
Anthropogenic Disturbances Create a New Vegetation Toposequence in the Gatineau River Valley, Quebec


Abstract
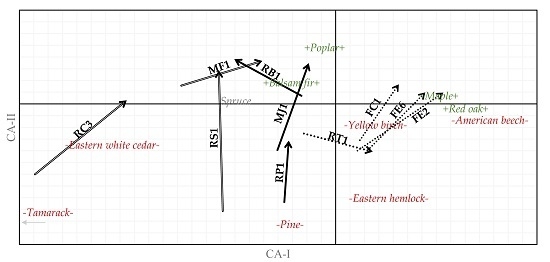
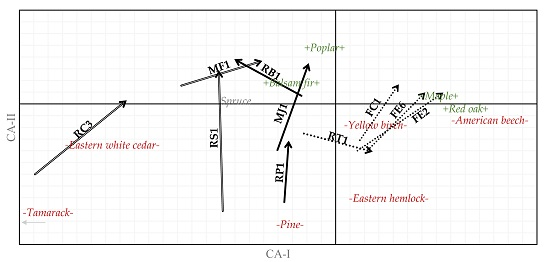
:
1. Introduction
2. Materials and Methods
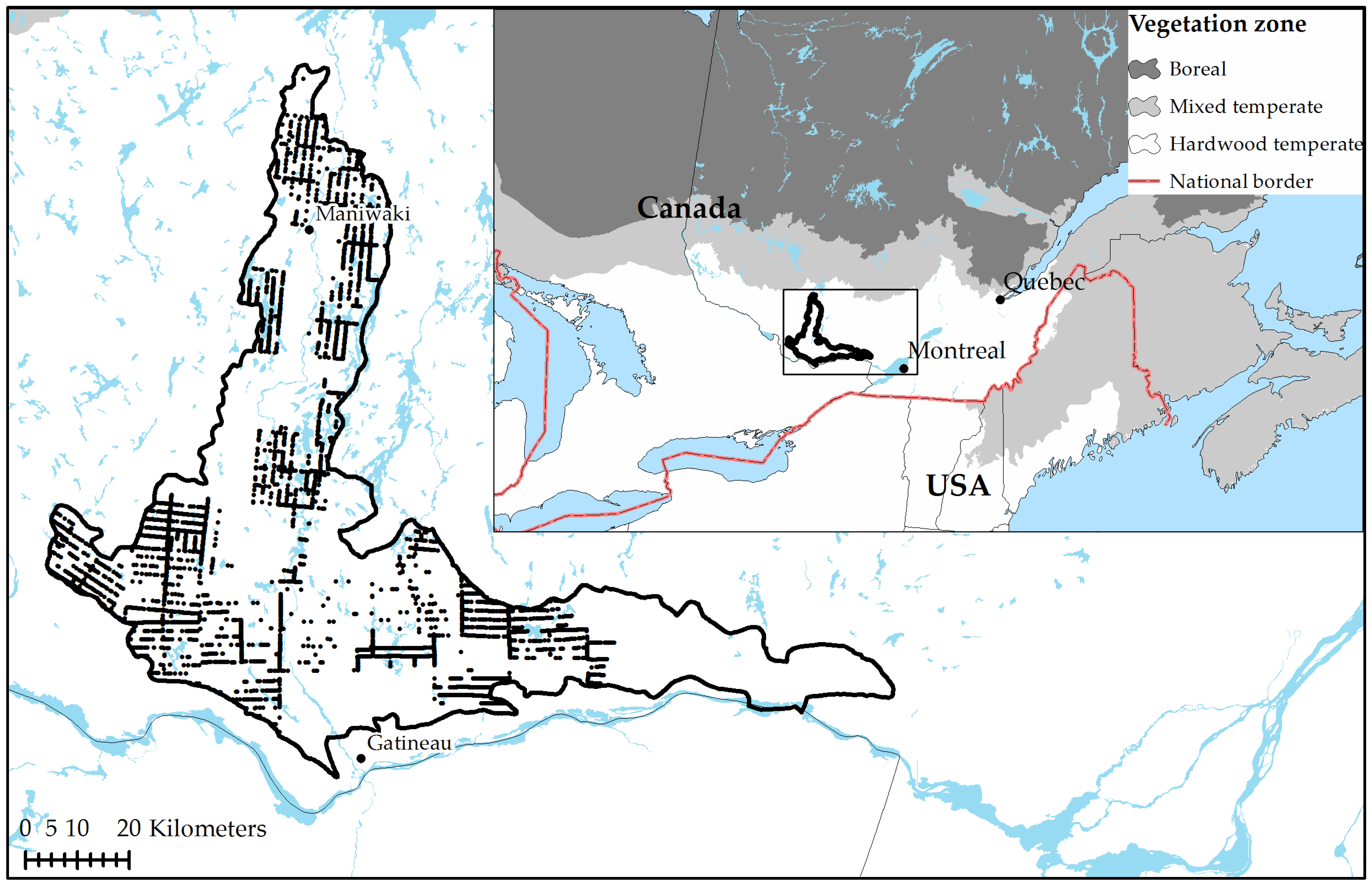
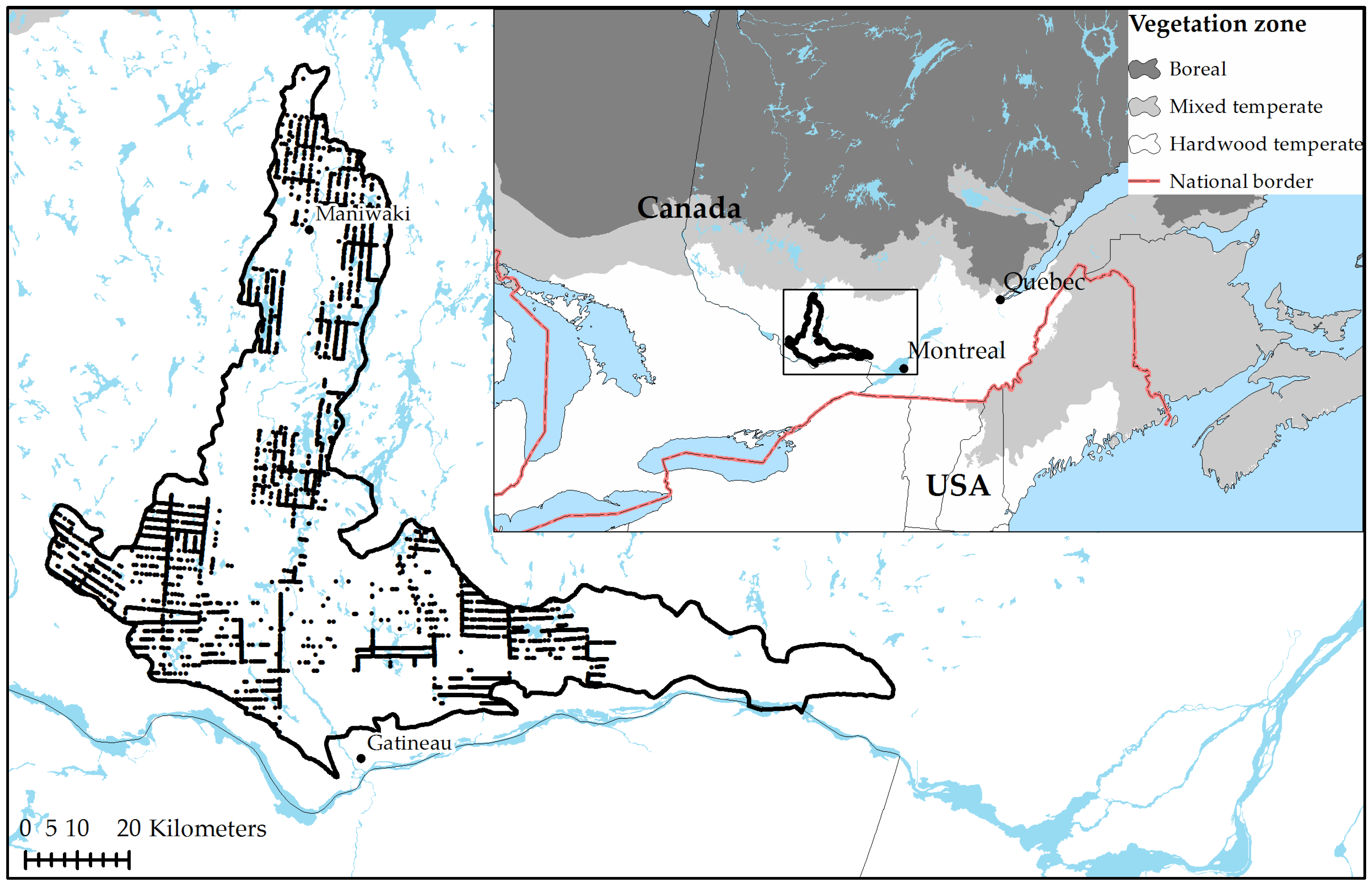
2.1. Study Area
2.2. Preindustrial Forest Composition
2.3. Current Forest Composition
2.4. Evolution of the Forest Composition of Potential Vegetation Types
3. Results
3.1. Cover Type
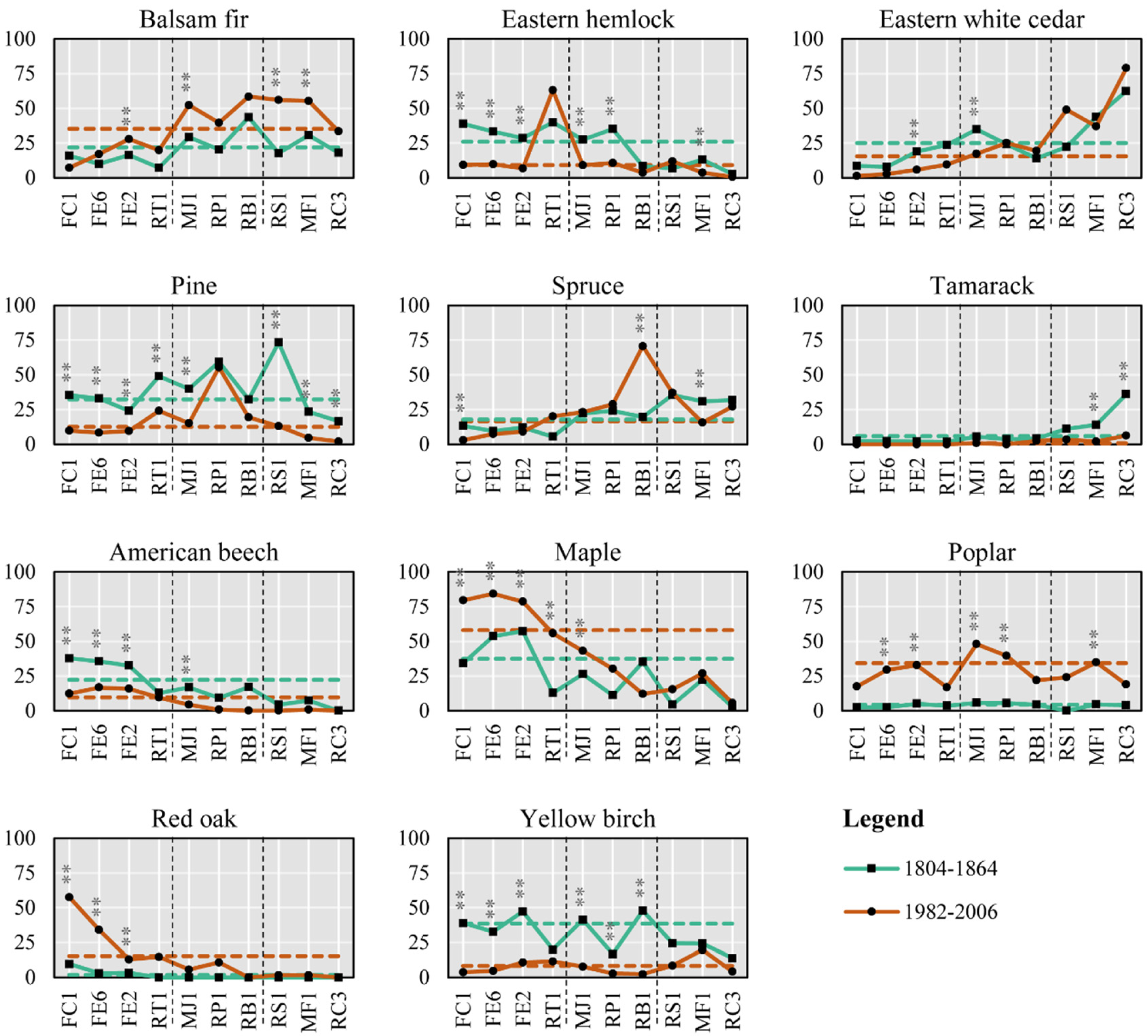
3.2. Tree Species
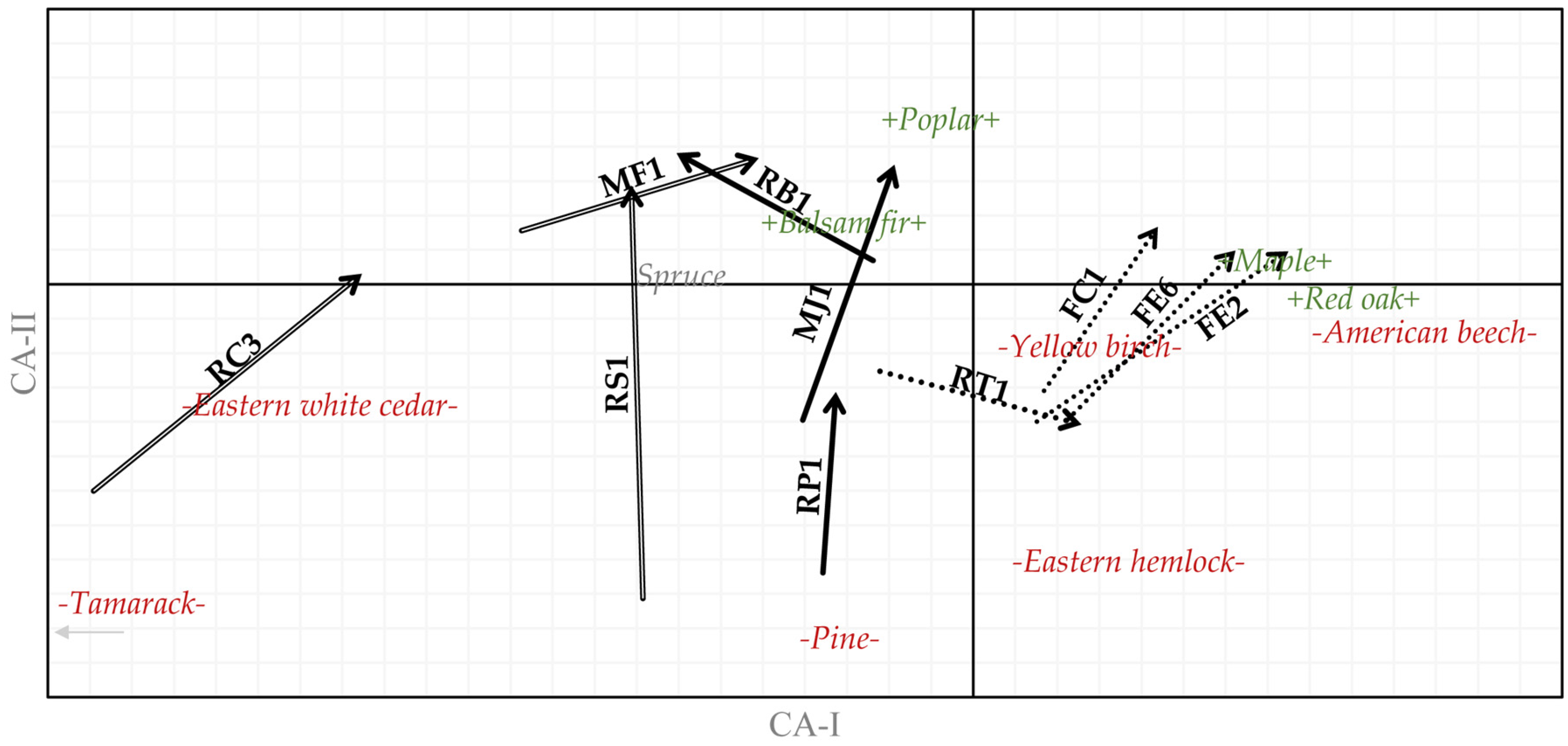
3.3. Forest Types
4. Discussion
4.1. Loss of Targeted Species
4.1.1. Pine: The Great Decline
4.1.2. Hemlock Has Been Replaced by Maple
4.2. Modification of Internal Stand Composition
Loss of the Two Main Companion Tree Species to Maple
4.3. Novel Ecosystems
4.3.1. Old Fields
4.3.2. Red Oak Stands
4.3.3. Early Successional Stands: The Case of Poplar
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preindustrial Forest Type # | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| n | 282 | 373 | 447 | 360 | 298 | 207 | 101 | 110 | 70 | 252 |
| Acer spp. | 21.0 | 63.3 | 3.1 | 0.8 | 3.0 | 8.5 | 3.8 | 2.1 | 0.0 | 6.9 |
| Betula alleghaniensis | 7.9 | 11.9 | 3.6 | 2.7 | 7.4 | 59.8 | 5.1 | 9.5 | 1.5 | 5.9 |
| Betula papyrifera | 0.4 | 0.1 | 0.5 | 0.3 | 1.0 | 0.0 | 0.3 | 0.8 | 0.6 | 8.4 |
| Fagus grandifolia | 63.8 | 10.0 | 0.6 | 0.1 | 2.8 | 2.7 | 0.4 | 2.2 | 0.0 | 0.7 |
| Fraxinus spp. | 0.0 | 0.3 | 0.3 | 2.2 | 0.1 | 0.5 | 1.0 | 0.8 | 0.3 | 29.6 |
| Ostrya virginiana | 0.6 | 0.5 | 0.1 | 0.0 | 0.0 | 0.4 | 0.0 | 0.1 | 0.0 | 1.5 |
| Populus spp. | 0.0 | 0.5 | 0.8 | 0.1 | 0.0 | 1.1 | 0.0 | 1.6 | 0.2 | 10.7 |
| Quercus spp. | 0.1 | 0.5 | 0.1 | 0.0 | 0.2 | 0.2 | 0.0 | 0.1 | 0.0 | 5.4 |
| Tilia americana | 0.7 | 4.4 | 0.2 | 0.2 | 0.1 | 2.0 | 0.0 | 0.2 | 0.0 | 4.3 |
| Ulmus spp. | 0.1 | 0.9 | 0.1 | 0.3 | 0.0 | 0.9 | 0.2 | 0.6 | 0.0 | 12.8 |
| Abies balsamea | 0.4 | 1.8 | 2.4 | 3.6 | 1.7 | 9.4 | 7.3 | 63.5 | 1.2 | 3.0 |
| Larix laricina | 0.0 | 0.0 | 0.4 | 2.1 | 0.0 | 0.3 | 3.9 | 0.5 | 84.2 | 0.2 |
| Picea spp. | 0.0 | 0.5 | 3.4 | 4.5 | 1.3 | 5.9 | 65.8 | 9.3 | 3.5 | 1.6 |
| Pinus spp. | 0.8 | 1.0 | 77.7 | 2.9 | 8.2 | 4.7 | 5.0 | 4.6 | 0.5 | 2.2 |
| Thuja occidentalis | 0.0 | 1.7 | 2.4 | 77.3 | 4.0 | 1.3 | 3.9 | 2.7 | 4.8 | 3.7 |
| Tsuga canadensis | 4.1 | 2.4 | 4.3 | 2.6 | 70.3 | 2.5 | 0.3 | 0.5 | 0.0 | 1.2 |
| Other | 0.0 | 0.1 | 0.0 | 0.3 | 0.0 | 0.0 | 2.9 | 0.9 | 3.2 | 1.9 |
| Present-Day Forest Type # | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| n | 1242 | 676 | 365 | 458 | 278 | 223 | 178 | 175 | 562 |
| Acer spp. | 69.3 | 9.0 | 1.8 | 6.8 | 15.7 | 5.9 | 2.3 | 13.4 | 11.3 |
| Betula alleghaniensis | 1.8 | 0.3 | 1.1 | 1.1 | 0.0 | 0.3 | 0.2 | 3.8 | 9.1 |
| Betula papyrifera | 1.7 | 3.0 | 1.5 | 3.0 | 1.3 | 1.8 | 2.8 | 2.3 | 11.5 |
| Fagus grandifolia | 3.9 | 0.5 | 0.0 | 0.0 | 1.1 | 0.2 | 0.0 | 3.3 | 11.7 |
| Fraxinus spp. | 1.4 | 0.8 | 4.0 | 1.5 | 0.5 | 0.3 | 1.0 | 0.5 | 13.3 |
| Ostrya virginiana | 2.5 | 0.3 | 0.1 | 0.4 | 4.0 | 0.7 | 0.7 | 0.4 | 6.9 |
| Populus spp. | 3.7 | 67.2 | 4.5 | 7.9 | 2.0 | 5.1 | 5.6 | 1.1 | 3.8 |
| Quercus spp. | 3.2 | 1.3 | 0.0 | 0.7 | 67.1 | 3.5 | 0.5 | 2.4 | 1.1 |
| Tilia americana | 3.9 | 1.0 | 0.6 | 1.1 | 1.0 | 0.3 | 0.3 | 0.6 | 10.1 |
| Ulmus spp. | 0.2 | 0.2 | 0.0 | 0.2 | 0.2 | 0.0 | 0.5 | 0.2 | 1.9 |
| Abies balsamea | 3.9 | 8.6 | 8.7 | 64.8 | 1.4 | 5.1 | 8.2 | 2.4 | 4.9 |
| Larix laricina | 0.0 | 0.0 | 0.5 | 0.2 | 0.0 | 0.1 | 0.3 | 0.0 | 0.9 |
| Picea spp. | 0.9 | 2.9 | 2.7 | 5.1 | 1.1 | 5.0 | 70.1 | 1.4 | 1.8 |
| Pinus spp. | 0.9 | 2.1 | 0.8 | 2.0 | 3.4 | 67.1 | 3.3 | 1.3 | 0.6 |
| Thuja occidentalis | 0.4 | 1.6 | 73.0 | 3.5 | 0.0 | 2.5 | 2.6 | 0.8 | 2.3 |
| Tsuga canadensis | 1.8 | 0.4 | 0.3 | 0.8 | 1.0 | 1.5 | 0.3 | 66.1 | 0.7 |
| Other | 0.5 | 0.8 | 0.4 | 0.8 | 0.1 | 0.5 | 1.2 | 0.1 | 7.9 |
References
- Fuller, J.L.; Foster, D.R.; McLachlan, J.S.; Drake, N. Impact of human activity on regional forest composition and dynamics in central New England. Ecosystems 1998, 1, 76–95. [Google Scholar] [CrossRef]
- Thompson, J.R.; Carpenter, D.N.; Cogbill, C.V.; Foster, D.R. Four centuries of change in northeastern United States forests. PLoS ONE 2013, 8, e72540. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, G.J.; Abrams, M.D. Is climate an important driver of post-European vegetation change in the Eastern United States? Glob. Chang. Biol. 2015, 21, 314–334. [Google Scholar] [CrossRef] [PubMed]
- Simard, H.; Bouchard, A. The precolonial 19th century forest of the Upper St. Lawrence Region of Quebec: A record of its exploitation and transformation through notary deeds of wood sales. Can. J. For. Res. 1996, 26, 1670–1676. [Google Scholar] [CrossRef]
- Thompson, I.D.; Simard, J.H.; Titman, R.D. Historical changes in white pine (Pinus strobus L.) density in Algonquin Park, Ontario, during the 19th century. Nat. Area J. 2006, 26, 61–71. [Google Scholar] [CrossRef]
- Marchand, N.; Filion, L. A dendroecological analysis of eastern hemlock and white pine in relation to logging in La Mauricie National Park (Québec, Canada). For. Chron. 2014, 90, 351–360. [Google Scholar] [CrossRef]
- Dupuis, S.; Arseneault, D.; Sirois, L. Change from pre-settlement to present-day forest composition reconstructed from early land survey records in eastern Québec, Canada. J. Veg. Sci. 2011, 22, 564–575. [Google Scholar] [CrossRef]
- Danneyrolles, V.; Arseneault, D.; Bergeron, Y. Pre-industrial landscape composition patterns and post-industrial changes at the temperate–boreal forest interface in western Quebec, Canada. J. Veg. Sci. 2016, 27, 470–481. [Google Scholar] [CrossRef]
- Gauthier, S.; Vaillancourt, M.-A.; Kneeshaw, D.D.; Drapeau, P.; de Grandpré, L.; Claveau, Y.; Paré, D. Forest ecosystem management: Origins and foundations. In Ecosystem Management in the Boreal Forest; Gauthier, S., Vaillancourt, M.-A., Leduc, A., de Grandpré, L., Kneeshaw, D.D., Morin, H., Drapeau, P., Bergeron, Y., Eds.; Les Presses de l’Université du Québec: Quebec, QC, Canada, 2009; pp. 13–38. [Google Scholar]
- Boucher, Y.; Bouchard, M.; Grondin, P.; Tardif, P. Le Registre des états de Référence: Intégration des Connaissances sur la Structure, la Composition et la Dynamique des Paysages Forestiers Naturels du Québec Méridional; Gouvernement du Québec, Direction de la Recherche Forestière: Quebec, QC, Canada, 2011; pp. 1–14.
- Whitney, G.G. From Coastal Wilderness to Fruited Plain. A History of Environmental Change in Temperate North America from 1500 to the Present; Cambridge University Press: Cambridge, UK, 1994; pp. 1–38. [Google Scholar]
- Foster, D.R.; Motzkin, G.; Slater, B. Land-use history as long-term broad-scale disturbance: Regional forest dynamics in central New England. Ecosystems 1998, 1, 96–119. [Google Scholar] [CrossRef]
- Radeloff, V.C.; Mladenoff, D.J.; He, H.S.; Boyce, M.S. Forest landscape change in the northwestern Wisconsin Pine Barrens from pre-European settlement to the present. Can. J. For. Res. 1999, 29, 1649–1659. [Google Scholar] [CrossRef]
- Rhemtulla, J.M.; Mladenoff, D.J.; Clayton, M.K. Legacies of historical land use on regional forest composition and structure in Wisconsin, USA (mid-1800s–1930s–2000s). Ecol. Appl. 2009, 19, 1061–1078. [Google Scholar] [CrossRef] [PubMed]
- Siccama, T.G. Presettlement and present forest vegetation in northern Vermont with special reference to Chittenden County. Am. Midl. Nat. 1971, 85, 153–172. [Google Scholar] [CrossRef]
- Whitney, G.G. An ecological history of the great lakes forest of Michigan. J. Ecol. 1987, 75, 667–684. [Google Scholar] [CrossRef]
- Abrams, M.D.; Ruffner, C.M. Physiographic analysis of witness-tree distribution (1765–1798) and present forest cover through north central Pennsylvania. Can. J. For. Res. 1995, 25, 659–668. [Google Scholar] [CrossRef]
- Zhang, Q.; Pregitzer, K.S.; Reed, D.D. Historical changes in the forests of the Luce District of the Upper Peninsula of Michigan. Am. Midl. Nat. 2000, 143, 94–110. [Google Scholar] [CrossRef]
- Leahy, M.J.; Pregitzer, K.S. A comparaison of presettlement and present-day forests in northeastern Lower Michigan. Am. Midl. Nat. 2003, 149, 71–89. [Google Scholar] [CrossRef]
- Vadeboncoeur, M.A.; Hamburg, S.P.; Cogbill, C.V.; Sugimura, W.Y. A comparison of presettlement and modern forest composition along an elevation gradient in central New Hampshire. Can. J. For. Res. 2012, 42, 190–202. [Google Scholar] [CrossRef]
- Leadbitter, P.; Euler, D.; Naylor, B. A comparison of historical and current forest cover in selected areas of the Great Lakes—St. Lawrence forest of central Ontario. For. Chron. 2002, 78, 522–529. [Google Scholar] [CrossRef]
- Brisson, J.; Bouchard, A. In the past two centuries, human activities have caused major changes in the tree species composition of southern Québec, Canada. Ecoscience 2003, 10, 235–246. [Google Scholar]
- Aubé, M. The pre-European settlement forest composition of the Miramichi River watershed, New Brunswick, as reconstructed using witness trees from original land surveys. Can. J. For. Res. 2008, 38, 1159–1183. [Google Scholar] [CrossRef]
- Pinto, F.; Rornaniuk, S.; Ferguson, M. Changes to preindustrial forest tree composition in central and northeastern Ontario, Canada. Can. J. For. Res. 2008, 38, 1842–1854. [Google Scholar] [CrossRef]
- Zelazny, V.F.; Veen, H.; Colpitts, M.C. Potential Forests of the Fundy Model Forest; New Brunswick Department of Natural Resources and Energy, Forest Management Branch: Fredericton, NB, Canada, 1997.
- Saucier, J.-P.; Robitaille, A.; Grondin, P. Cadre bioclimatique du Québec. In Manuel de Foresterie: Écologie Forestière, 2nd ed.; Saucier, J.-P., Grondin, P., Robitaille, A., Gosselin, J., Morneau, C., Richard, P.J.H., Brisson, J., Sirois, L., Leduc, A., Morin, H., et al., Eds.; Éditions MultiMondes: Quebec, QC, Canada, 2009; pp. 186–206. [Google Scholar]
- Grondin, P.; Gauthier, S.; Borcard, D.; Bergeron, Y.; Noël, J. A new approach to ecological land classification for the Canadian boreal forest that integrates disturbances. Landsc. Ecol. 2014, 29, 1–16. [Google Scholar] [CrossRef]
- Ferree, C.; Anderson, M.G. A Map of Terrestrial Habitats of the Northeastern United States: Methods and Approach; The Nature Conservancy, Eastern Conservation Science, Eastern Regional Office: Boston, MA, USA, 2013; pp. 1–45. [Google Scholar]
- Hobbs, R.J.; Arico, S.; Aronson, J.; Baron, J.S.; Bridgewater, P.; Cramer, V.A.; Epstein, P.R.; Ewel, J.J.; Klink, C.A.; Lugo, A.E.; et al. Novel ecosystems: Theoretical and management aspects of the new ecological world order. Glob. Ecol. Biogeogr. 2006, 15, 1–7. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.; Hall, C.M.; Bridgewater, P.; Chapin, F.S.; Ellis, E.C.; Ewel, J.J.; Hallett, L.M.; Harris, J.; Hulvey, K.B.; et al. Managing the whole landscape: Historical, hybrid, and novel ecosystems. Front. Ecol. Environ. 2014, 12, 557–564. [Google Scholar] [CrossRef]
- Jackson, S.T.; Hobbs, R.J. Ecological restoration in the light of ecological history. Science 2009, 325, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, J.; Doyon, F. Description des domaines bioclimatiques: Domaine de l’érablière à tilleul. In Manuel de Foresterie: Écologie Forestière, 2nd ed.; Saucier, J.-P., Grondin, P., Robitaille, A., Gosselin, J., Morneau, C., Richard, P.J.H., Brisson, J., Sirois, L., Leduc, A., Morin, H., et al., Eds.; Éditions MultiMondes: Quebec, QC, Canada, 2009; pp. 215–227. [Google Scholar]
- Dyke, A.S. An outline of North American deglaciation with emphasis on central and northern Canada. In Quaternary Glaciations-Extent and Chronology—Part II: North America; Ehlers, J., Gibbard, P.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 373–424. [Google Scholar]
- Brandt, J.P. The extent of the North American boreal zone. Environ. Rev. 2009, 17, 101–161. [Google Scholar] [CrossRef]
- Gosselin, J. Guide de Reconnaissance des Types Écologiques: Région Écologique 2a, Collines de la Basse Gatineau; Gouvernement du Québec, Direction des Inventaires Forestiers: Quebec, QC, Canada, 2004; pp. 48–53.
- Greffe de l’Arpenteur Général du Québec. Available online: https://appli.mern.gouv.qc.ca/Gagq (accessed on 22 August 2016).
- Fournier, R. Plantes Vasculaires du Nouveau-Brunswick: Dicotylédones—Synonymes; Université de Moncton, Campus d’Edmundston, Faculté de foresterie: Edmunston, NB, Canada, 2006; pp. 2–141. [Google Scholar]
- Fournier, R. Plantes Vasculaires du Nouveau-Brunswick: Ptéridophytes et Gymnospermes—Synonymes; Université de Moncton, Campus d’Edmundston, Faculté de foresterie: Edmunston, NB, Canada, 2006; pp. 2–97. [Google Scholar]
- Jackson, S.M.; Pinto, F.; Malcolm, J.R.; Wilson, E.R. A comparison of pre-European settlement (1857) and current (1981–1995) forest composition in central Ontario. Can. J. For. Res. 2000, 30, 605–612. [Google Scholar] [CrossRef]
- Seischab, F.K. Presettlement forests of the Phelps and Gorham purchase in Western New York. Bull. Torrey Bot. Club 1990, 117, 27–38. [Google Scholar] [CrossRef]
- Scull, P.R.; Richardson, J.L. A method to use ranked timber observations to perform forest composition reconstruction from land survey data. Am. Midl. Nat. 2007, 158, 446–460. [Google Scholar] [CrossRef]
- Terrail, R.; Arseneault, D.; Fortin, M.-J.; Dupuis, S.; Boucher, Y. An early forest inventory indicates high accuracy of forest composition data in pre-settlement land survey records. J. Veg. Sci. 2014, 25, 691–702. [Google Scholar] [CrossRef]
- Frontier, S. Decrease of eigenvalues in principal component analysis—Comparison with broken stick model. J. Exp. Mar. Biol. Ecol. 1976, 25, 67–75. [Google Scholar] [CrossRef]
- Les Placettes-Échantillons Temporaires. Available online: https://www.mffp.gouv.qc.ca/forets/inventaire/inventaire-placettes.jsp (accessed on 10 October 2016).
- Oksanen, J.; Blanchet, G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. vegan: Community Ecology Package. R Package Version 1.17-4. 2010. Available online: https://CRAN.R-project.org/package=vegan (accessed on 19 August 2016).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010. [Google Scholar]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Wang, Y.-C. Spatial patterns and vegetation-site relationships of the presettlement forests in western New York, USA. J. Biogeogr. 2007, 34, 500–513. [Google Scholar] [CrossRef]
- Palik, B.J.; Pregitzer, K.S. A comparison of presettlement and present-day forests on two bigtooth aspen-dominated landscapes in northern Lower Michigan. Am. Midl. Nat. 1992, 127, 327–338. [Google Scholar] [CrossRef]
- Cowell, C.M. Historical change in vegetation and disturbance on the Georgia piedmont. Am. Mild. Nat. 1998, 140, 78–89. [Google Scholar] [CrossRef]
- Friedman, S.K.; Reich, P.B. Regional legacies of logging: Departure from presettlement forest conditions in northern Minnesota. Ecol. Appl. 2005, 15, 726–744. [Google Scholar] [CrossRef]
- Mauri Ortuno, E. Modélisation de la Distribution Précoloniale du Pin Blanc en Moyenne-Mauricie à partir des Carnets d’Arpentage. Master’s Thesis, Université Laval, Quebec, QC, Canada, 2010. [Google Scholar]
- Mauri Ortuno, E.; Doyon, F. Estimation de la Distribution des Essences Forestières au 19e Siècle dans l’Outaouais à l’aide des Carnets d’Arpentage des Limites des Concessions Forestières; Institut Québécois d’Aménagement de la Forêt Feuillue (IQAFF): Ripon, QC, Canada, 2010. [Google Scholar]
- Sands, B.A.; Abrams, M.D. A 183-year history of fire and recent fire suppression impacts in select pine and oak forest stands of the Menominee Indian Reservation, Wisconsin. Am. Midl. Nat. 2011, 166, 325–338. [Google Scholar] [CrossRef]
- Wendel, G.W.; Smith, H.C. Eastern white pine. In Silvics of North America—Volume 1: Conifers; Forest Service Agriculture Handbook; Burns, R.M., Honkala, B.H., Eds.; United States Department of Agriculture: Washington, DC, USA, 1990; pp. 972–999. [Google Scholar]
- Fahey, T.J.; Reiners, W.A. Fire in the forests of Maine and New Hampshire. Bull. Torrey Bot. Club 1981, 108, 362–373. [Google Scholar] [CrossRef]
- Doyon, F.; Bouffard, D. Enjeux Écologiques de la Forêt Feuillue Tempérée Québécoise; Institut Québécois d’Aménagement de la Forêt Feuillue (IQAFF): Ripon, QC, Canada, 2009. [Google Scholar]
- Raymond, P.; Munson, A.D.; Ruel, J.C.; Nolet, P. Group and single-tree selection cutting in mixed tolerant hardwood-white pine stands: Early establishment dynamics of white pine and associated species. For. Chron. 2003, 79, 1093–1106. [Google Scholar] [CrossRef]
- Quenneville, R.; Thériault, M. La restauration des écosystèmes de pin blanc (Pinus strobus): Un enjeu majeur pour le parc national de la Mauricie. Nat. Can. 2001, 125, 39–42. [Google Scholar]
- White, M.A.; Mladenoff, D.J. Old-growth forest landscape transitions from pre-European settlement to present. Landsc. Ecol. 1994, 9, 191–205. [Google Scholar] [CrossRef]
- Suffling, R.; Evans, M.; Perera, A. Presettlement forest in southern Ontario: Ecosystems measured through a cultural prism. For. Chron. 2003, 79, 485–501. [Google Scholar] [CrossRef]
- Mladenoff, D.J.; Stearns, F. Eastern hemlock regeneration and deer browsing in the Great Lakes region: A re-examination and model simulation. Conserv. Biol. 1993, 7, 889–900. [Google Scholar] [CrossRef]
- Payette, S.; Pilon, V.; Couillard, P.-L.; Frégeau, M. Holocene dynamics of an eastern hemlock (Tsuga canadensis) forest site at the northern range of the species limit. Holocene 2015, 25, 1246–1256. [Google Scholar] [CrossRef]
- Brisson, J.; Bergeron, Y.; Bouchard, A. Les successions secondaires sur sites mésiques dans le Haut-Saint-Laurent, Québec, Canada. Can. J. Bot. 1988, 66, 1192–1203. [Google Scholar] [CrossRef]
- Réserve Écologique de la Forêt-la-Blanche. Available online: http://www.mddelcc.gouv.qc.ca/biodiversite/reserves/foret_lablanche/res_69.htm (accessed on 19 August 2016).
- Nolet, P.; Delagrange, S.; Bouffard, D.; Doyon, F.; Forget, E. The successional status of sugar maple (Acer saccharum), revisited. Ann. For. Sci. 2008, 65, 208. [Google Scholar] [CrossRef]
- Doyon, F.; Bouchard, A.; Gagnon, D. Tree productivity and successional status in Quebec northern hardwoods. Ecoscience 1998, 5, 222–231. [Google Scholar]
- Angers, V.A.; Messier, C.; Beaudet, M.; Leduc, A. Comparing composition and structure in old-growth and harvested northern hardwood stands in Quebec. For. Ecol. Manag. 2005, 217, 275–293. [Google Scholar] [CrossRef]
- Doyon, F.; Bouffard, D. Étude sur le Bois Mort, les Microsites et la Régénération après Chablis Catastrophiques dans les Érablières du Témiscamingue; Institut Québécois d’Aménagement de la Forêt Feuillue (IQAFF): Ripon, QC, Canada, 2008. [Google Scholar]
- Cogbill, C.V. Historical biogeography of American beech. In Beech Bark Disease, Proceedings of the Beech Bark Disease Symposium, Saranac Lake, NY, USA, 16–18 June 2004; USDA Forest Service: Newtown, PA, USA, 2005; pp. 16–24. [Google Scholar]
- Grandtner, M. La Végétation Forestière du Québec Méridional; Les Presses de l’Université Laval: Quebec, QC, Canada, 1966. [Google Scholar]
- Beaudet, M.; Messier, C. Growth and morphological responses of yellow birch, sugar maple, and beech seedlings growing under a natural light gradient. Can. J. For. Res. 1998, 28, 1007–1015. [Google Scholar] [CrossRef]
- Drinkwater, M.H. Field Spruce in Nova Scotia; Government of Canada, Department of Northern Affairs and National Resources, Forestry Branch: Ottawa, ON, Canada, 1957.
- Wang, Y.-C.; Kronenfeld, B.J.; Larsen, C.P.S. Spatial distribution of forest landscape change in western New York from presettlement to the present. Can. J. For. Res. 2009, 39, 76–88. [Google Scholar] [CrossRef]
- Abrams, M.D. Fire and the development of oak forests. BioScience 1992, 42, 346–353. [Google Scholar] [CrossRef]
- Terrail, R. Influence de la Colonisation sur les Transformations du Paysage Forestier depuis l’Époque Préindustrielle dans l’est du Québec (Canada). Ph.D. Thesis, Université du Québec à Rimouski, Rimouski, QC, Canada, 2013. [Google Scholar]
- Drever, C.R.; Messier, C.; Bergeron, Y.; Doyon, F. Fire and canopy species composition in the Great Lakes-St. Lawrence forest of Témiscamingue, Québec. For. Ecol. Manag. 2006, 231, 27–37. [Google Scholar] [CrossRef]
- Dey, D.C.; Guyette, R.P. Anthropogenic fire history and red oak forests in south-central Ontario. For. Chron. 2000, 76, 339–347. [Google Scholar] [CrossRef]
- Perala, D.A. Quaking aspen. In Silvics of North America—Volume 2: Hardwoods; Forest Service Agriculture Handbook; Burns, R.M., Honkala, B.H., Eds.; United States Department of Agriculture: Washington, DC, USA, 1990; pp. 1082–1115. [Google Scholar]
- Boucher, Y.; Grondin, P.; Auger, I. Land use history (1840–2005) and physiography as determinants of southern boreal forests. Landsc. Ecol. 2014, 29, 437–450. [Google Scholar] [CrossRef]
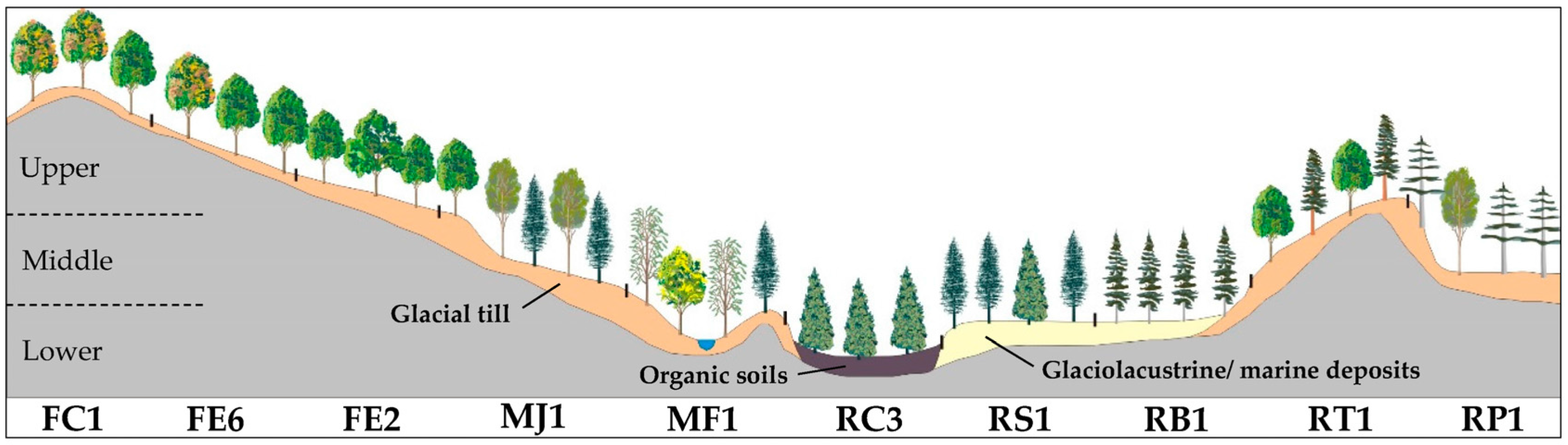

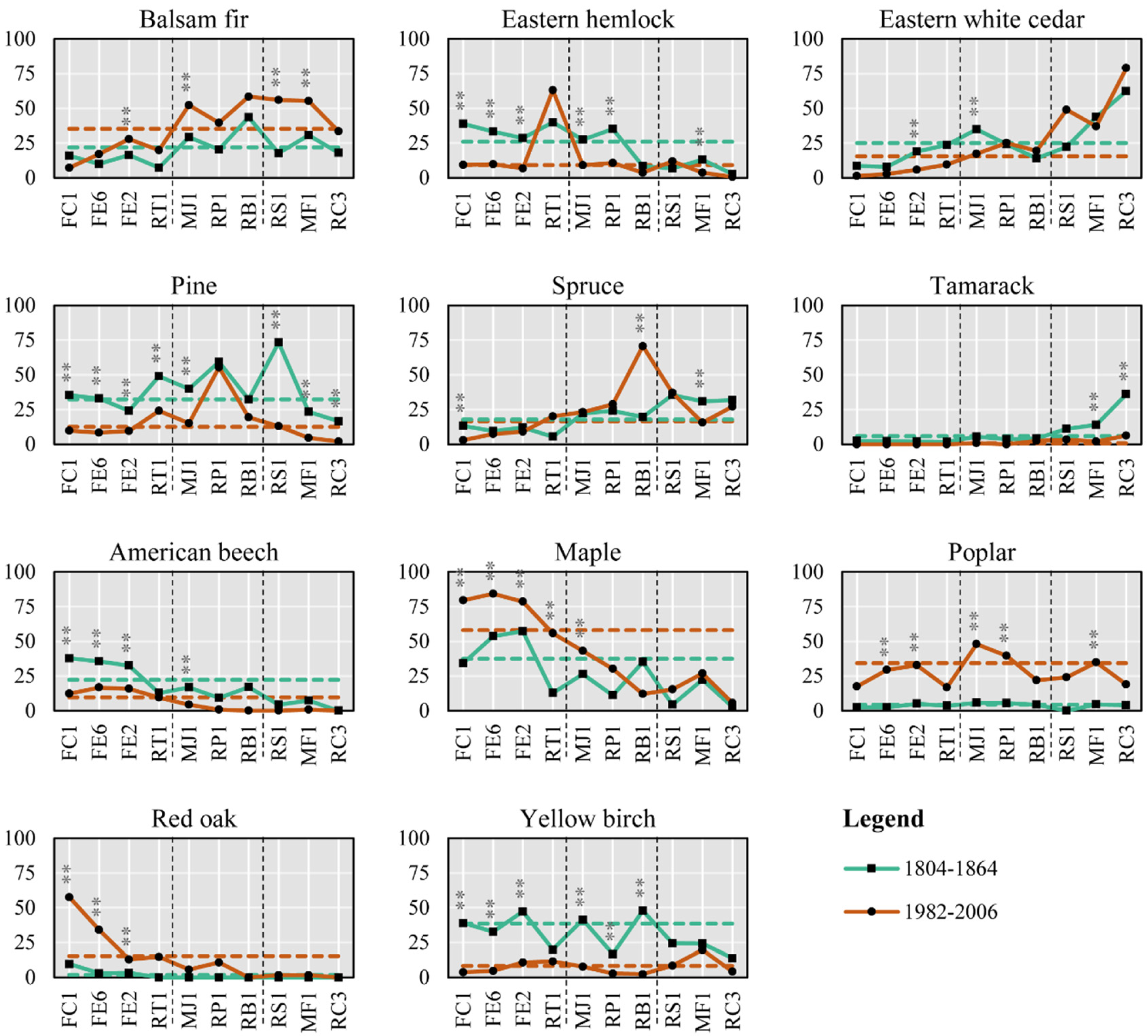
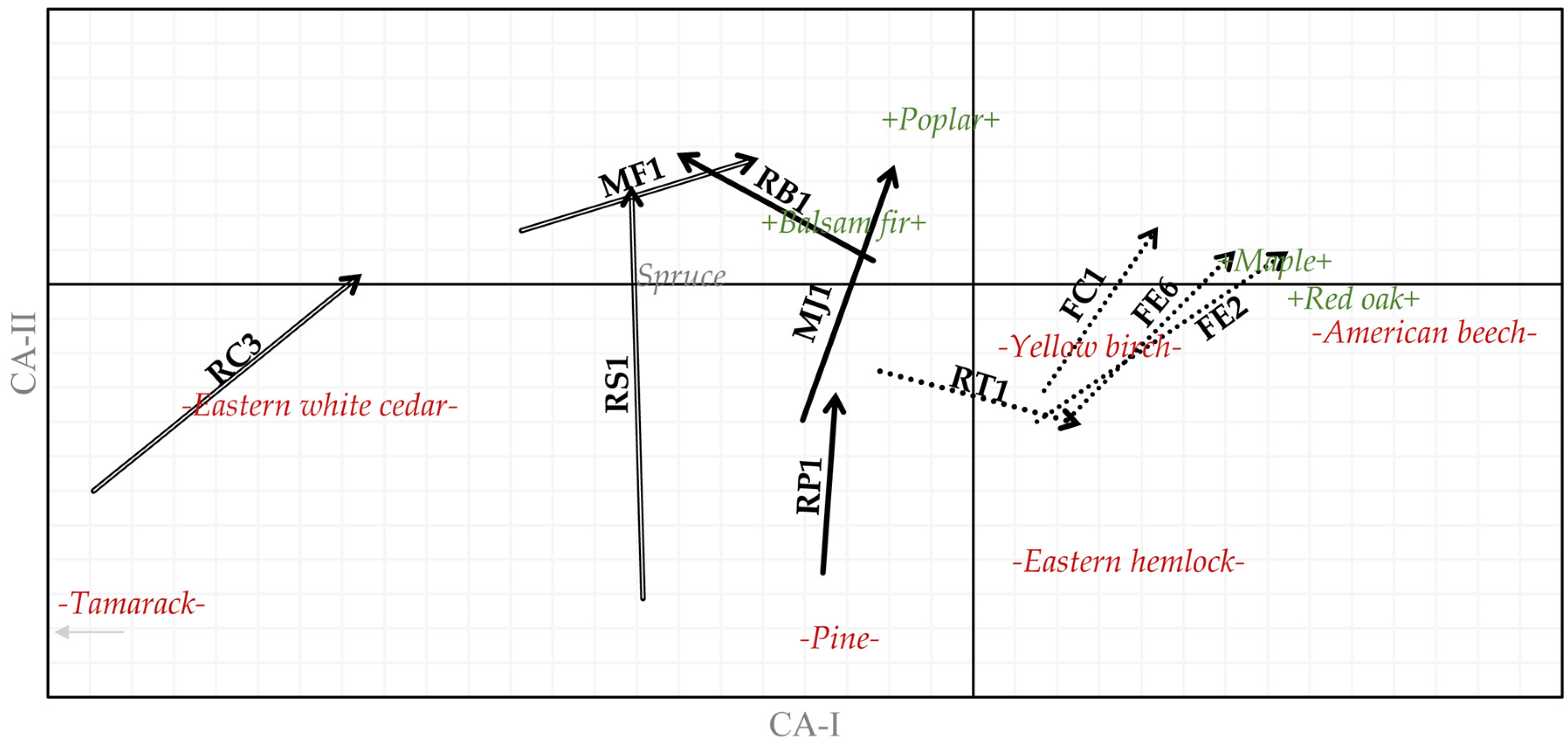
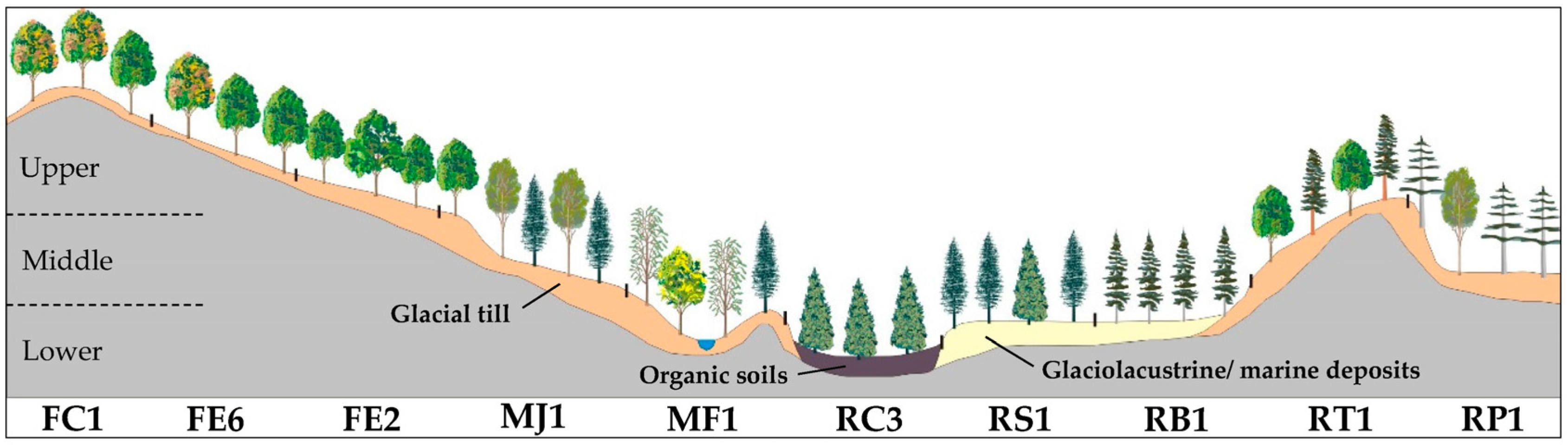
| Slope Position | Code | Potential Vegetation Types (PVT) | Mean Elevation | Mean Slope | Area |
|---|---|---|---|---|---|
| (m) | (%) | (%) | |||
| Upper | FC1 | Quercus rubra | 273 ± 68 | 21 ± 10 | 6.1 |
| FE6 | Acer saccharum–Quercus rubra | 247 ± 57 | 17 ± 8 | 15.7 | |
| FE2 | Acer saccharum–Tilia americana | 230 ± 57 | 13 ± 7 | 32.1 | |
| RT1 | Tsuga canadensis | 239 ± 59 | 21 ± 11 | 2.6 | |
| Middle | MJ1 | Betula alleghaniensis–Abies balsamea–Acer saccharum | 199 ± 46 | 8 ± 6 | 23.9 |
| RP1 | Pinus strobus–Pinus resinosa | 199 ± 41 | 13 ± 10 | 4.4 | |
| RB1 | Picea glauca–Thuja occidentalis (old fields) | 182 ± 29 | 6 ± 4 | 5.5 | |
| Lower | RS1 | Abies balsamea–Thuja occidentalis | 187 ± 30 | 7 ± 6 | 3.9 |
| MF1 | Fraxinus nigra–Abies balsamea | 185 ± 34 | 4 ± 3 | 2.2 | |
| RC3 | Thuja occidentalis–Abies balsamea | 195 ± 28 | 3 ± 3 | 2.5 | |
| Other PVTs | 1.2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laflamme, J.; Munson, A.D.; Grondin, P.; Arseneault, D. Anthropogenic Disturbances Create a New Vegetation Toposequence in the Gatineau River Valley, Quebec. Forests 2016, 7, 254. https://doi.org/10.3390/f7110254
Laflamme J, Munson AD, Grondin P, Arseneault D. Anthropogenic Disturbances Create a New Vegetation Toposequence in the Gatineau River Valley, Quebec. Forests. 2016; 7(11):254. https://doi.org/10.3390/f7110254
Chicago/Turabian StyleLaflamme, Jason, Alison D. Munson, Pierre Grondin, and Dominique Arseneault. 2016. "Anthropogenic Disturbances Create a New Vegetation Toposequence in the Gatineau River Valley, Quebec" Forests 7, no. 11: 254. https://doi.org/10.3390/f7110254
APA StyleLaflamme, J., Munson, A. D., Grondin, P., & Arseneault, D. (2016). Anthropogenic Disturbances Create a New Vegetation Toposequence in the Gatineau River Valley, Quebec. Forests, 7(11), 254. https://doi.org/10.3390/f7110254