
Can Retention Harvest Maintain Natural Structural Complexity? A Comparison of Post-Harvest and Post-Fire Residual Patches in Boreal Forest

Abstract
:1. Introduction
2. Materials and Methods
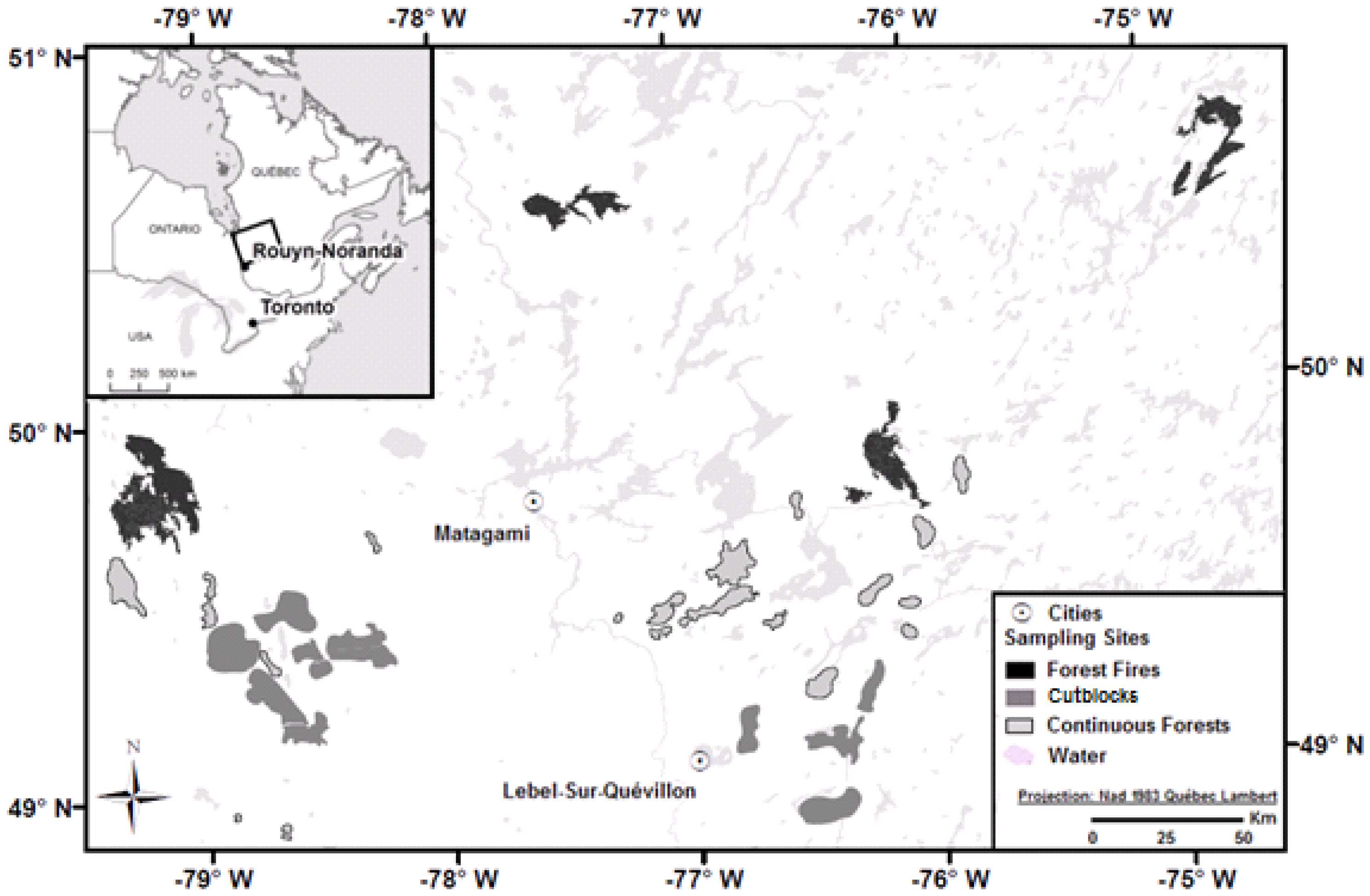
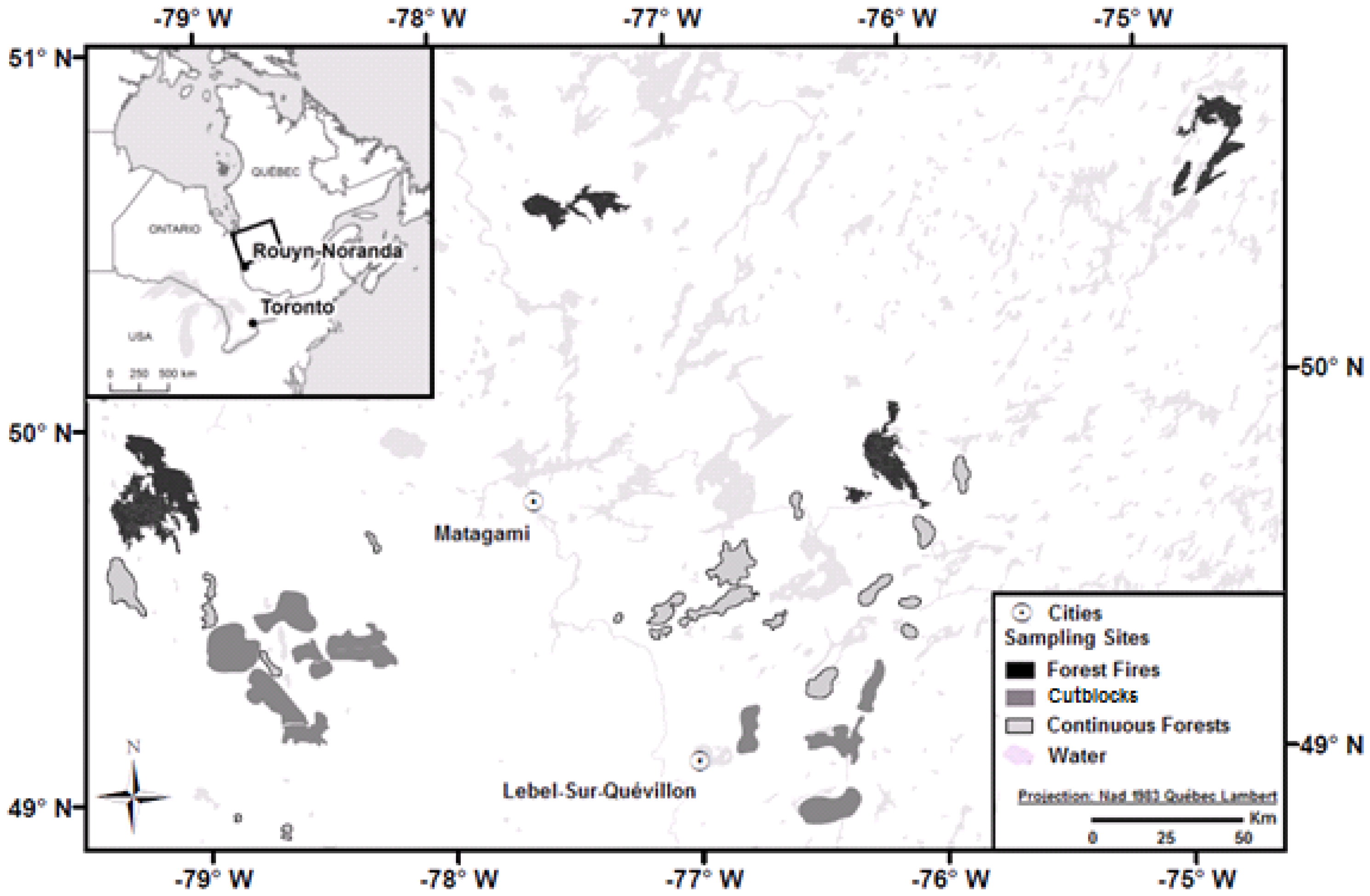
2.1. Study Area
2.2. Location of the Study Stands
2.3. Data Collection
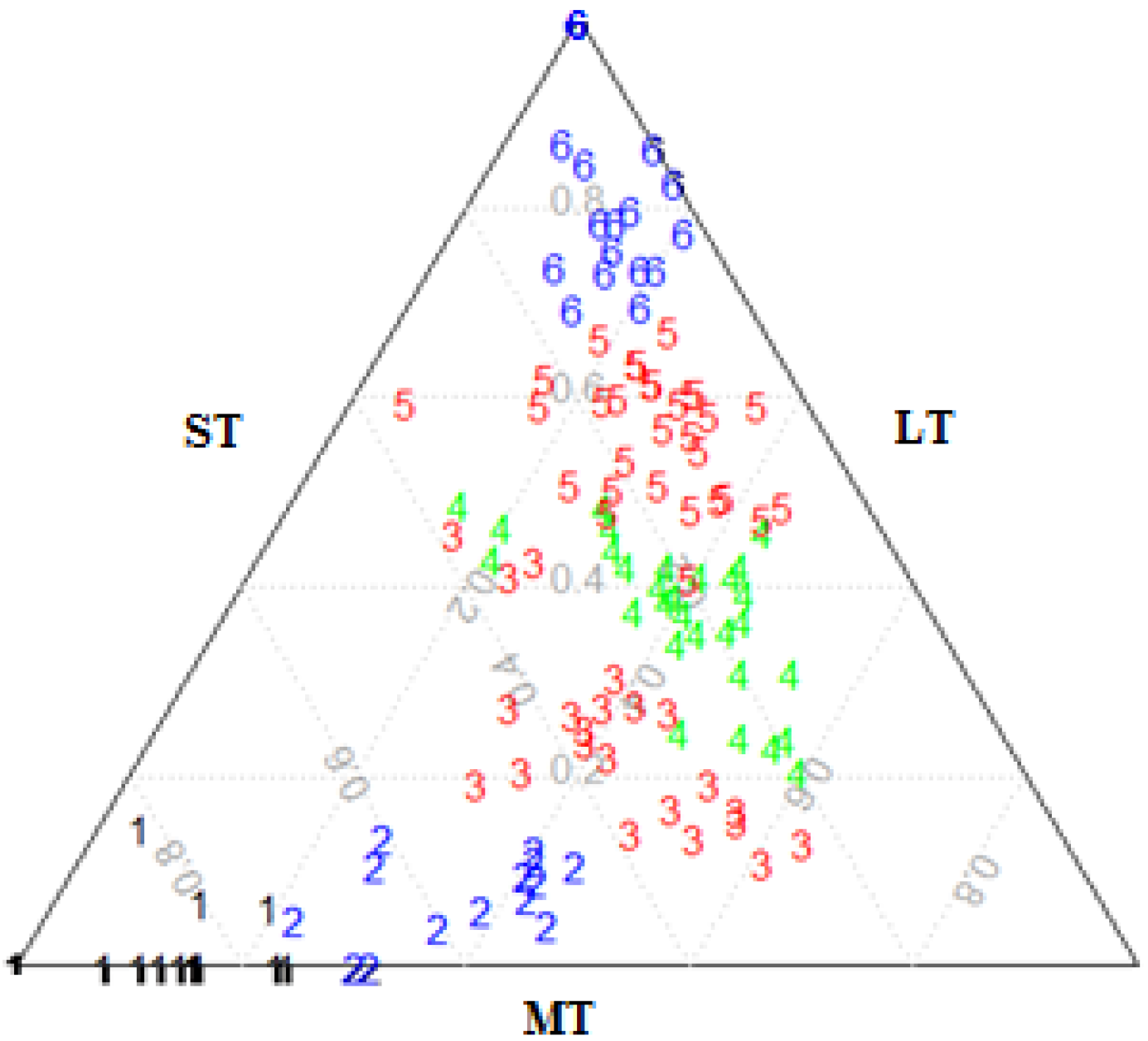
2.4. Structural Classification of Stands
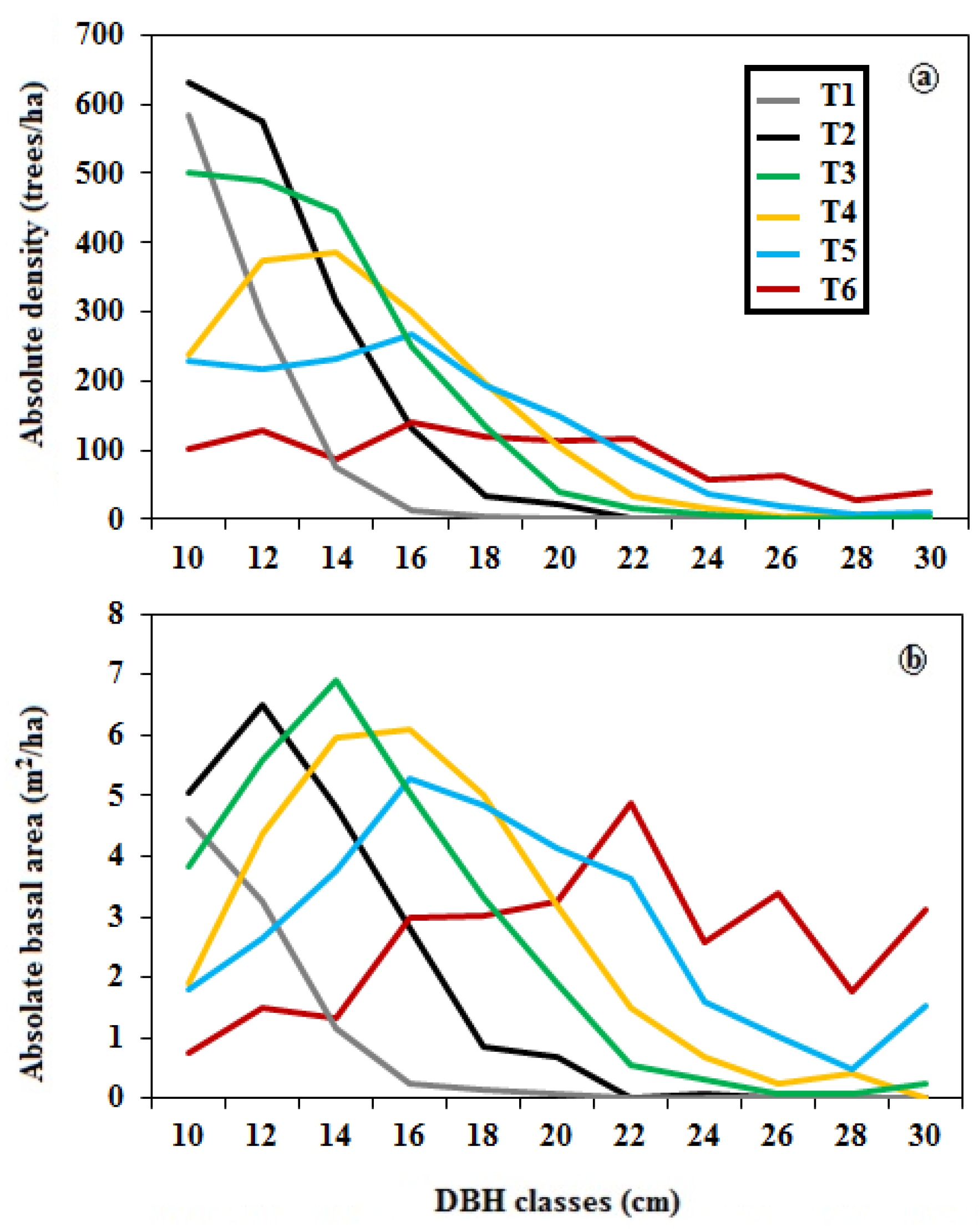
2.5. Description of Structural Types
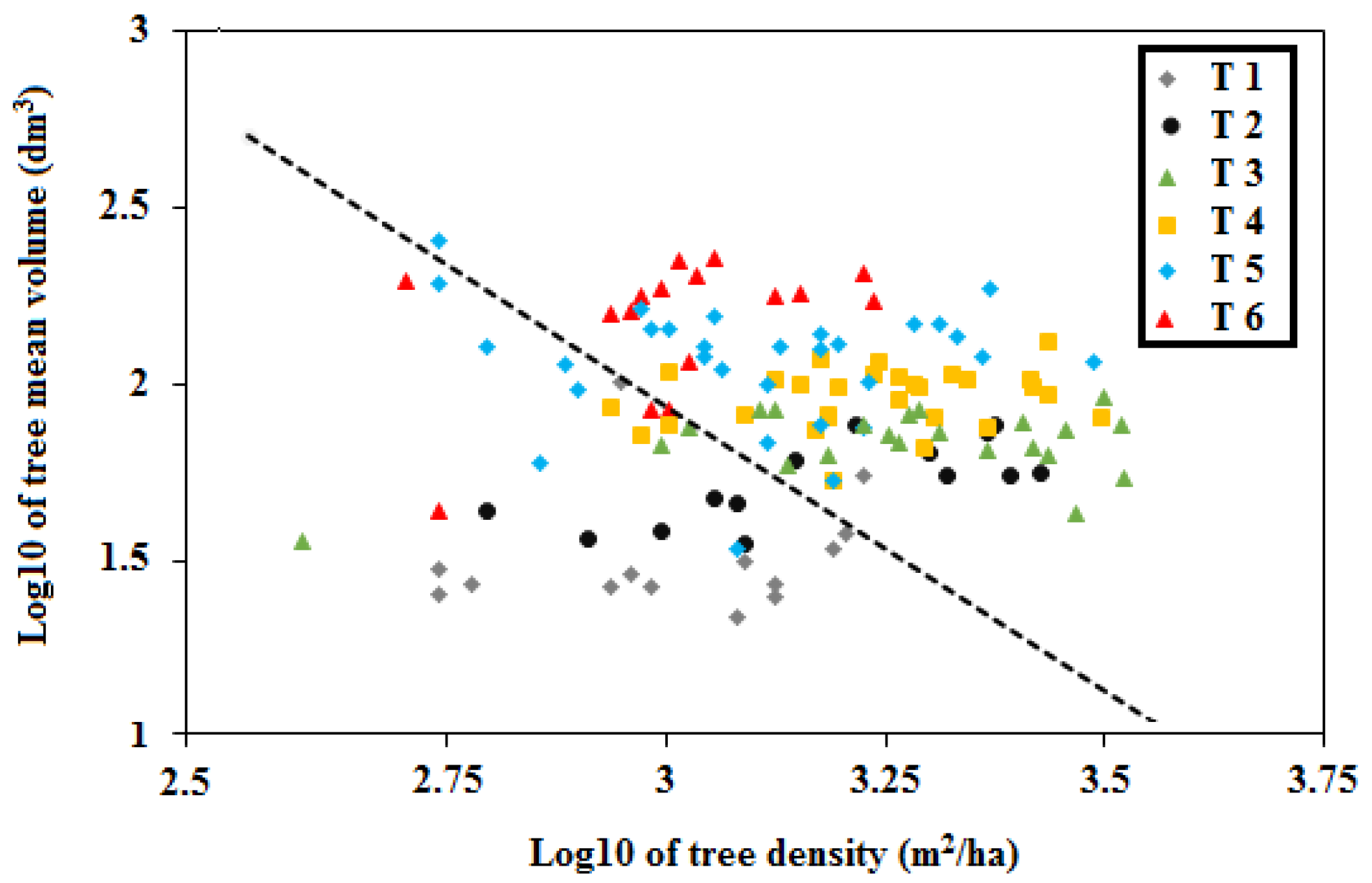
2.5.1. Internal Structure Complexity
2.5.2. Forest Canopy Closure
2.6. Factors Influencing Stand Structure
2.7. Structural Types in Post-Harvest Residual Patches versus Natural Stands
3. Results
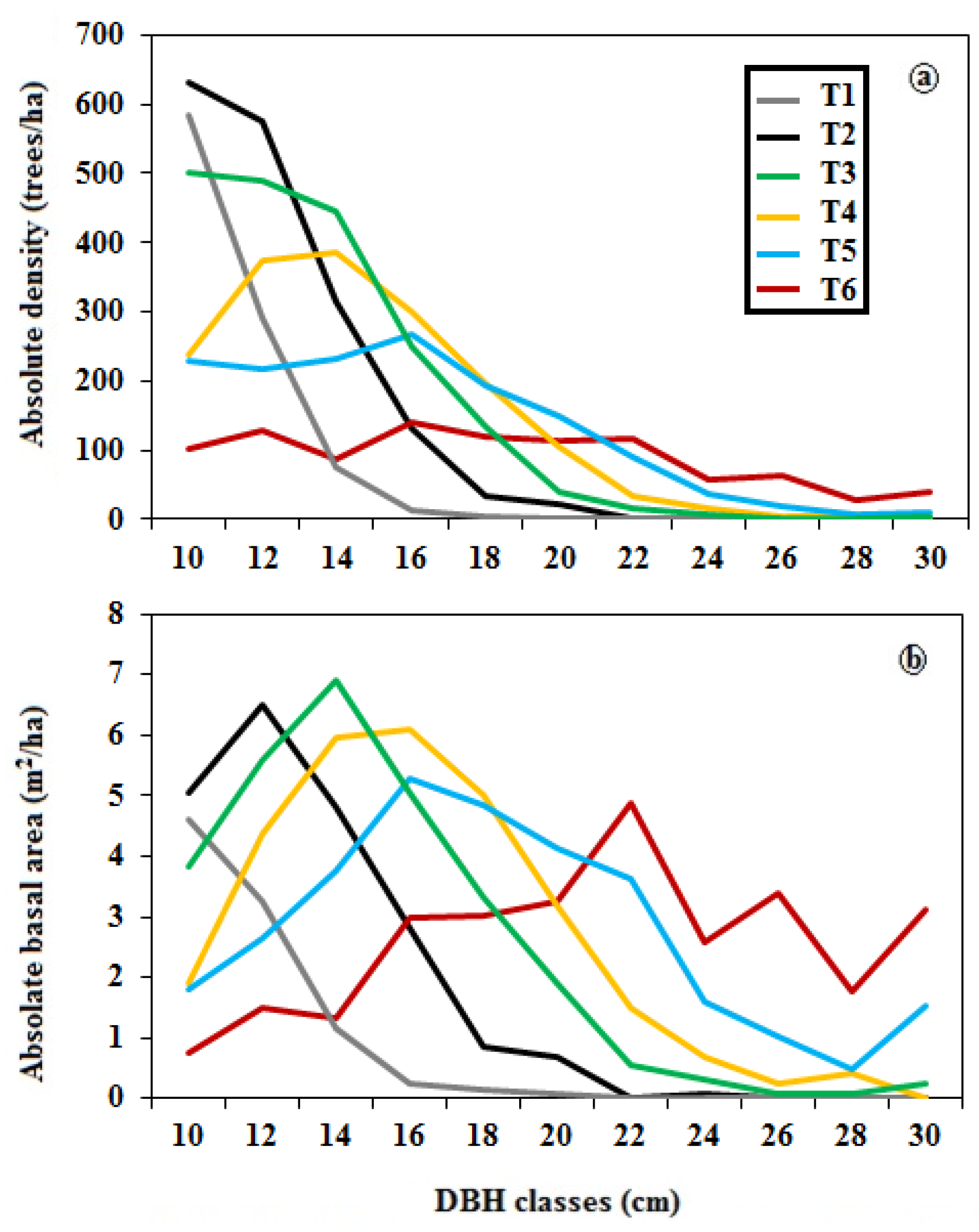
3.1. Structural Classification of Residual Patch Stands
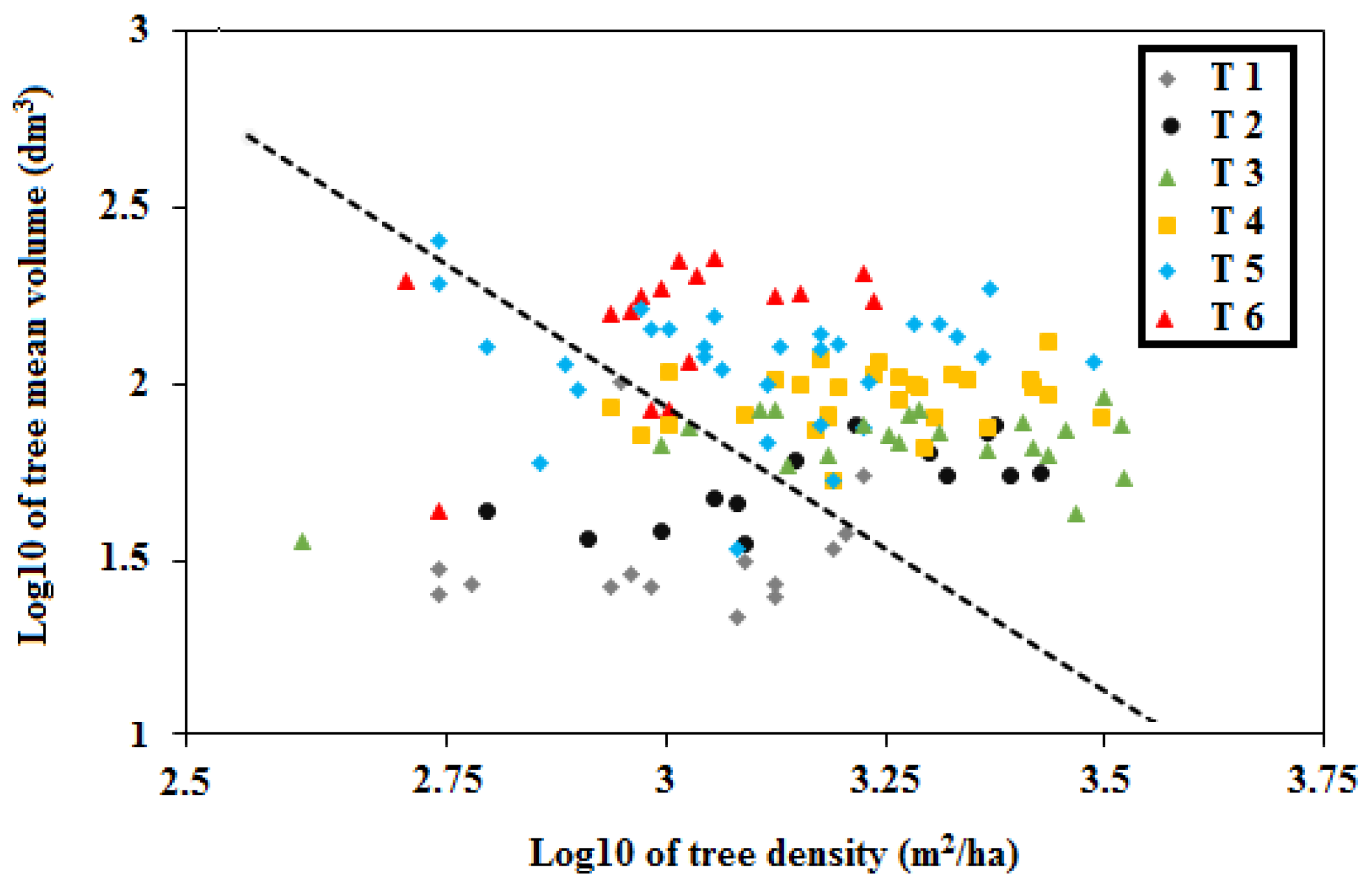
3.2. Complexity of Structural Stand Types
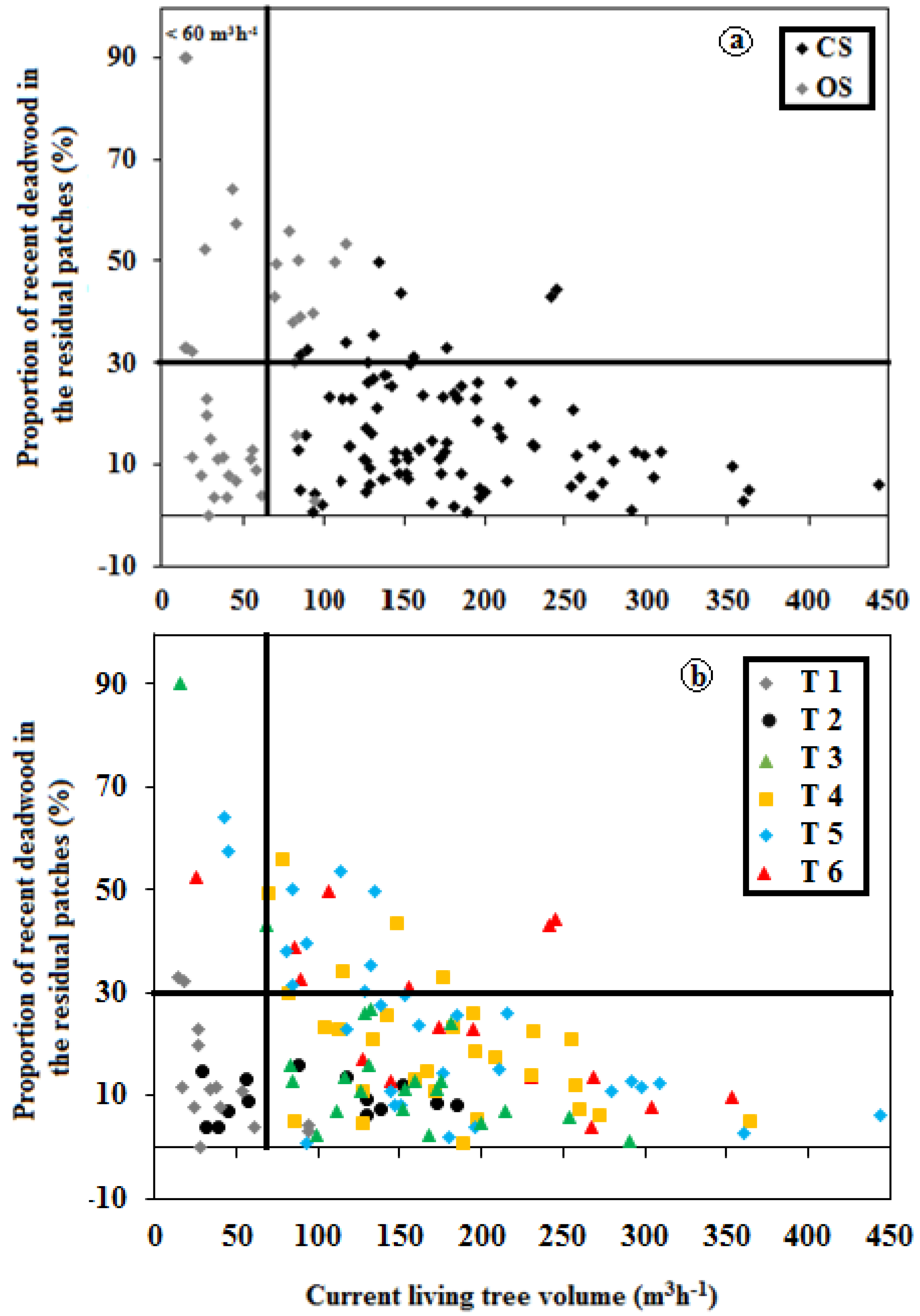
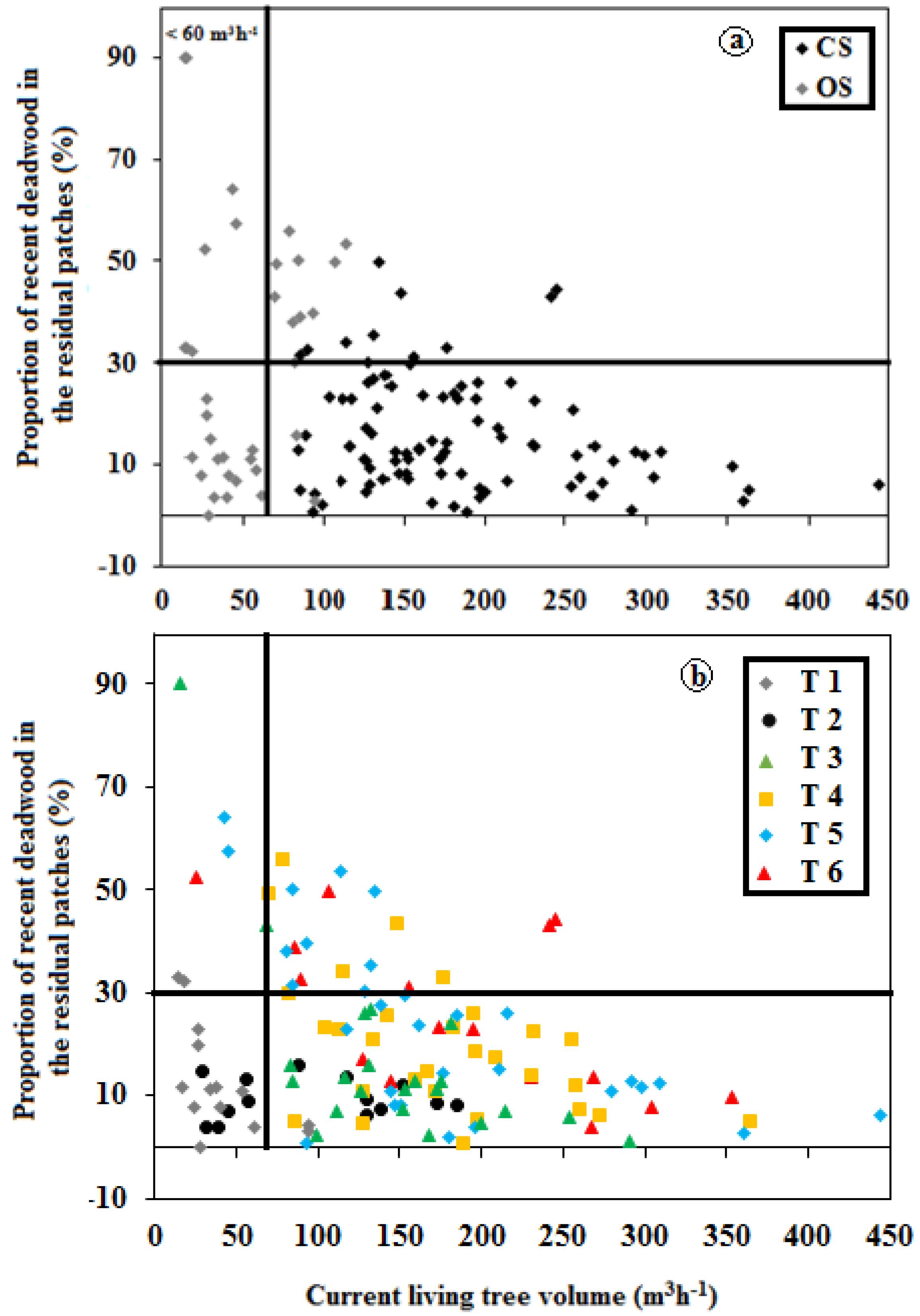
3.3. Influence of TSF on Structure
3.4. Structural Types: Post-Harvest versus Post-Fire Residual Patches
4. Discussion
5. Conclusions and Silvicultural Implications
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bergeron, Y.; Engelmark, O.; Harvey, B.; Morin, H.; Sirois, L. Key issues in disturbance dynamics in boreal forests: Introduction. J. Veg. Sci. 1998, 9, 464–468. [Google Scholar] [CrossRef]
- Hylander, K.; Johnson, S. In situ survival of forest bryophytes in small-scale refugia after an intense forest fire. J. Veg. Sci. 2010, 21, 1099–1109. [Google Scholar] [CrossRef]
- Perhans, K.; Appelgren, L.; Jonsson, F.; Nordin, U.; Söderström, B.; Gustafsson, L. Retention patches as potential refugia for bryophytes and lichens in managed forest landscapes. Biol. Conserv. 2009, 142, 1125–1133. [Google Scholar] [CrossRef]
- Schmiegelow, F.K.; Stepnisky, D.P.; Stambaugh, C.A.; Koivula, M. Reconciling salvage logging of boreal forests with a natural-disturbance management model. Conserv. Biol. 2006, 20, 971–983. [Google Scholar] [CrossRef] [PubMed]
- DeLong, S.C.; Kessler, W.B. Ecological characteristics of mature forest remnants left by wildfire. For. Ecol. Manag. 2000, 131, 93–106. [Google Scholar] [CrossRef]
- Madoui, A.; Leduc, A.; Gauthier, S.; Bergeron, Y. Spatial pattern analyses of post-fire residual stands in the black spruce boreal forest of western Quebec. Int. J. Wildland Fire 2011, 19, 1110–1126. [Google Scholar] [CrossRef]
- Bergeron, Y.; Drapeau, P.; Gauthier, S.; Lecomte, N. Using knowledge of natural disturbances to support sustainable forest management in the northern clay belt. For. Chron. 2007, 83, 326–337. [Google Scholar] [CrossRef]
- Cyr, D.; Gauthier, S.; Bergeron, Y.; Carcaillet, C. Forest management is driving the eastern north American boreal forest outside its natural range of variability. Front. Ecol. Environ. 2009, 7, 519–524. [Google Scholar] [CrossRef]
- Franklin, J.F.; Berg, D.R.; Thornburgh, D.A.; Tappeiner, J.C. Alternative silvicultural approaches to timber harvesting: Variable retention harvest systems. In Creating a Forestry for the 21st Century: The Science of Ecosystem Management; Island Press: Washington, DC, USA, 1997; pp. 111–139. [Google Scholar]
- Drapeau, P.; Nappi, A.; Giroux, J.-F.; Leduc, A.; Savard, J.-P. Distribution patterns of birds associated with snags in natural and managed eastern boreal forests. Ecol. Manag. Dead Wood Weste. For. 2002, 193–205. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; Van Pelt, R.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef]
- Gauthier, S.; Leduc, A.; Harvey, B.; Bergeron, Y.; Drapeau, P. Les perturbations naturelles et la diversité écosystémique. Nat. Can. 2001, 125, 10–17. [Google Scholar]
- Beese, W.; Dunsworth, B.; Zielke, K.; Bancroft, B. Maintaining attributes of old-growth forests in coastal BC through variable retention. For. Chron. 2003, 79, 570–578. [Google Scholar] [CrossRef]
- Hunter, M.L.J. Wildlife, Forests, and Forestry: Priciples of Managing Forests for Biological Diversity; Prentice-Hall: Englewood Cliffs, NJ, USA, 1990; p. 370. [Google Scholar]
- Kuuluvainen, T. Natural variability of forests as a reference for restoring and managing biological diversity in boreal fennoscandia. Silva Fenn. 2002, 36, 97–125. [Google Scholar] [CrossRef]
- Larson, B.C.; Oliver, C.D. Forest Stand Dynamics, 2nd ed.; John Wiley and Sons Inc.: New York, NY, USA, 1996; p. 520. [Google Scholar]
- Fenton, N.J.; Frego, K.A. Bryophyte (moss and liverwort) conservation under remnant canopy in managed forests. Biol Conserv. 2005, 122, 417–430. [Google Scholar] [CrossRef]
- Lachance, É.; Pothier, D.; Bouchard, M. Forest structure and understory plant communities inside and outside tree retention groups in boreal forests. Ecoscience 2013, 20, 252–263. [Google Scholar] [CrossRef]
- Gandhi, K.J.; Spence, J.R.; Langor, D.W.; Morgantini, L.E.; Cryer, K.J. Harvest retention patches are insufficient as stand analogues of fire residuals for litter-dwelling beetles in northern coniferous forests. Can. J. For. Res. 2004, 34, 1319–1331. [Google Scholar] [CrossRef]
- Bouchard, M.; Hébert, C. Beetle community response to residual forest patch size in managed boreal forest landscapes: Feeding habits matter. For. Ecol. Manag. 2016, 368, 63–70. [Google Scholar] [CrossRef]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology; John Wiley and Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Angelstam, P.; Kuuluvainen, T. Boreal forest disturbance regimes, successional dynamics and landscape structures: A european perspective. Ecol. Bull. 2004, 51, 117–136. [Google Scholar]
- Gauthier, S.; Bernier, P.; Kuuluvainen, T.; Shvidenko, A.; Schepaschenko, D. Boreal forest health and global change. Science 2015, 349, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Smith, F. Mortality in the Yukon: Post-Harvest Effects on Structural Retention. Master’s Thesis, University of Toronto, Toronto, ON, USA, 2010. [Google Scholar]
- Ouarmim, S.; Asselin, H.; Bergeron, Y.; Ali, A.A.; Hély, C. Stand structure in fire refuges of the eastern Canadian boreal mixedwood forest. For. Ecol. Manag. 2014, 324, 1–7. [Google Scholar] [CrossRef]
- Bolton, D.K.; Coops, N.C.; Wulder, M.A. Characterizing residual structure and forest recovery following high-severity fire in the western boreal of canada using landsat time-series and airborne lidar data. Remote Sens. Environ. 2015, 163, 48–163. [Google Scholar] [CrossRef]
- Moussaoui, L.; Fenton, N.J.; Leduc, A.; Bergeron, Y. Deadwood abundance in post-harvest and post-fire residual patches: An evaluation of patch temporal dynamics in black spruce boreal forest. For. Ecol. Manag. 2016, 368, 17–27. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Mackey, B.; Mullen, I.; McCarthy, M.; Gill, A.; Cunningham, R.; Donnelly, C. Factors affecting stand structure in forests–are there climatic and topographic determinants? For. Ecol. Manag. 1999, 123, 55–63. [Google Scholar] [CrossRef]
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar]
- Lindenmayer, D.; Franklin, J.; Lõhmus, A.; Baker, S.; Bauhus, J.; Beese, W.; Brodie, A.; Kiehl, B.; Kouki, J.; Pastur, G.M. A major shift to the retention approach for forestry can help resolve some global forest sustainability issues. Conserv. Lett. 2012, 5, 421–431. [Google Scholar] [CrossRef]
- Harper, K.A.; Drapeau, P.; Lesieur, D.; Bergeron, Y. Forest structure and composition at fire edges of different ages: Evidence of persistent structural features on the landscape. For. Ecol. Manag. 2014, 314, 131–140. [Google Scholar] [CrossRef]
- Work, T.T.; Shorthouse, D.P.; Spence, J.R.; Volney, W.J.A.; Langor, D. Stand composition and structure of the boreal mixedwood and epigaeic arthropods of the ecosystem management emulating natural disturbance (emend) landbase in northwestern Alberta. Can. J. For. Res. 2004, 34, 417–430. [Google Scholar] [CrossRef]
- Lavoie, S.; Ruel, J.-C.; Bergeron, Y.; Harvey, B.D. Windthrow after group and dispersed tree retention in eastern Canada. For. Ecol. Manag. 2012, 269, 158–167. [Google Scholar] [CrossRef]
- Urgenson, L.S.; Halpern, C.B.; Anderson, P.D. Level and pattern of overstory retention influence rates and forms of tree mortality in mature, coniferous forests of the pacific northwest, USA. For. Ecol. Manag. 2013, 308, 116–127. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J. Paysages Régionaux du Québec Méridional. Direction de la Gestion des Stocks Forestiers et Direction des Relations Publiques, Ministère des Ressources Naturelles du Québec; Les Publications du Quebec: Québec, QC, Canada, 1998.
- Bergeron, J.-F.; Grondin, P.; Blouin, J. Rapport de Classification Ecologique du Sous-Domaine Bioclimatique de la Pessière à Mousses de L’ouest; Ministere des Ressources Naturelles, Forêt Québec: Ville de Québec, QC, Canada, 1999.
- Blouin, J.; Berger, J. Guide de reconnaissance des types écologiques de la région écologique 6a—436 plaine du lac matagami et 6b—plaine de la baie de rupert. In Ministère des Ressources Naturelles du Québec, Forêt-Québec, Direction des Inventaires Forestiers, Division de la Classification Ecologique et Productivité des Stations; Forêt-Québec: Québec, QC, Canada, 2005; p. 188. [Google Scholar]
- Payette, S. Fire as a controlling process in the north american boreal forest. In A Systems Analysis of the Boreal Forest; Shugart, H.H., Leemans, R., Bonan, G.B., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 144–169. [Google Scholar]
- Bergeron, Y.; Gauthier, S.; Kafka, V.; Lefort, P.; Lesieur, D. Natural fire frequency for the eastern Canadian boreal forest: Consequences for sustainable forestry. Can. J. For. Res. 2001, 31, 384–391. [Google Scholar] [CrossRef]
- Bergeron, Y.; Gauthier, S.; Flannigan, M.; Kafka, V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 2004, 85, 1916–1932. [Google Scholar] [CrossRef]
- Gouvernement du Québec. Règlement sur les Normes D’intervention dans les Forêts du Domaine de L’état-loi sur les Forêts; LRQ c. F-4.1, a: Québec, QC, Canada, 1988; p. 171.
- Bergeron, Y.; Harvey, B.; Leduc, A.; Gauthier, S. Forest management guidelines based on natural disturbance dynamics: Stand-and forest-level considerations. For. Chron. 1999, 75, 49–54. [Google Scholar] [CrossRef]
- Harvey, B.D.; Leduc, A.; Gauthier, S.; Bergeron, Y. Stand-landscape integration in natural disturbance-based management of the southern boreal forest. For. Ecol. Manag. 2002, 155, 369–385. [Google Scholar] [CrossRef]
- Chaieb, C.; Fenton, N.J.; Lafleur, B.; Bergeron, Y. Can we use forest inventory mapping as a coarse filter in ecosystem based management in the black spruce boreal forest? Forests 2015, 6, 1195–1207. [Google Scholar] [CrossRef]
- Harper, K.A.; Lesieur, D.; Bergeron, Y.; Drapeau, P. Forest structure and composition at young fire and cut edges in black spruce boreal forest. Can. J. For. Res. 2004, 34, 289–302. [Google Scholar] [CrossRef]
- Fortin, M.; DeBlois, J.; Bernier, S.; Blais, G. Mise au point d’un tarif de cubage général pour les forêts québécoises: Une approche pour mieux évaluer l’incertitude associée aux prévisions. For. Chron. 2007, 83, 754–765. [Google Scholar] [CrossRef]
- Van Wagner, C. The line intersect method in forest fuel sampling. For. Sci. 1968, 14, 20–26. [Google Scholar]
- Thomas, J.W.; Anderson, R.G.; Maser, C.; Bull, E.L. Wildlife Habitats in Managed Forests of the Blue Mountains of Oregon and Washington; United States Department of Agriculture, Forest Service, Agricultural Handbook; US Forest Service: Washington, DC, USA, 1979; p. 553.
- McGarigal, K.; Marks, B.J. Spatial pattern analysis program for quantifying landscape structure. Dolores (CO) PO Box 1994, 606, 67. [Google Scholar]
- Wagner, C.V. Age-class distribution and the forest fire cycle. Can. J. For. Res. 1978, 8, 220–227. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Silvics of north america. Volume 1. Conifers. In Agriculture Handbook; Forest Service, USDA: Washington, DC, USA, 1990; p. 654. [Google Scholar]
- Moss, I. Stand Structure Classification, Succession, and Mapping Using Lidar. Ph.D. Thesis, The University of British Columbia, Vancouver, BC, Canada, 2012. [Google Scholar]
- Nock, R.; Nielsen, F. On weighting clustering. IEEE Trans. Pattern Anal. Mach. Intell. 2006, 28, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: Breinigsville, PA, USA, 2011; p. 306. [Google Scholar]
- Brock, G.; Pihur, V.; Datta, S.; Datta, S. Clvalid, an r package for cluster validation. J. Stat. Softw. 2008, 1, 22–25. [Google Scholar]
- De Liocourt, F.D. De l’amenagement des sapinieres. Bull. Soc. Franche-Comté Belfort 1898, 4, 396–409. [Google Scholar]
- Lessard, G.; Côté, S. Détermination des paramètres des forêts aptes au régime du jardinage (phase i). Centre Collégial de Transfert de Technologie en Foresterie (CERFO), Rapport du CERFO 2005, 4, 289. [Google Scholar]
- Pinheiro, J.; Bates, D. Mixed-Effects Models in S and S-Plus; Chambers, J., Eddy, W., Hardle, W., Sheather, S., Tierney, L., Eds.; Springer: New York, NY, USA, 2000. [Google Scholar]
- R Development-CORE-TEAM. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Available online: http://www.R-project.org/ (accessed on 12 December 2011).
- Newton, P.F. A decision-support system for forest density management within upland black spruce stand-types. Environ. Model. Softw. 2012, 35, 171–187. [Google Scholar] [CrossRef]
- Simard, M.; Bernier, P.Y.; Bergeron, Y.; Pare, D.; Guérine, L. Paludification dynamics in the boreal forest of the james bay lowlands: Effect of time since fire and topography. Can J. For. Res. 2009, 39, 546–552. [Google Scholar] [CrossRef]
- Nlungu-Kweta, P.; Leduc, A.; Bergeron, Y. Climate and disturbance regime effects on aspen (Populus tremuloides Michx) stand structure and composition along an east-west transect in Canada’s boreal forest. Int. J. For. Res. 2016. [Google Scholar] [CrossRef]
- Heikkala, O.; Seibold, S.; Koivula, M.; Martikainen, P.; Müller, J.; Thorn, S.; Kouki, J. Retention forestry and prescribed burning result in functionally different saproxylic beetle assemblages than clear-cutting. For. Ecol. Manag. 2016, 359, 51–58. [Google Scholar] [CrossRef]
- Clark, D.F.; Antos, J.A.; Bradfield, G.E. Succession in sub-boreal forests of west-central british columbia. J. Veg. Sci. 2003, 14, 721–732. [Google Scholar] [CrossRef]
- Lee, P.C.; Crites, S.; Nietfeld, M.; Nguyen, H.V.; Stelfox, J.B. Characteristics and origins of deadwood material in aspen-dominated boreal forests. Ecol. Appl. 1997, 7, 691–701. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y. Stand structural dynamics of north american boreal forests. Crit. Rev. Plant Sci. 2006, 25, 115–137. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Burton, P. Canopy and age structures of some old sub-boreal picea stands in British Columbia. J. Veg. Sci. 1997, 8, 615–625. [Google Scholar] [CrossRef]
- Boucher, D.; Gauthier, S.; De Grandpré, L. Structural changes in coniferous stands along a chronosequence and a productivity gradient in the northeastern boreal forest of québec. Ecoscience 2006, 13, 172–180. [Google Scholar] [CrossRef]
- Chen, H.Y.; Popadiouk, R.V. Dynamics of north american boreal mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Smyth, C.; Schieck, J.; Boutin, S.; Wasel, S. Influence of stand size on pattern of live trees in mixedwood landscapes following wildfire. For. Chron. 2005, 81, 125–132. [Google Scholar] [CrossRef]
- Brassard, B.W.; Chen, H.Y.; Wang, J.R.; Duinker, P.N. Effects of time since stand-replacing fire and overstory composition on live-tree structural diversity in the boreal forest of central Canada. Can. J. For. Res. 2008, 38, 52–62. [Google Scholar] [CrossRef]
- Simard, M.; Lecomte, N.; Bergeron, Y.; Bernier, P.Y.; Paré, D. Forest productivity decline caused by successional paludification of boreal soils. Ecol. Appl. 2007, 17, 1619–1637. [Google Scholar] [CrossRef] [PubMed]
- Ruel, J.-C. Understanding windthrow: Silvicultural implications. For. Chron. 1995, 71, 434–445. [Google Scholar] [CrossRef]
- Laamrani, A.; Valeria, O.; Bergeron, Y.; Fenton, N.; Cheng, L.Z.; Anyomi, K. Effects of topography and thickness of organic layer on productivity of black spruce boreal forests of the canadian clay belt region. For. Ecol. Manag. 2014, 330, 144–157. [Google Scholar] [CrossRef]
- Cyr, D.; Gauthier, S.; Bergeron, Y. Scale-dependent determinants of heterogeneity in fire frequency in a coniferous boreal forest of eastern Canada. Landsc. Ecol. 2007, 22, 1325–1339. [Google Scholar] [CrossRef]
- Rich, R.L.; Frelich, L.E.; Reich, P.B. Wind-throw mortality in the southern boreal forest: Effects of species, diameter and stand age. J. Ecol. 2007, 95, 1261–1273. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Mayerhofer, M.S.; Biswas, S.R.; Esseen, P.A.; Hylander, K.; Stewart, K.J.; Mallik, A.U.; Drapeau, P.; Jonsson, B.G. Edge influence on vegetation at natural and anthropogenic edges of boreal forests in Canada and Fennoscandia. J. Ecol. 2015, 103, 550–562. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | T1 (n = 14) | T2 (n = 14) | T3 (n = 22) | T4 (n = 29) | T5 (n = 30) | T6 (n = 16) |
|---|---|---|---|---|---|---|
| Living Wood | ||||||
| Mean height of living trees (m) | 9.8 ± 0.3 a | 12.5 ± 0.5 b | 13.7 ± 0.4 bc | 14.8 ± 0.3 c | 16.1 ± 0.4 e | 16.3 ± 0.6 e |
| Mean tree diameter at breast height (cm) | 11.1 ± 0.2 a | 12.9 ± 0.4 ab | 13.7 ± 0.3 bc | 14.7 ± 0.3 ce | 15.4 ± 0.3 e | 17.7 ± 0.8 d |
| Mean total tree density (trees/ha) | 1153.6 ± 105.2 b | 1712.5 ± 182.1 ab | 2114.8 ± 174.6 a | 1881.9 ± 108.5 a | 1449.1 ± 109.6 b | 1139.1 ± 85.8 b |
| Mean tree basal area (m2/h) | 11 ± 1.1 a | 20.5 ± 2.3 a | 30.6 ± 2.5 b | 33.1 ± 1.9 b | 31.2 ± 2.4 b | 34 ± 2.4 b |
| Mean total tree volume (m3/h) | 40.82 ± 7 a | 98.1 ± 14.7 ac | 146.2 ± 13.1 bc | 173.8 ± 12.8 bc | 173.3 ± 17.2 b | 188.4 ± 22.5 b |
| Mean sapling density (saplings/ha) | 4941.7 ± 771.4 a | 4177.8 ± 867.8 a | 2081.3 ± 486.7 b | 1356.3 ± 419.4 b | 1718.2 ± 346.5 b | 1353.8 ± 352.9 b |
| Mean sapling basal area (m2/h) | 7.2 ± 1.2 a | 5.4 ± 0.6 ac | 3.9 ± 0.9 bc | 2.1 ± 0.5 b | 3.1 ± 0.7 bc | 1.7 ± 0.5 b |
| Mean small timber basal area (m2/h) | 9.5 ± 0.9 ab | 11.5 ± 1.2 a | 10.6 ± 1 a | 7.2 ± 0.4 b | 4.4 ± 0.4 c | 2.2 ± 0.4 c |
| Mean medium timber basal area (m2/h) | 1.5 ± 0.3 a | 7.6 ± 1 c | 12.8 ± 1.2 b | 13.6 ± 1.1 b | 9.1 ± 1 c | 4.8 ± 0.7 ac |
| Mean large timber basal area (m2/h) | 0.2 ± 0.1 a | 1.6 ± 0.3 a | 7.3 ± 0.8 b | 12.4 ± 0.9 c | 17.2 ± 1.3 d | 26.8 ± 2.1 e |
| Deadwood | ||||||
| Mean diameter of snags (cm) | 11.1 ± 0.4 a | 12.1 ± 0.4 ab | 12.8 ± 0.4 ab | 13.6 ± 0.5 bc | 15.6 ± 0.5 c | 18.5 ± 1.1 d |
| Mean total volume of snags (m3/h) | 2.2 ± 0.6 a | 5.5 ±1.1 a | 11.6 ± 1.7 ab | 19.3 ± 2.7 b | 31.3 ± 3.5 bc | 36.3 ± 5.5 c |
| Mean total volume of logs (m3/h) | 7.9 ± 1.6 a | 19.22 ± 4.2 a | 53.1 ± 12.5 ab | 75.5 ± 11.8 b | 85.6 ± 9.1 bc | 123.7 ±19.5 c |
| Recent deadwood volume (m3/h) | 4.6 ± 0.7 a | 10.5 ± 1.8 a | 26.6 ± 6.1 ab | 41.5 ± 5.3 bc | 47.7 ± 5.9 bc | 62.1 ± 13.6 c |
| Old deadwood volume (m3/h) | 5.4 ± 1.6 a | 14.2 ± 2.8 a | 38.1 ± 10.8 ab | 53.3 ± 10.1 b | 69.1 ± 7.1 bc | 98.1 ± 12.5 c |
| Environmental Factors | ||||||
| Thickness of the organic layer (cm) | 43.4 ± 6.4 a | 38.6 ± 4.8 a | 44.7 ± 9.1 a | 35.8 ± 4 a | 41.6 ± 4.5 a | 49.2 ± 9.1 a |
| Mean shape index | 1.2 ± 0.07 a | 1.1 ± 0.02 a | 1.2 ± 0.05 a | 1.1 ± 0.03 a | 1.3 ± 0.05 a | 1.2 ± 0.03 a |
| Area of residual patch (ha) | 4.5 ± 2.4 a | 10.6 ± 4.9 a | 8.4 ± 2.3 a | 11.7 ± 2.4 a | 7.4 ± 1.3 a | 9.3 ± 4.2 a |
| Time since the last fire (year) | 97 ± 7.3 a | 115.2 ± 13.6 ab | 120 ± 12.1 ab | 126.7 ± 12.7 ab | 149.2 ± 11.2 b | 143.6 ± 11.7 b |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moussaoui, L.; Fenton, N.J.; Leduc, A.; Bergeron, Y. Can Retention Harvest Maintain Natural Structural Complexity? A Comparison of Post-Harvest and Post-Fire Residual Patches in Boreal Forest. Forests 2016, 7, 243. https://doi.org/10.3390/f7100243
Moussaoui L, Fenton NJ, Leduc A, Bergeron Y. Can Retention Harvest Maintain Natural Structural Complexity? A Comparison of Post-Harvest and Post-Fire Residual Patches in Boreal Forest. Forests. 2016; 7(10):243. https://doi.org/10.3390/f7100243
Chicago/Turabian StyleMoussaoui, Louiza, Nicole J. Fenton, Alain Leduc, and Yves Bergeron. 2016. "Can Retention Harvest Maintain Natural Structural Complexity? A Comparison of Post-Harvest and Post-Fire Residual Patches in Boreal Forest" Forests 7, no. 10: 243. https://doi.org/10.3390/f7100243
APA StyleMoussaoui, L., Fenton, N. J., Leduc, A., & Bergeron, Y. (2016). Can Retention Harvest Maintain Natural Structural Complexity? A Comparison of Post-Harvest and Post-Fire Residual Patches in Boreal Forest. Forests, 7(10), 243. https://doi.org/10.3390/f7100243