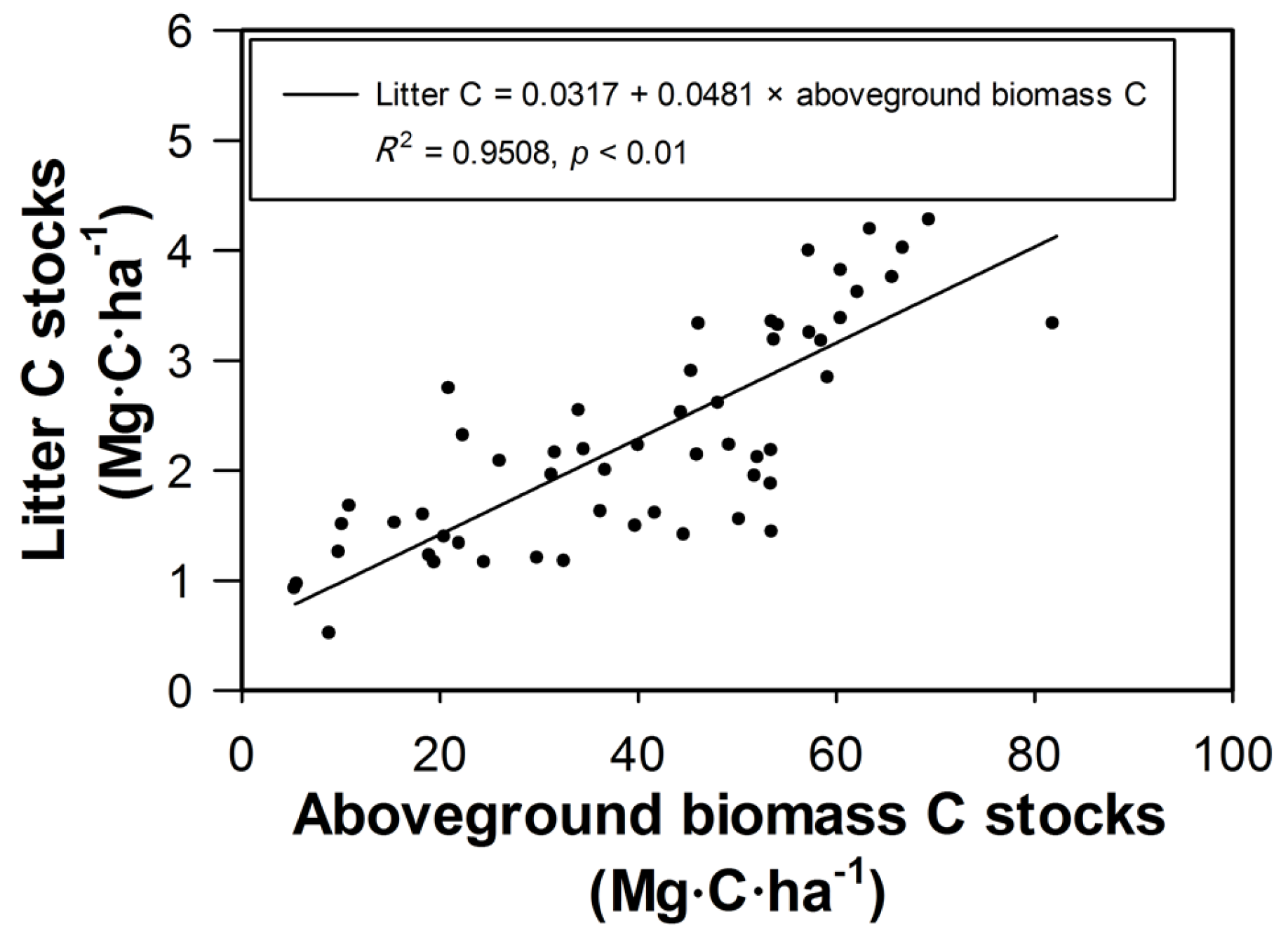
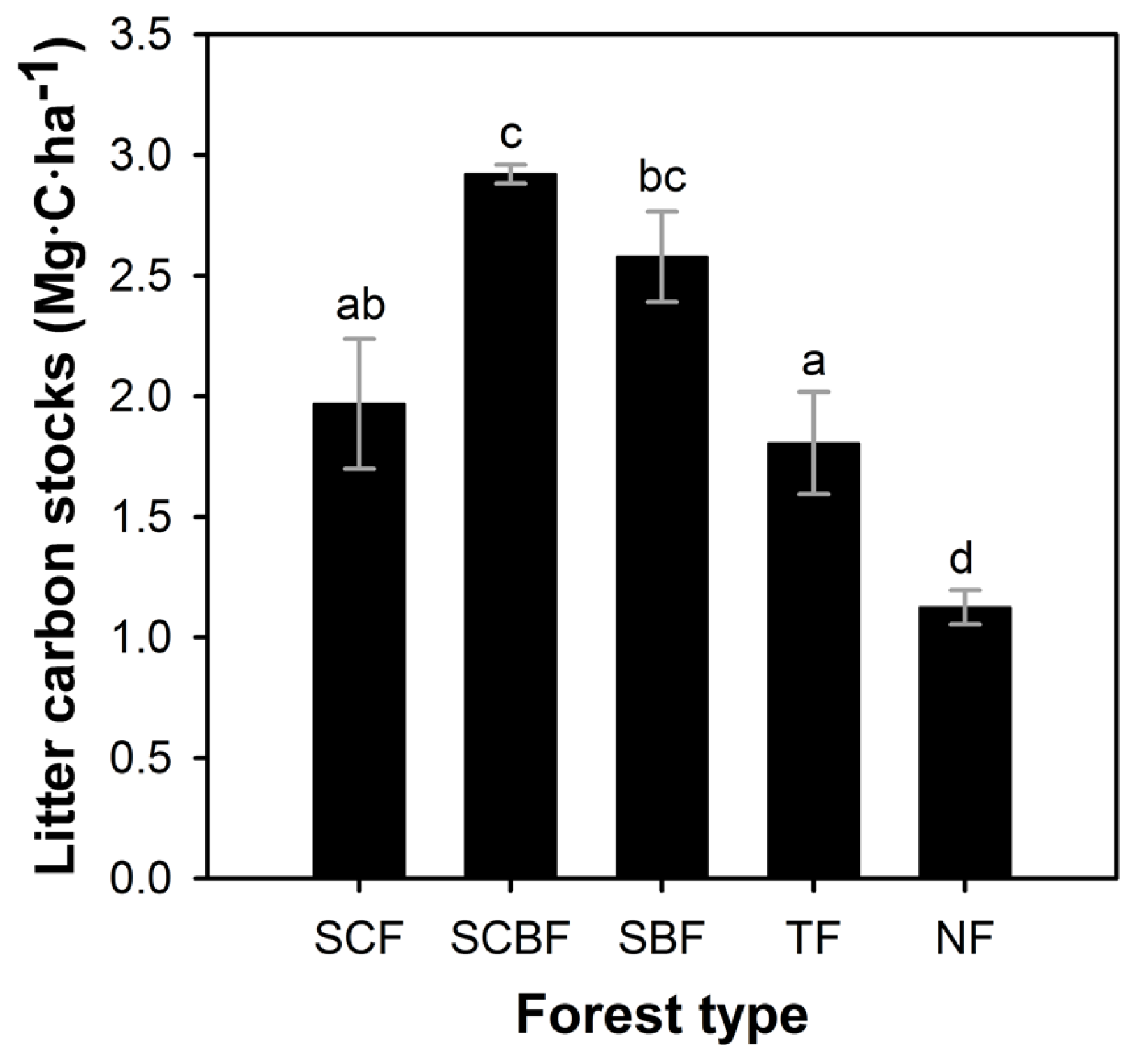
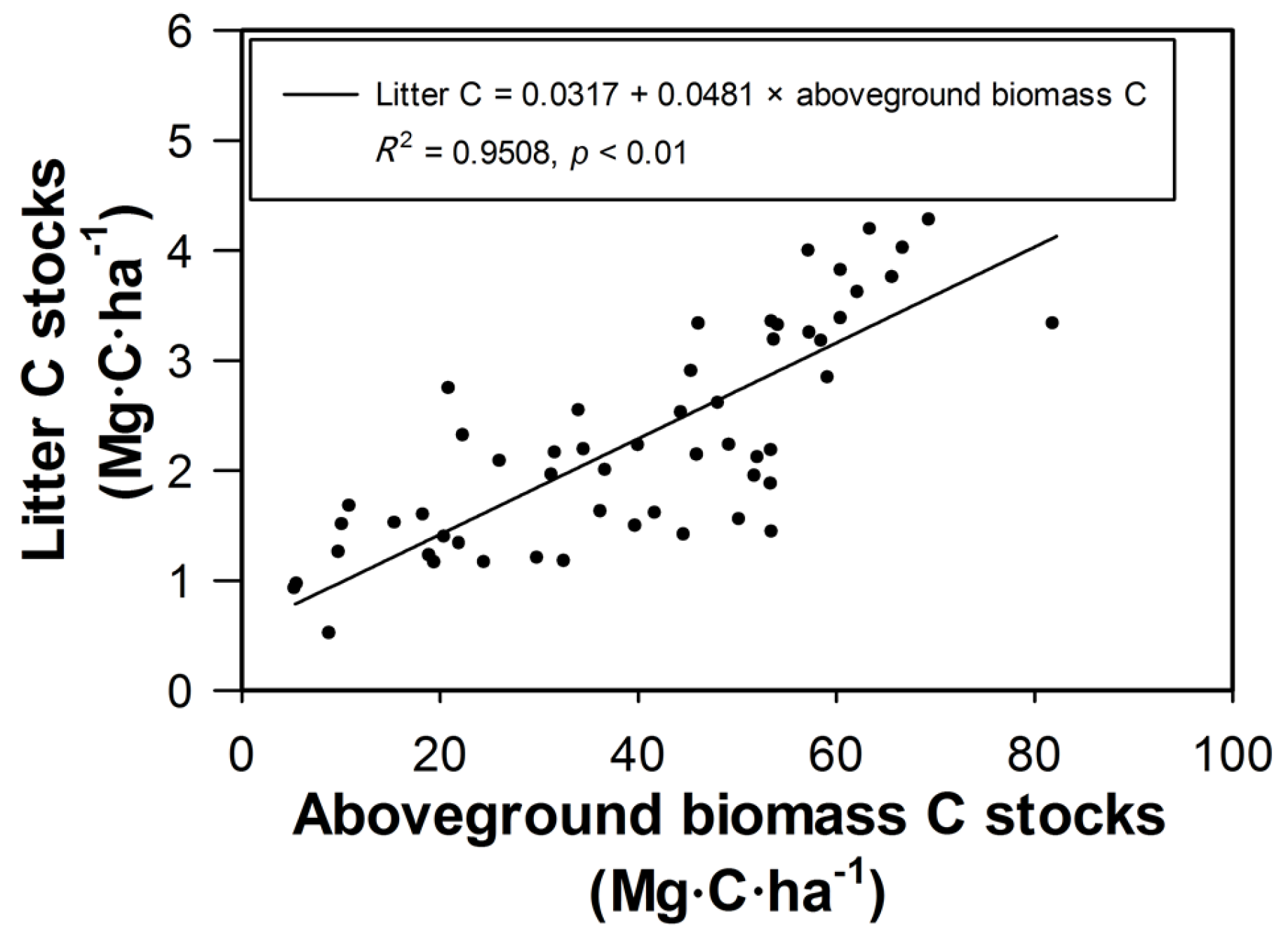
3.3. Litter Carbon
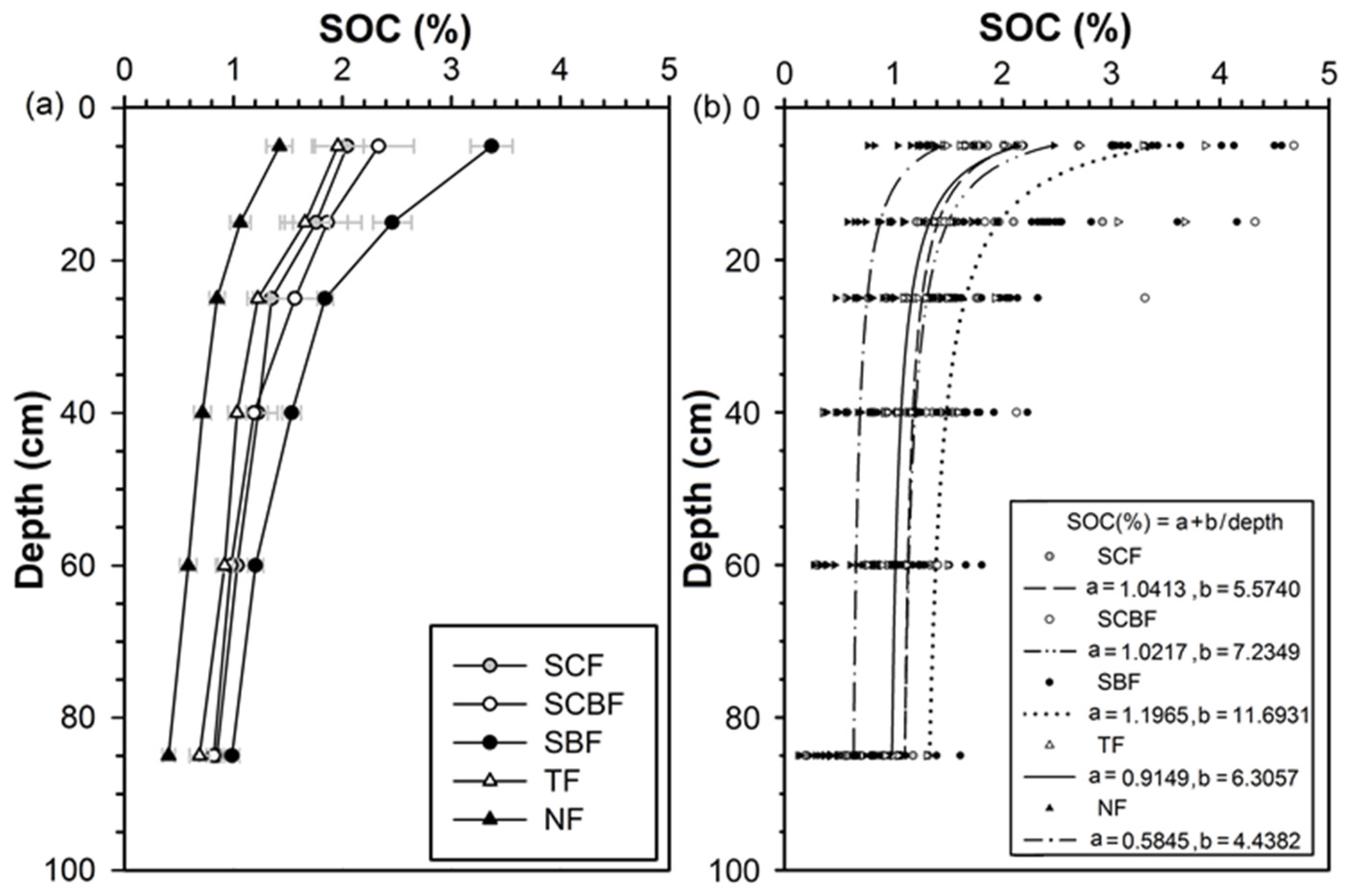
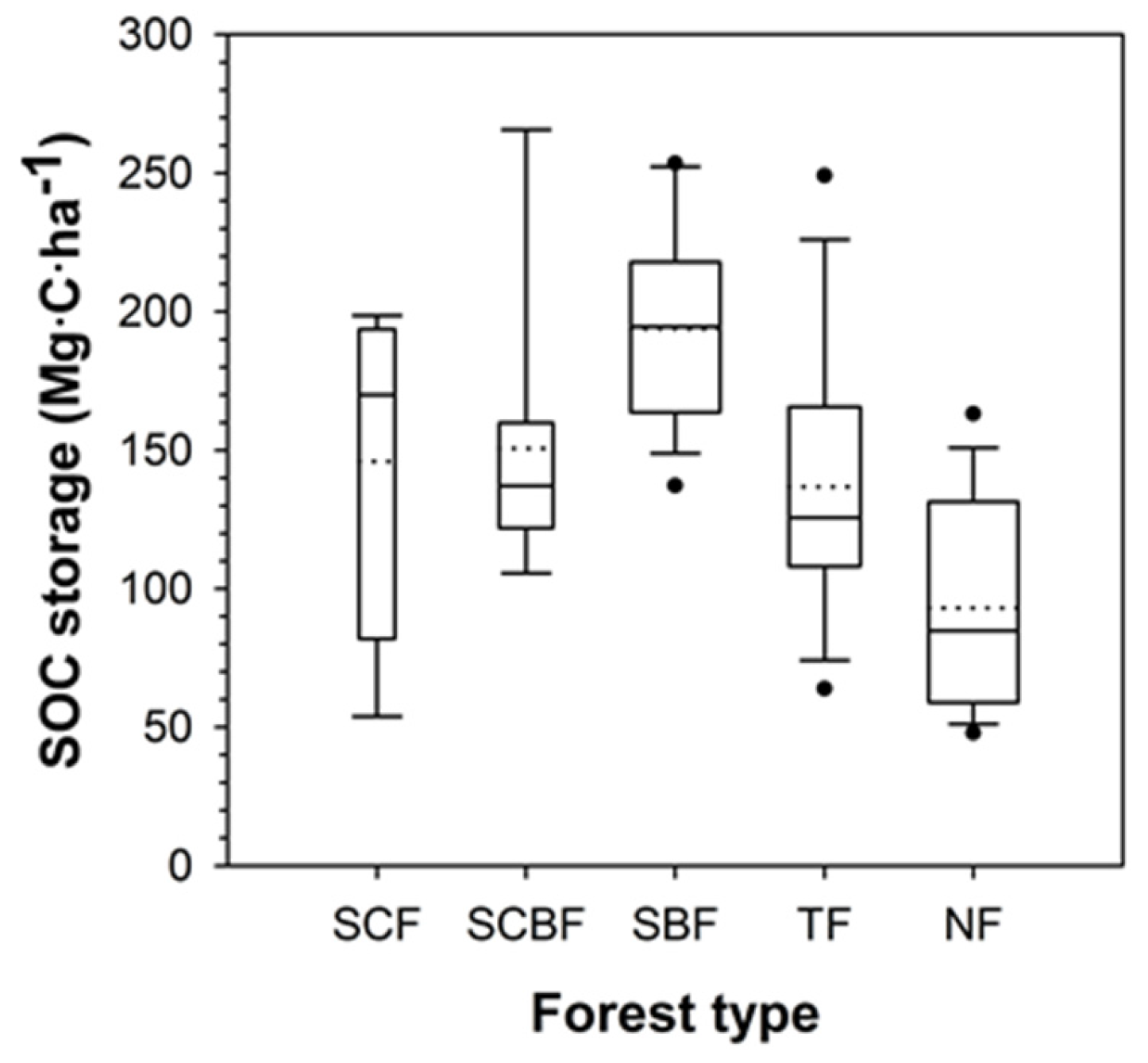
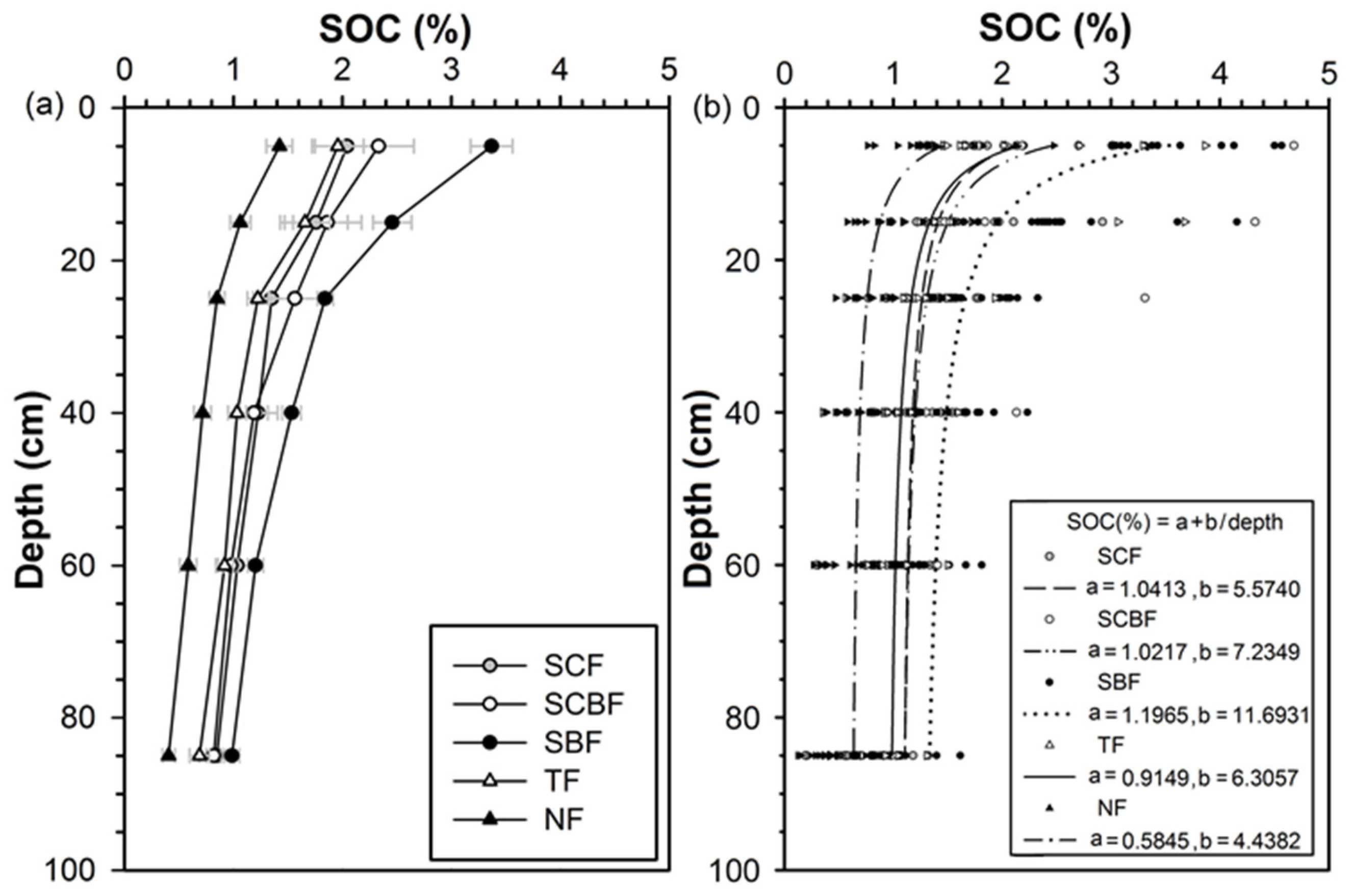
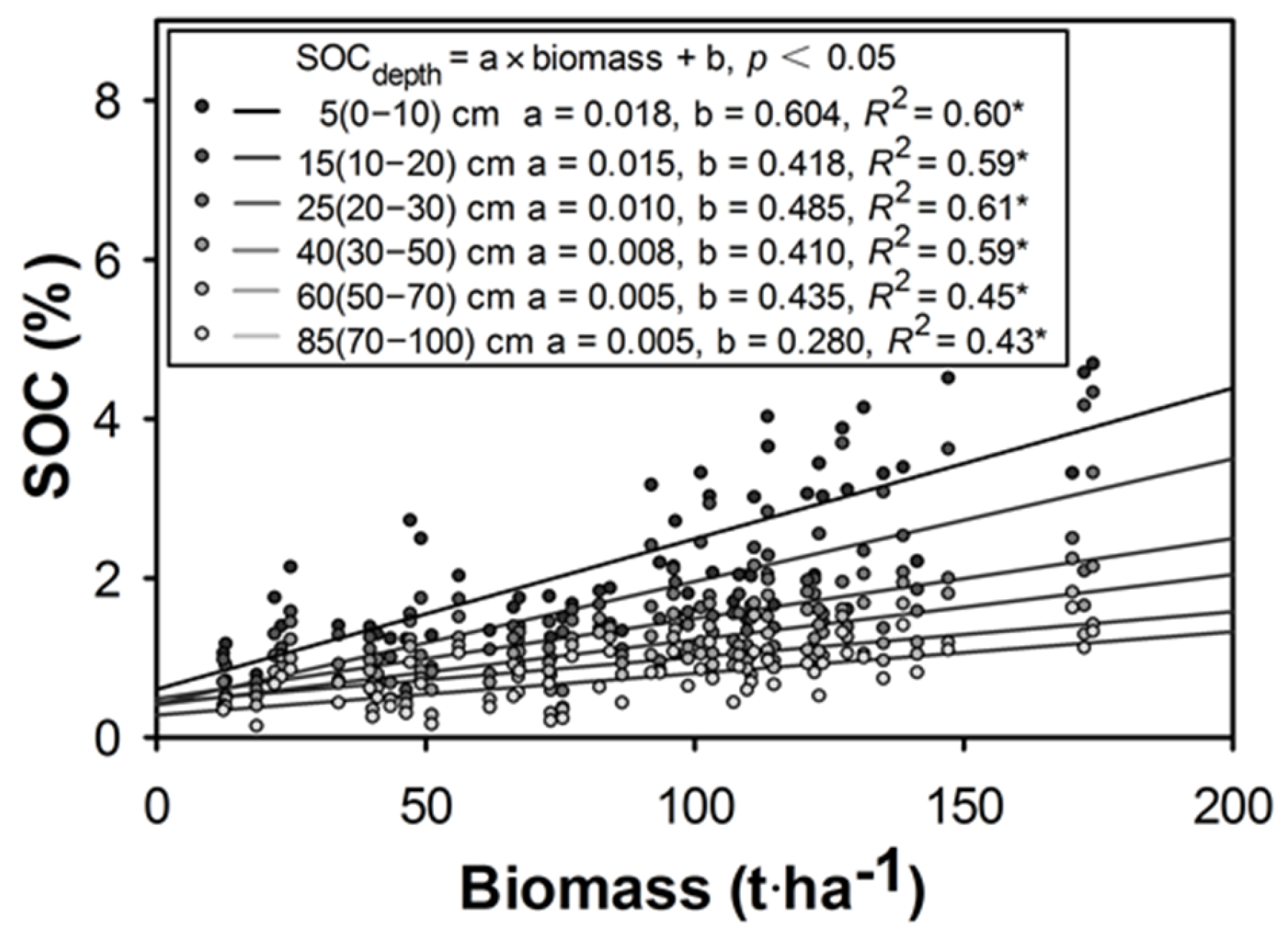
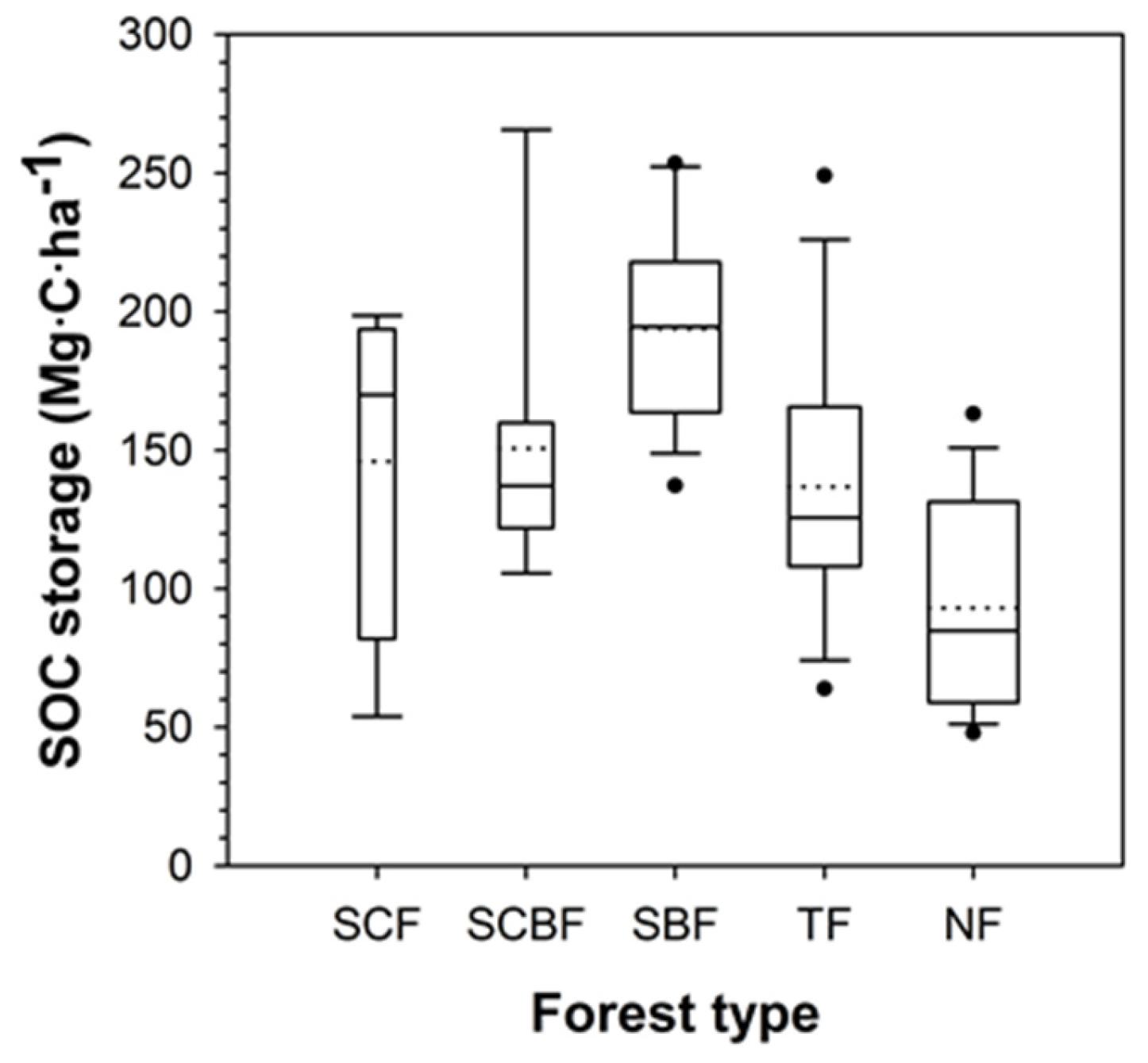
3.4. Soil Organic Carbon
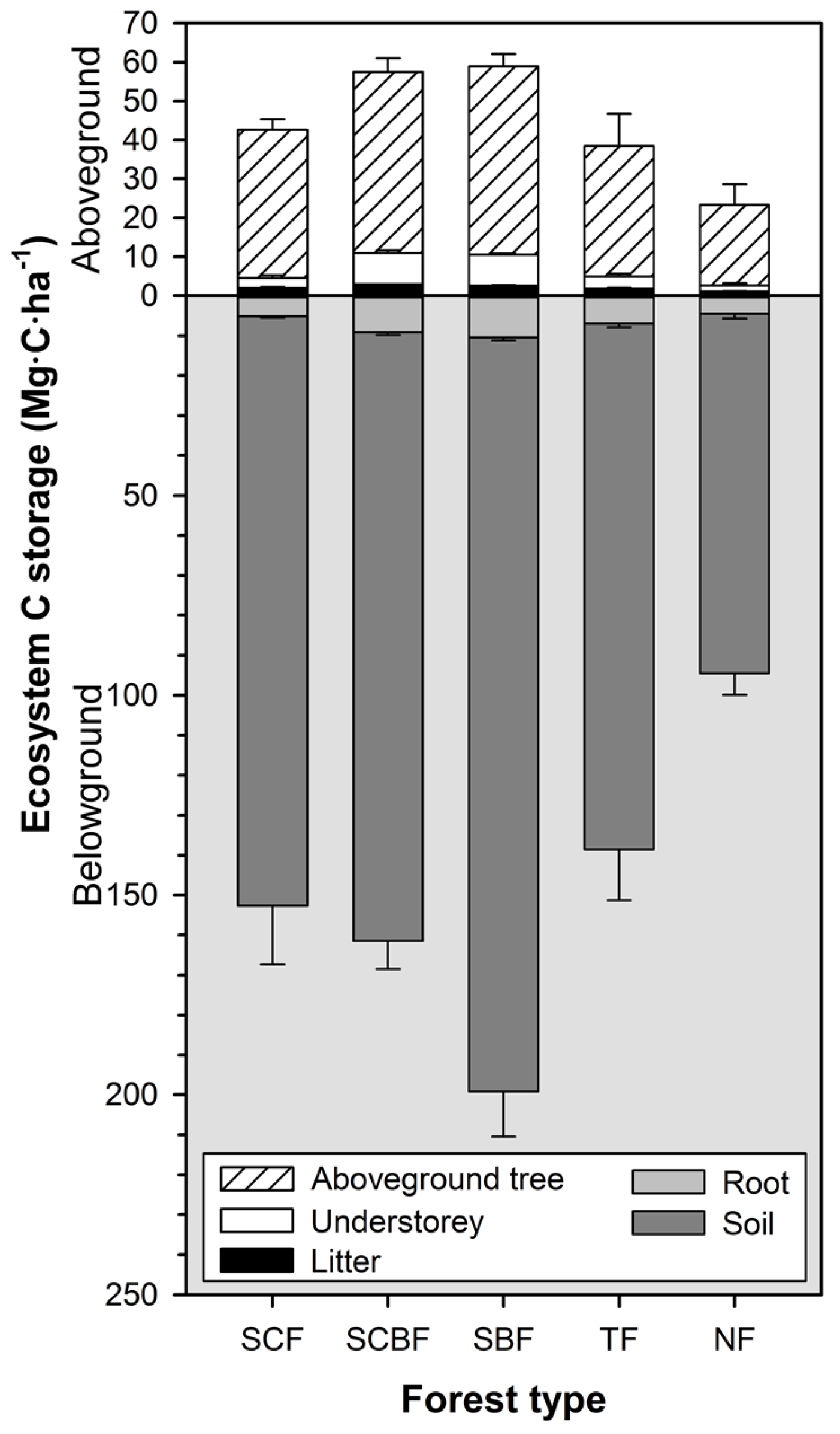
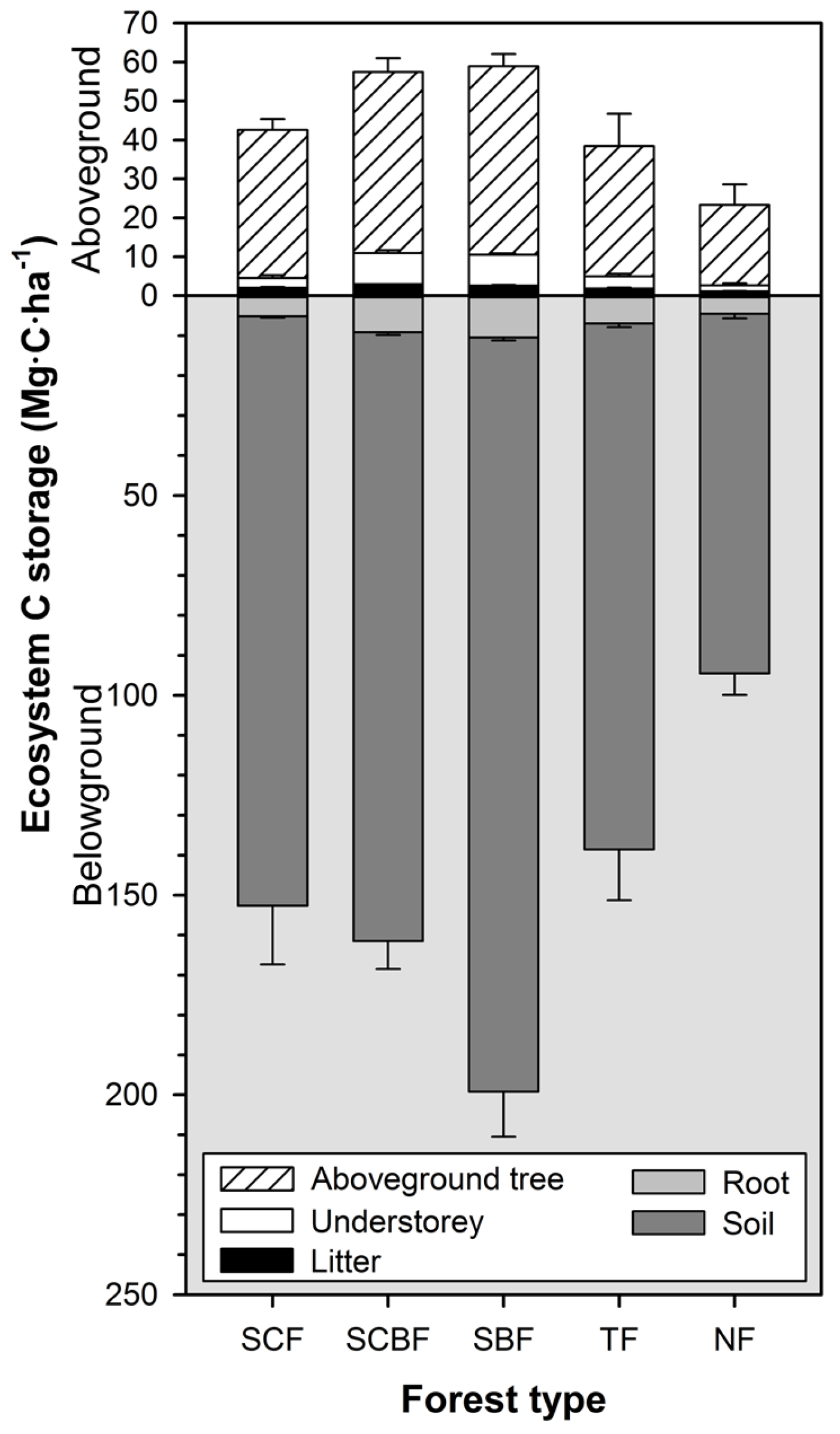
3.5. Structure of Forest Carbon Stocks
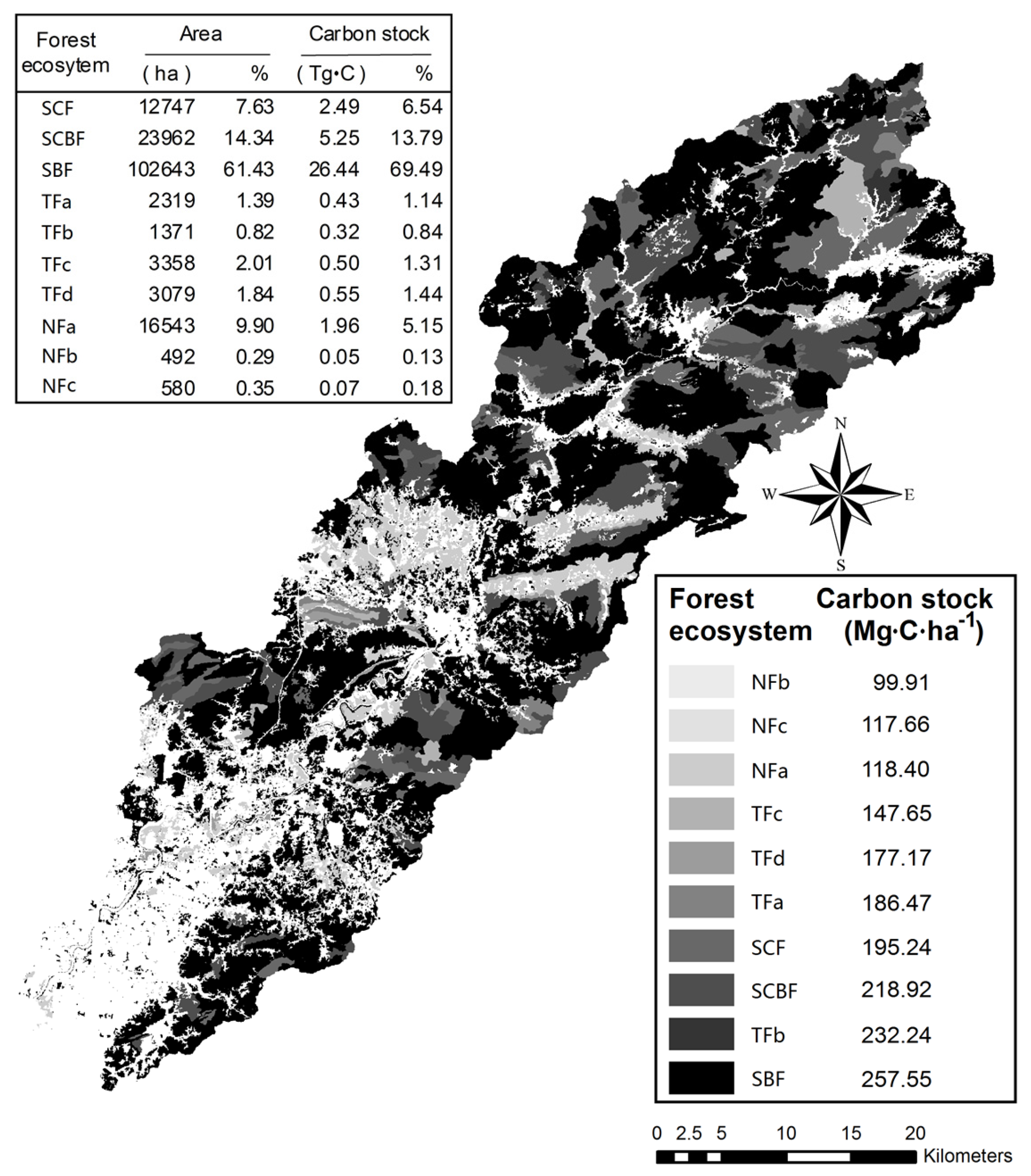
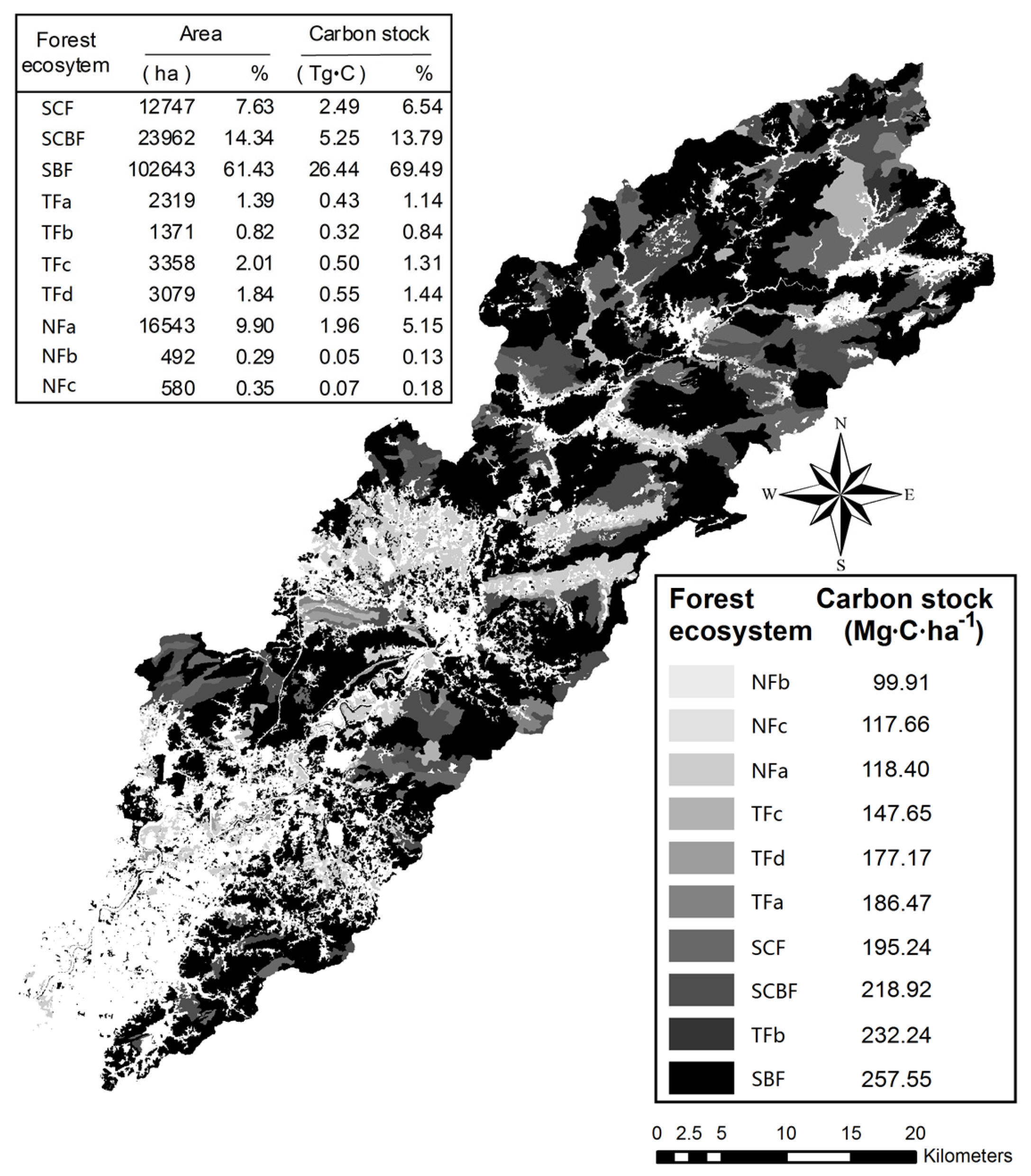
3.6. Spatial Distribution of Forest Ecosystem Carbon Stocks
4. Discussion
4.1. Forest Succession and Age Growth
4.2. Carbon Stock Contributions of Different Forests
4.3. Variation and Distribution of Forest Ecosystem Carbon Stocks
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koh, L.P.; Ghazoul, J. Spatially explicit scenario analysis for reconciling agricultural expansion, forest protection, and carbon conservation in Indonesia. Proc. Natl. Acad. Sci. USA 2010, 107, 11140–11144. [Google Scholar] [CrossRef] [PubMed]
- Aryal, D.R.; De Jong, B.H.J.; Ochoa Gaona, S.; Esparza Olguin, L.; Mendoza Vega, J. Carbon stocks and changes in tropical secondary forests of southern Mexico. Agric. Ecosyst. Environ. 2014, 195, 220–230. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Climate Change 2014: Mitigation of Climate Change; Contribution of Working Group III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Edenhofer, O., Madruge, R.P., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., et al., Eds.; Cambridge University Press: Cambrige, UK; New York, NY, USA, 2014. [Google Scholar]
- Budiharta, S.; Meijaard, E.; Erskine, P.D.; Rondinini, C.; Pacifici, M.; Wilson, K.A. Restoring degraded tropical forests for carbon and biodiversity. Environ. Res. Lett. 2014, 9, 114020. [Google Scholar] [CrossRef]
- Carrasco, L.R.; Papworth, S.K. A ranking of net national contributions to climate change mitigation through tropical forest conservation. J. Environ. Manag. 2014, 146, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Fernandez Romero, M.L.; Lozano Garcia, B.; Parras Alcantara, L. Topography and land use change effects on the soil organic carbon stock of forest soils in Mediterranean natural areas. Agric. Ecosyst. Environ. 2014, 195, 1–9. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Global Forest Resources Assessment 2010: Key Findings; FAO: Roma, Italy, 2010. [Google Scholar]
- Michigan Education Association (MEA). Millennium Ecosystem Assessment; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Asner, G.P. Painting the world REDD: Addressing scientific barriers to monitoring emissions from tropical forests. Environ. Res. Lett. 2011, 6, 021002. [Google Scholar] [CrossRef]
- Drake, J.E.; Davis, S.C.; Raetz, L.M.; DeLucia, E.H. Mechanisms of age-related changes in forest production: The influence of physiological and successional changes. Glob. Chang. Biol. 2011, 17, 1522–1535. [Google Scholar] [CrossRef]
- Ren, Y.; Wei, X.; Wang, D.; Luo, Y.; Song, X.; Wang, Y.; Yang, Y.; Hua, L. Linking landscape patterns with ecological functions: A case study examining the interaction between landscape heterogeneity and carbon stock of urban forests in Xiamen, China. For. Ecol. Manag. 2013, 293, 122–131. [Google Scholar] [CrossRef]
- Trofymow, J.A.; Stinson, G.; Kurz, W.A. Derivation of a spatially explicit 86-year retrospective carbon budget for a landscape undergoing conversion from old-growth to managed forests on Vancouver Island, BC. For. Ecol. Manag. 2008, 256, 1677–1691. [Google Scholar] [CrossRef]
- Jacobi, J.; Andres, C.; Schneider, M.; Pillco, M.; Calizaya, P.; Rist, S. Carbon stocks, tree diversity, and the role of organic certification in different cocoa production systems in Alto Beni, Bolivia. Agrofor. Syst. 2014, 88, 1117–1132. [Google Scholar] [CrossRef]
- Profft, I.; Mund, M.; Weber, G.; Weller, E.; Schulze, E. Forest management and carbon sequestration in wood products. Eur. J. For. Res. 2009, 128, 399–413. [Google Scholar] [CrossRef]
- Stoate, C.; Báldi, A.; Beja, P.; Boatman, N.D.; Herzon, I.; van Doorn, A.; de Snoo, G.R.; Rakosy, L.; Ramwell, C. Ecological impacts of early 21st century agricultural change in Europe—A review. J. Environ. Manag. 2009, 91, 22–46. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.J.; Ingram, J.S.I. Global change and food and forest production: Future scientific challenges. Agric. Ecosyst. Environ. 2001, 82, 3–14. [Google Scholar] [CrossRef]
- Winjum, J.K.; Brown, S.; Schlamadinger, B. Forest harvests and wood products: Sources and sinks of atmospheric carbon dioxide. For. Sci. 1998, 44, 272–284. [Google Scholar]
- Rosenqvist, A.; Milne, A.; Lucas, R.; Imhoff, M.; Dobson, C. A review of remote sensing technology in support of the Kyoto Protocol. Environ. Sci. Policy 2003, 6, 441–455. [Google Scholar] [CrossRef]
- Fu, T.; Zhu, J.; Xiao, W.; Zeng, L. Above-ground biomass distribution models for arbor layer of eight subtropical forest types. Sci. Silv. Sin. 2014, 50, 1–9. [Google Scholar]
- Mbaabu, P.R.; Hussin, Y.A.; Weir, M.; Gilani, H. Quantification of carbon stock to understand two different forest management regimes in Kayar Khola watershed, Chitwan, Nepal. J. Indian Soc. Remote Sens. 2014, 42, 745–754. [Google Scholar] [CrossRef]
- Graae, B.J.; Okland, R.; Petersen, P.M.; Jensen, K.; Fritzboger, B. Influence of historical, geographical understory composition and environmental variables on richness in Danish forests. J. Veg. Sci. 2004, 15, 465–474. [Google Scholar] [CrossRef]
- Bruelheide, H.; Udelhoven, P. Correspondence of the fine-scale spatial variation in soil chemistry and the herb layer vegetation in beech forests. For. Ecol. Manag. 2005, 210, 205–223. [Google Scholar] [CrossRef]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. OIKOS 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Guo, L.B.; Sims, R. Litter production and nutrient return in New Zealand eucalypt short-rotation forests: Implications for land management. Agric. Ecosyst. Environ. 1999, 73, 93–100. [Google Scholar] [CrossRef]
- Sitch, S.; Smith, B.; Prentice, I.C.; Arneth, A.; Bondeau, A.; Cramer, W.; Kaplan, J.O.; Levis, S.; Lucht, W.; Sykes, M.T.; et al. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Glob. Chang. Biol. 2003, 9, 161–185. [Google Scholar] [CrossRef]
- Yang, K.; Zhu, J. Impact of tree litter decomposition on soil biochemical properties obtained from a temperate secondary forest in Northeast China. J. Soil Sediment 2015, 15, 13–23. [Google Scholar] [CrossRef]
- Prentice, I.C. The Carbon Cycle and Atmospheric Carbon Dioxide; Climate Change 2001: The Scientific Basis Cambridge University Press: Cambridge, UK, 2001; pp. 183–237. [Google Scholar]
- Dixon, R.K.; Brown, S.; Houghton, R.A.; Solomon, A.M.; Trexler, M.C.; Wisniewski, J. Carbon pools and flux of global forest ecosystems. Science 1994, 263, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.A.; Kolka, R.K. Soil carbon storage estimation in a forested watershed using quantitative soil-landscape modeling. Soil Sci. Soc. Am. J. 2005, 69, 1086–1093. [Google Scholar] [CrossRef]
- Zushi, K. Spatial distribution of soil carbon and nitrogen storage and forest productivity in a watershed planted to Japanese cedar (Cryptomeria japonica D. Don). J. For. Res. 2006, 11, 351–358. [Google Scholar] [CrossRef]
- Mueller, A.L.; Dalmago, G.A.; Cunha Fernandes, J.M.; Da Cunha, G.R.; Santi, A.; Kovaleski, S.; Bolis, L.M.; Schweig, E.; Fochesatto, E. Growth and biomass accumulation in mixed araucaria forest in southern Brazil. Rev. Arvore 2014, 38, 221–231. [Google Scholar]
- Schlesinger, W.H. Evidence from chronosequence studies for a low carbon storage potential of soils. Nature 1990, 348, 232–234. [Google Scholar] [CrossRef]
- Houghton, R.A.; Davidson, E.A.; Woodwell, G.M. Missing sinks, feedbacks, and understanding the role of terrestrial ecosystems in the global carbon balance. Glob. Biogeochem. Cycles 1998, 12, 25–34. [Google Scholar] [CrossRef]
- Department of Energy. Carbon Sequestration: Research and Development; A US Department of Energy Report; Office of Science and Office of Fossil Energy, National Technical Information Service: Springfield, VA, USA, 1999.
- Black, K.; Creamer, R.E.; Xenakis, G.; Cook, S. Improving forest soil carbon models using spatial data and geostatistical approaches. Geoderma 2014, 487–499. [Google Scholar] [CrossRef]
- Zhang, H.; Song, T.; Wang, K.; Du, H.; Yue, Y.; Wang, G.; Zeng, F. Biomass and carbon storage in an age-sequence of Cyclobalanopsis glauca plantations in southwest China. Ecol. Eng. 2014, 73, 184–191. [Google Scholar] [CrossRef]
- Schulp, C.J.E.; Nabulars, G.; Verburg, P.H.; De Waal, R.W. Effect of tree species on carbon stocks in forest floor and mineral soil and implications for soil carbon inventories. For. Ecol. Manag. 2008, 256, 482–490. [Google Scholar] [CrossRef]
- Diaz Pines, E.; Schindlbacher, A.; Godino, M.; Kitzler, B.; Jandl, R.; Zechmeister Boltenstern, S.; Rubio, A. Effects of tree species composition on the CO2 and N2O efflux of a Mediterranean mountain forest soil. Plant Soil 2014, 384, 243–257. [Google Scholar] [CrossRef]
- Johnson, D.W.; Todd, D.E.; Trettin, C.F.; Sedinger, J.S. Soil carbon and nitrogen changes in forests of Walker Branch watershed, 1972 to 2004. Soil Sci. Soc. Am. J. 2007, 71, 1639–1646. [Google Scholar] [CrossRef]
- Liski, J.; Perruchoud, D.; Karjalainen, T. Increasing carbon stocks in the forest soils of Western Europe. For. Ecol. Manag. 2002, 169, 159–175. [Google Scholar] [CrossRef]
- Feng, Z.W.; Wang, X.K.; Wu, G. The Forest Ecosystem Biomass and Productivity in China; Science Press: Beijing, China, 1999. [Google Scholar]
- Li, Y.; Zhang, J.; Duan, A.; Xiang, C. Selection of biomass estimation models for Chinese fir plantation. CHN J. Appl. Ecol. 2010, 21, 3036–3046. [Google Scholar]
- Chen, Y.; Liu, Z.; Rao, X.; Wang, X.; Liang, C.; Lin, Y.; Zhou, L.; Cai, X.; Fu, S. Carbon storage and allocation pattern in plant biomass among different forest plantation stands in Guangdong, China. Forests 2015, 6, 794–808. [Google Scholar] [CrossRef]
- Bao, F.C.; Jiang, Z.H. Wood Properties of Main Tree Species from Plantation in China; China Forestry Publishing House: Beijing, China, 1998. [Google Scholar]
- Jiang, Z.H.; Peng, Z.H. Wood Properties of the Global Important Tree Species; Science Press: Beijing, China, 2001. [Google Scholar]
- Intergovernmental Panel on Climate Change (IPCC). Good Practice Guidance for Land Use, Land-use Change and Forestry; Institute for Global Environmental Strategies (IGES) for the IPCC: Kanagawa, Japan, 2003. [Google Scholar]
- Jiang, P.K.; Xu, Q.F. Abundance and dynamics of soil labile carbon pools under different types of forest vegetation. Pedosphere 2006, 16, 505–511. [Google Scholar] [CrossRef]
- Van Breugel, M.; Martinez Ramos, M.; Bongers, F. Community dynamics during early secondary succession in Mexican tropical rain forests. J. Trop. Ecol. 2006, 22, 663–674. [Google Scholar] [CrossRef]
- Lebrija Trejos, E.; Meave, J.A.; Poorter, L.; Pérez García, E.A.; Bongers, F. Pathways, mechanisms and predictability of vegetation change during tropical dry forest succession. Perspec. Plant Ecol. 2010, 12, 267–275. [Google Scholar] [CrossRef]
- Finegan, B.; Delgado, D. Structural and floristic heterogeneity in a 30-year-old costa rican rain forest restored, on pasture through natural secondary succession. Restor. Ecol. 2000, 8, 380–393. [Google Scholar] [CrossRef]
- Hao, Z.Q.; Zhang, J.; Li, B.H.; Ye, J.; Wang, X.G.; Yao, X.L. Natural secondary poplar-birch forest in Changbai Mountain: Species composition and community structure. Chin. J. Plant Ecol. 2008, 32, 238–250. [Google Scholar]
- Fahey, T.J.; Woodbury, P.B.; Battles, J.J.; Goodale, C.L.; Hamburg, S.P.; Ollinger, S.V.; Woodall, C.W. Forest carbon storage: Ecology, management, and policy. Front. Ecol. Environ. 2010, 8, 245–252. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, K.S.; Euskirchen, E.S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Glob. Chang. Biol. 2004, 10, 2052–2077. [Google Scholar] [CrossRef]
- Fang, Y.T.; Mo, J.M.; Peng, S.L.; Li, D.J. Role of forest succession on carbon sequestration of forest ecosystems in lower subtropical China. Acta Ecol. Sin. 2003, 23, 1685–1694. [Google Scholar]
- Tang, X.L.; Zhou, G.Y.; Wen, D.Z.; Zhang, D.Q.; Yan, J.H. Distribution of carbon storage in a lower subtropical monsoon evergreen broad-leaved forest in Dinghushan Nature Reserve. Acta Ecol. Sin. 2003, 23, 90–97. [Google Scholar]
- Muukkonen, P.; Makipaa, R. Empirical biomass models of understory vegetation in boreal forests according to stand and site attributes. Boreal Environ. Res. 2006, 11, 355–369. [Google Scholar]
- Tolunay, D. Carbon concentrations of tree components, forest floor and understory in young Pinus sylvestris stands in north-western Turkey. Scand. J. For. Res. 2009, 24, 394–402. [Google Scholar] [CrossRef]
- Van Calster, H.; Baeten, L.; De Schrijver, A.; De Keersmaeker, L.; Rogister, J.E.; Verheyen, K.; Hermy, M. Management driven changes (1967–2005) in soil acidity and the understory plant community following conversion of a coppice-with-standards forest. For. Ecol. Manag. 2007, 241, 258–271. [Google Scholar] [CrossRef]
- Manninen, O.H.; Stark, S.; Kytoviita, M.M.; Lampinen, L.; Tolvanen, A. Understory plant and soil responses to disturbance and increased nitrogen in boreal forests. J. Veg. Sci. 2009, 20, 311–322. [Google Scholar] [CrossRef]
- Gakis, S.F.; Orfanoudakis, M.Z.; Papaioannou, A.G.; Mantzanas, K.T.; Papanastasis, V.P.; Alifragis, D.A.; Seilopoulos, D.G.; Kostakis, S.N. Long term evolution of tree growth, understory vegetation and soil properties in a silvopastoral system of northern Greece. Ann. For. Res. 2014, 57, 247–265. [Google Scholar]
- Muukkonen, P.; Makipaa, R.; Laiho, R.; Minkkinen, K.; Vasander, H.; Finer, L. Relationship between biomass and percentage cover in understory vegetation of boreal coniferous forests. Silva Fenn. 2006, 40, 231–245. [Google Scholar] [CrossRef]
- Woziwoda, B.; Parzych, A.; Kopec, D. Species diversity, biomass accumulation and carbon sequestration in the understory of post-agricultural Scots pine forests. Silva Fenn. 2014, 48, 1119. [Google Scholar] [CrossRef]
- Hansen, R.A.; Coleman, D.C. Litter complexity and composition are determinants of the diversity and species composition of oribatid mites (Acari: Oribatida) in litterbags. Appl. Soil Ecol. 1998, 9, 17–23. [Google Scholar] [CrossRef]
- Li, X.; Yin, X.; Wang, Z.; Fan, W. Litter mass loss and nutrient release influenced by soil fauna of Betula ermanii forest floor of the Changbai Mountains, China. Appl. Soil Ecol. 2015, 95, 15–22. [Google Scholar] [CrossRef]
- Attiwill, P.M. The disturbance of forest ecosystems: The ecological basis for conservative management. For. Ecol. Manag. 1994, 63, 247–300. [Google Scholar] [CrossRef]
- Roberts, M.R.; Gilliam, F.S. Patterns and mechanisms of plant diversity in forested ecosystems: Implications for forest management. Ecol. Appl. 1995, 5, 969–977. [Google Scholar] [CrossRef]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of European forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Peng, S.L.; Ren, H.; Wu, J.G.; Lu, H.F. Effects of litter removal on plant species diversity a case study in tropical Eucalyptus forest ecosystems in South China. Chin. J. Environ. Sci. 2003, 15, 367–371. [Google Scholar]
- Borchers, J.G.; Perry, D.A. The influence of soil texture and aggregation on carbon and nitrogen dynamics in southwest Oregon forests and clear cuts. Can. J. For. Res. 1992, 22, 298–305. [Google Scholar] [CrossRef]
- Powers, J.S.; Schlesinger, W.H. Relationships among soil carbon distributions and biophysical factors at nested spatial scales in rain forests of northeastern Costa Rica. Geoderma 2002, 109, 165–190. [Google Scholar] [CrossRef]
- Pouyat, R.; Groffman, P.; Yesilonis, I.; Hernandez, L. Soil carbon pools and fluxes in urban ecosystems. Environ. Pollut. 2002, 1161, S107–S118. [Google Scholar] [CrossRef]
- Dilustro, J.J.; Collins, B.S.; Duncan, L.K.; Sharitz, R.R. Soil texture, land-use intensity, and vegetation of Fort Benning upland forest sites. J. Torrey Bot. Soc. 2002, 129, 289–297. [Google Scholar] [CrossRef]
- Jiang, J.P.; Xiong, Y.C.; Jiang, H.M.; Ye, D.; Song, Y.J.; Li, F.M. Soil microbial activity during secondary vegetation succession in semiarid abandoned lands of loess plateau. Pedosphere 2009, 19, 735–747. [Google Scholar] [CrossRef]
- Zech, W.; Guggenberger, G.; Schulten, H. Budgets and chemistry of dissolved organic carbon in forest soils: Effects of anthropogenic soil acidification. Sci. Total Environ. 1994, 152, 49–62. [Google Scholar] [CrossRef]
- Larionova, A.A.; Rozanova, L.N.; Evdokimov, I.V.; Ermolaev, A.M. Carbon budget in natural and anthropogenic forest-steppe ecosystems. Eurasian Soil Sci. 2002, 35, 156–164. [Google Scholar]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Hao, J.; Zhao, S.; Sang, Y.; Wang, H.; Di, X. Temporal and spacial changes of total soil organic carbon content as affected by an experimental forest fire in the greater Xingan Mountains. J. Soil Water Conserv. 2012, 26, 195–200. [Google Scholar]
- Wei, X.; Blanco, J.A.; Jiang, H.; Kimmins, J.P.H. Effects of nitrogen deposition on carbon sequestration in Chinese fir forest ecosystems. Sci. Total Environ. 2012, 416, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Fernandez, I.J.; Rustad, L.E.; Norton, S.A. Effects of nitrogen enrichment, wildfire, and harvesting on forest soil carbon and nitrogen. Soil Sci. Soc. Am. J. 2001, 65, 1248–1255. [Google Scholar] [CrossRef]
- Islam, K.R.; Weil, R.R. Land use effects on soil quality in a tropical forest ecosystem of Bangladesh. Agric. Ecosyst. Environ. 2000, 79, 9–16. [Google Scholar] [CrossRef]
- Kalbitz, K.; Solinger, S.; Park, J.H.; Michalzik, B.; Matzner, E. Controls on the dynamics of dissolved organic matter in soils: A review. Soil Sci. 2000, 165, 277–304. [Google Scholar] [CrossRef]
- Zhou, G.Y.; Morris, J.D.; Yan, J.H.; Yu, Z.Y.; Peng, S.L. Hydrological impacts of reafforestation with eucalypts and indigenous species: A case study in southern China. For. Ecol. Manag. 2002, 167, 209–222. [Google Scholar] [CrossRef]
- Li, X.; Ye, D.; Liang, H.; Zhu, H.; Qin, L.; Zhu, Y.; Wen, Y. Effects of successive rotation regimes on carbon stocks in eucalyptus plantations in subtropical China measured over a full rotation. PLoS ONE 2015, 10, e0132858. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Tang, X.; Fan, S.; Zhao, J.; Peng, C. Changes in soil carbon and nitrogen stocks followed the conversion from secondary forest to Chinese fir and Moso bamboo plantations. Catena 2015, 133, 455–460. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Community | Forest Age (Years) | Area (ha) | Plot Series | Number of Replicates | |
|---|---|---|---|---|---|---|
| Natural secondary forest | SCF | Pinus massoniana Lamb. + Adina pilulifera (Lam.) Franch. ex Drake | 20–25 | 12,747 | a | 3 |
| Cunninghamia lanceolata (Lamb.) Hook. + Psychotria rubra (Lour.) Poir. | 20–25 | b | 3 | |||
| SCBF | Pinus massoniana + Castanea henryi (Skan) Rehd. & E. H. Wils. + Schima superb Gardner & Champ. | 25–30 | 23,962 | a | 3 | |
| Pinus massoniana Lamb. + Castanopsis fissa (Champ. Ex Benth.) Rehd. & E. H. Wils. + Schima superb | 30–35 | b | 3 | |||
| Cunninghamia lanceolata + Schima superb + Psychotria rubra (Lour.) Poir. | 30–35 | c | 3 | |||
| SBF | Castanopsis fissa + Castanopsis carlesii (Hemsl.) Hayata + Schima superb | 30–35 | 102,643 | a | 3 | |
| Cryptocarya concinna Hance + Castanea henryi + Schima superb | 40–50 | b | 3 | |||
| Ficus variolosa Lindl. Ex Benth. + Castanopsis faberi Hance + Lithocarpus glaber (Thunb.) Nakai | 45–55 | c | 3 | |||
| Castanopsis carlesii + Cinnamomum porrectum (Roxb.) Kosterm + Machilus chinensis (Benth.) Hemsl. | 45–55 | d | 3 | |||
| Elaeocarpus sylvestris (Lour.) Poir. + Sloanea sinensis (Hance) Hemsl. + Pithecellobium clypearia (Jack) Benth. | 45–55 | e | 3 | |||
| Planted forest | TF | Pinus massoniana | 5–15 | 2319 | a | 3 |
| Cunninghamia lanceolata | 5–15 | 1371 | b | 3 | ||
| Eucalyptus urophylla S. T. Blakely | 3–7 | 3358 | c | 3 | ||
| Phyllostachys heterocycla (Carr.) Mitford’ Pubescens’ | — | 3079 | d | 3 | ||
| NF | Litchi chinensis Sonn. | 6–30 | 16,543 | a | 9 | |
| Vatica mangachapoi Blanco | 5–10 | 492 | b | 3 | ||
| Citrus reticulata Blanco | 5–10 | 580 | c | 3 | ||
| Tree Species | Regression Equation (B1, D2, H3) and the Statistics |
|---|---|
| Pinus massoniana | BA = 0.0245(D2H)1.0209, R2 = 0.97; BR = 0.0126(D2H)0.9024, R2 = 0.97 [43] |
| Cunninghamia lanceolata | BA = 0.093(D2H)0.8030, R2 = 0.98; BR = 0.0073D2.3125, R2 = 0.99 [42] |
| Eucalyptus urophylla | BA = 0.1691(D2H)0.7472, R2 = 0.95; BR = 0.1723(D2H)0.5630, R2 = 0.90 [43] |
| Phyllostachys heterocycla | BA = 0.6439D1.5373, R2 = 0.91; BR1 = 0.3404D1.1899, R2 = 0.97, BR2 = 0.3087D1.2892, R2 = 0.80, BR = BR1 + BR2, [41] |
| Cryptocarya concinna, Cinnamomum porrectum, Machilus chinensis | BS = 0.0440(D2H)0.9169, R2 = 0.99; BP = 0.023(D2H)0.7115, R2 = 0.95; BB = 0.0104(D2H)0.9994, R2 = 0.92; BL = 0.0188(D2H)0.8024, R2 = 0.91; BA = BS + BP + BB + BL; BR = 0.0197(D2H)0.8963, R2 = 0.99 [41] |
| Castanea henryi, Castanopsis fissa, Castanopsis carlesii, Castanopsis faberi, Lithocarpus glaber | BA = 0.120(D2H)0.8509, R2 = 0.99; BR = 0.098 + 0.0176(D2H), R2 = 0.99 [41] |
| Others | BA = (D2H)/(0.0014(D2H) + 15.990), R2 = 0.99; BR = 0.376 + 0.0162(D2H), R2 = 0.99 [41] |
| Forest Type | Carbon Concentration (%) | |
|---|---|---|
| Understory | Litter | |
| SCF | 45.07 ± 0.84 | 46.71 ± 0.19 |
| SCBF | 42.54 ± 0.51 | 43.95 ± 0.74 |
| SBF | 41.37 ± 0.56 | 42.40 ± 0.40 |
| TF | 40.41 ± 2.02 | 42.90 ± 2.78 |
| NF | 38.90 ± 1.78 | 40.53 ± 0.62 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Xia, B.; Lin, J. A Basin-Scale Estimation of Carbon Stocks of a Forest Ecosystem Characterized by Spatial Distribution and Contributive Features in the Liuxihe River Basin of Pearl River Delta. Forests 2016, 7, 299. https://doi.org/10.3390/f7120299
Zhang C, Xia B, Lin J. A Basin-Scale Estimation of Carbon Stocks of a Forest Ecosystem Characterized by Spatial Distribution and Contributive Features in the Liuxihe River Basin of Pearl River Delta. Forests. 2016; 7(12):299. https://doi.org/10.3390/f7120299
Chicago/Turabian StyleZhang, Chao, Beicheng Xia, and Junyu Lin. 2016. "A Basin-Scale Estimation of Carbon Stocks of a Forest Ecosystem Characterized by Spatial Distribution and Contributive Features in the Liuxihe River Basin of Pearl River Delta" Forests 7, no. 12: 299. https://doi.org/10.3390/f7120299
APA StyleZhang, C., Xia, B., & Lin, J. (2016). A Basin-Scale Estimation of Carbon Stocks of a Forest Ecosystem Characterized by Spatial Distribution and Contributive Features in the Liuxihe River Basin of Pearl River Delta. Forests, 7(12), 299. https://doi.org/10.3390/f7120299