4.1. Productivity Responses by Site Quality, Locale, Emission Scenario, and Species
The results of simulating the effects of climate change on the productivity of black spruce and jack pine plantations by site quality, geographic region, and emission scenario, using the SSDMMs, enabled a comparative evaluation of rotational productivity outcomes arising from anthropogenic-induced changes in growing conditions. Furthermore, analyzing responses in terms of stand development rates, rotational yields and productivity indices across a broader range of geographic locations and commitment periods, provided a more comprehensive assessment of climate change effects and their associated variability, than previous attempts. For example, Newton [
22] employed a similar approach but the analysis was restricted to analyzing the responses of jack pine plantations at the north-eastern and north-central locales for only the first two commitment periods (2011–2070) given computational limitations of the then available biophysical site-specific height-age function. Although that analysis suggested that jack pine would experience consequential declines in stand development rates resulting in decreases in rotational mean sizes, biomass yields, recoverable end-product volumes, and economic worth under both the B1 and A2 emission scenarios for the higher site qualities, the results were not uniform across all site qualities examined. However, with the advent of the development of the new biophysical site-specific height-age models by Sharma and others [
16] applicable to more than a single species, combined with the computational enhancements which corrected for the discrete and dramatic changes in height estimates predicted to occur beginning of the latter two commitment periods, it was possible to: (1) refresh the jack pine simulations by including additional geographic locations (north-western Ontario), deploy operationally relevant rotation lengths, and account for the critically important third commitment period during which growing conditions are expected to rapidly deteriorate, particularly under the A2 emission scenario; and (2) assess the consequences of a changing climate on another commercially important species in terms of Ontario’s future wood supply, i.e., black spruce. As a result, this study provides a more inclusive assessment of simulated climate change effects on the productivity of these species, than that of the previous attempt.
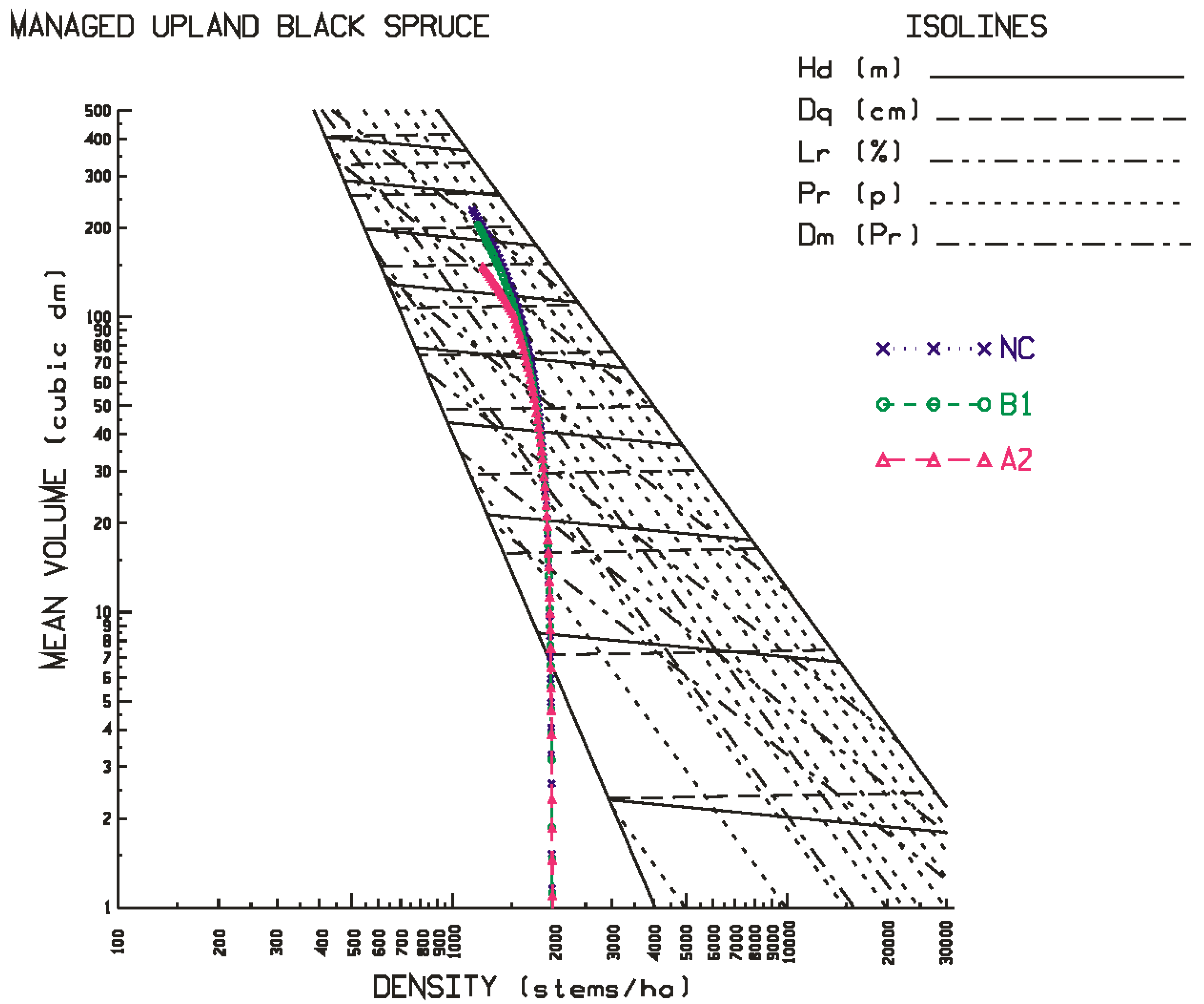
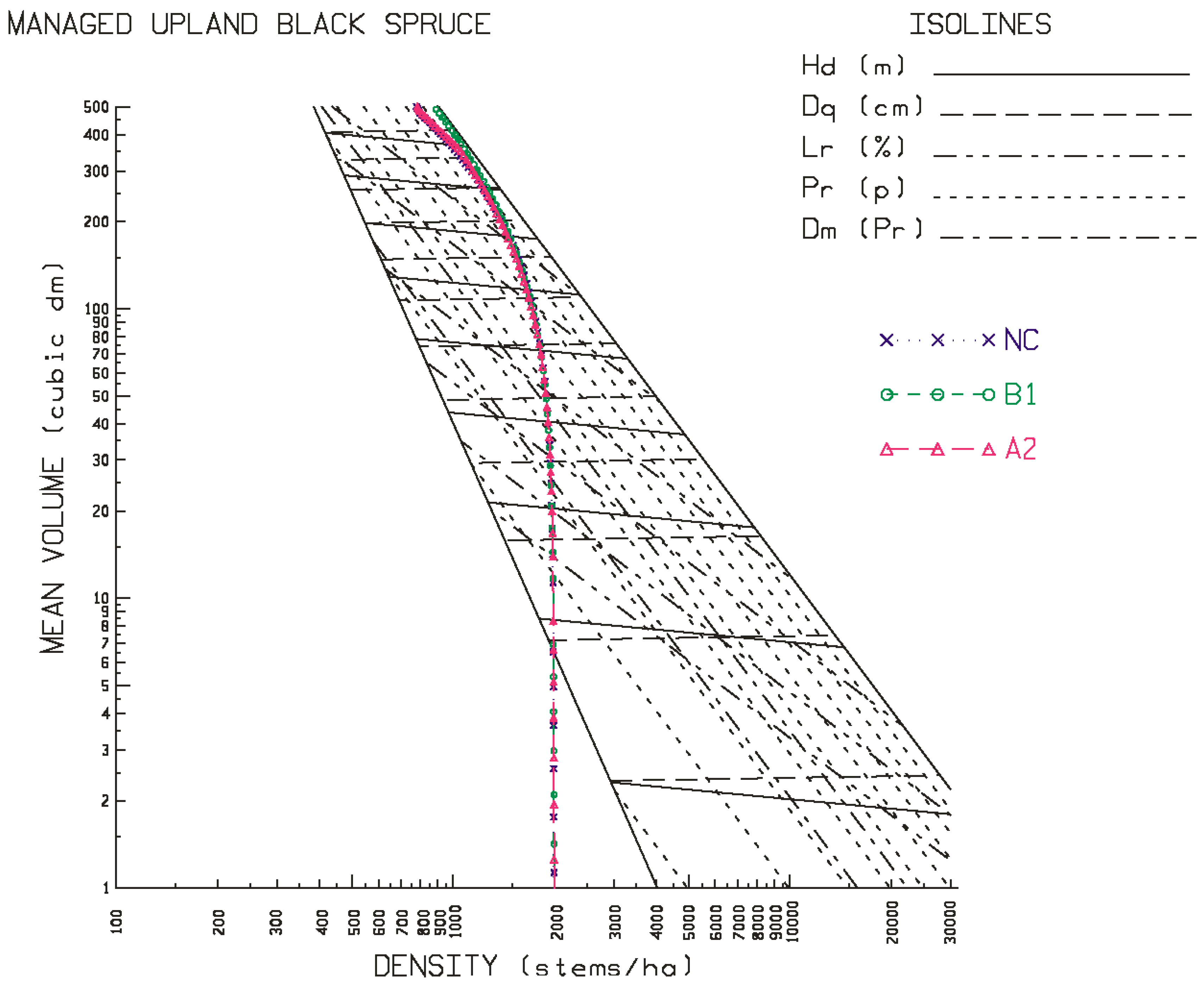
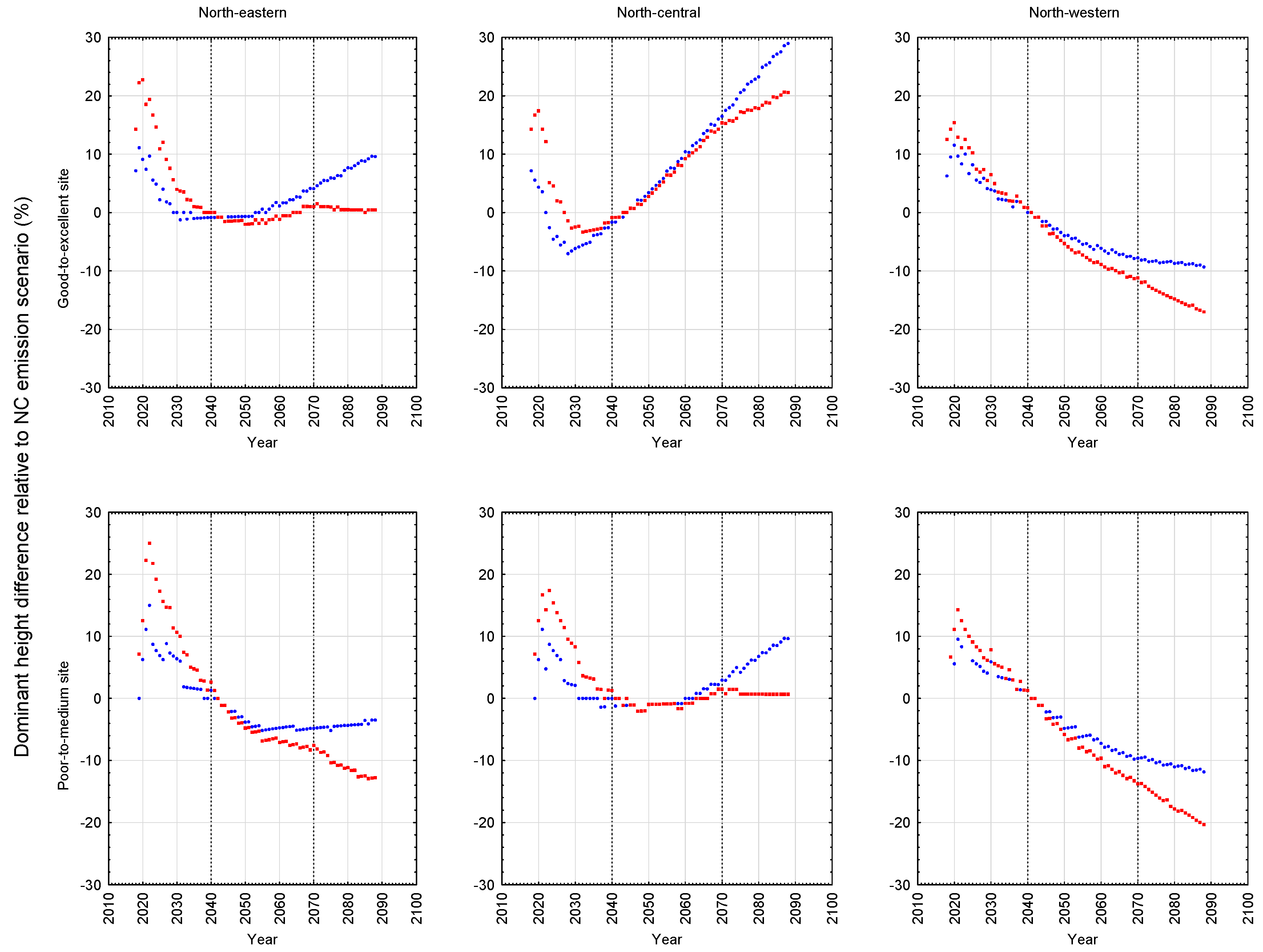
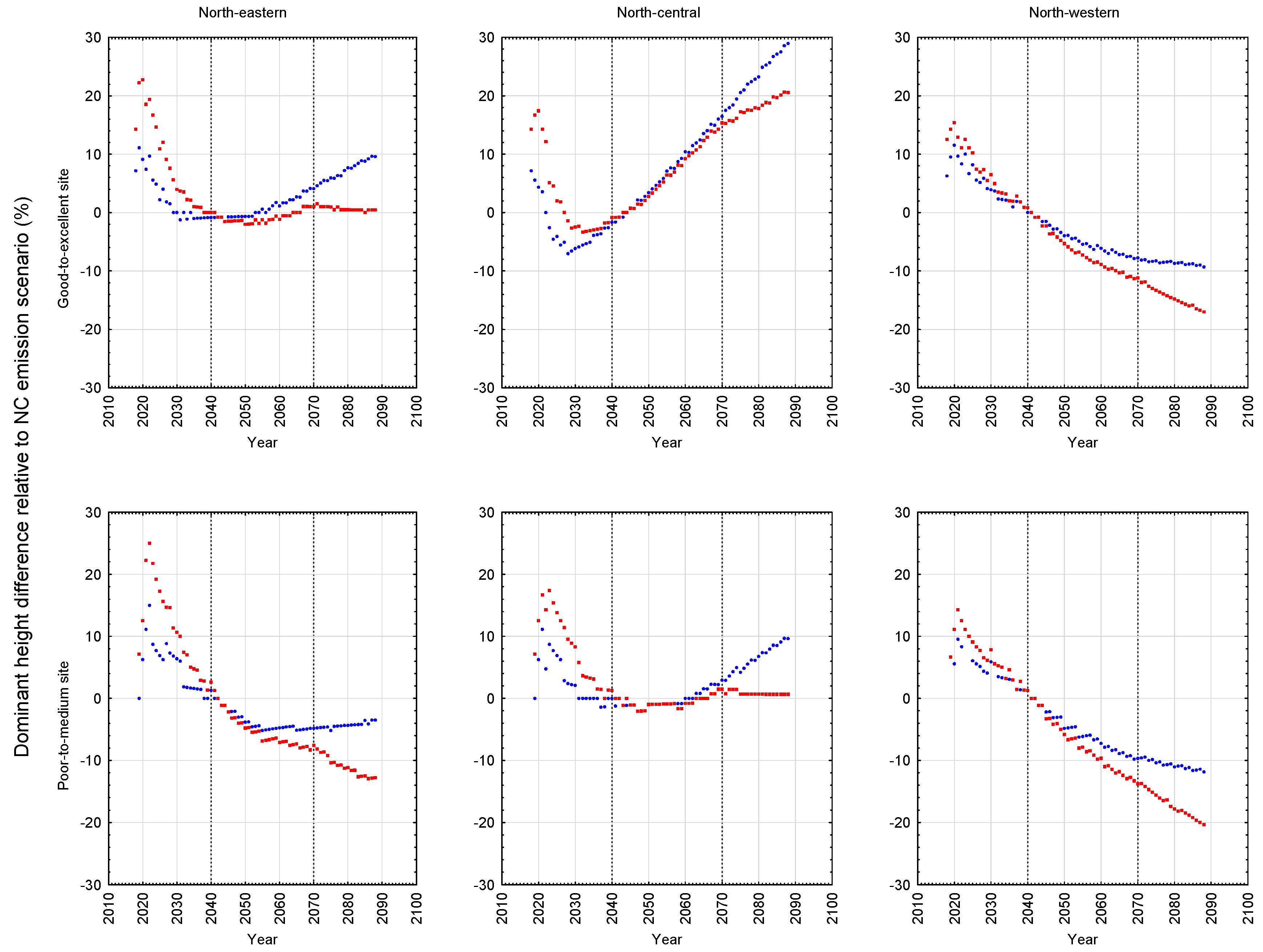
The results of these simulations indicated that black spruce plantations situated on both site qualities at the north-western location and the lower site quality at the north-eastern location, were negatively affected: declines in stand development rates resulted in decreases in rotational mean sizes, biomass yields, recoverable end-product volumes, and economic worth, arising from increased temperature and precipitation rates, with the A2 scenario exhibiting the largest effect. Conversely, black spruce plantations situated on both site qualities at the north-central location and the higher site quality at the north-eastern location were minimally or positively affected based on the same response metrics, with the B1 scenario exhibiting the largest positive effect. Across all locales examined, the black spruce plantations established at the north-western location exhibited the largest declines in productivity relative to those established at the north-central and north-eastern locales.
Model predictions of the productivity consequences of climate change for boreal species have exhibited considerable variability and hence arriving at a consensus on its impacts has been elusive. In the case of black spruce, a similar finding of positive effects was reported by Coulombe et al. [
23] for natural black spruce stands situated in north-central Quebec. Based on a process-based model (StandLeaf), they predicted an increase in productivity (merchantable volume per unit area) of approximately 22%–37% at 100 years (2110) relative to that which would be produced under the current climate. Conversely, Girardin [
24] using the same model for black spruce but in western Canada, predicted productivity declines which were attributed to the negative effect of increased temperature on soil moisture regimes. The productivity declines observed for the sites at the north-western locale in this study are consistent with this inference: north-western sites are predicted to be warmer and dryer than those at the north-central and north-eastern sites (
Table 1). Furthermore, projected temperature and moisture regimes under the A2 emission scenario for the Quebec locations that were utilized by Coulombe et al. [
23], indicated that these sites were wetter (23% more precipitation fell during the growing season) and was not as warm (11% cooler during the growing season) relative to that predicted for the north-western locale. Collectively, these inferences support the temperature-dependent moisture deficit and associated productivity decline relationship that may occur on some black spruce sites but not on others.
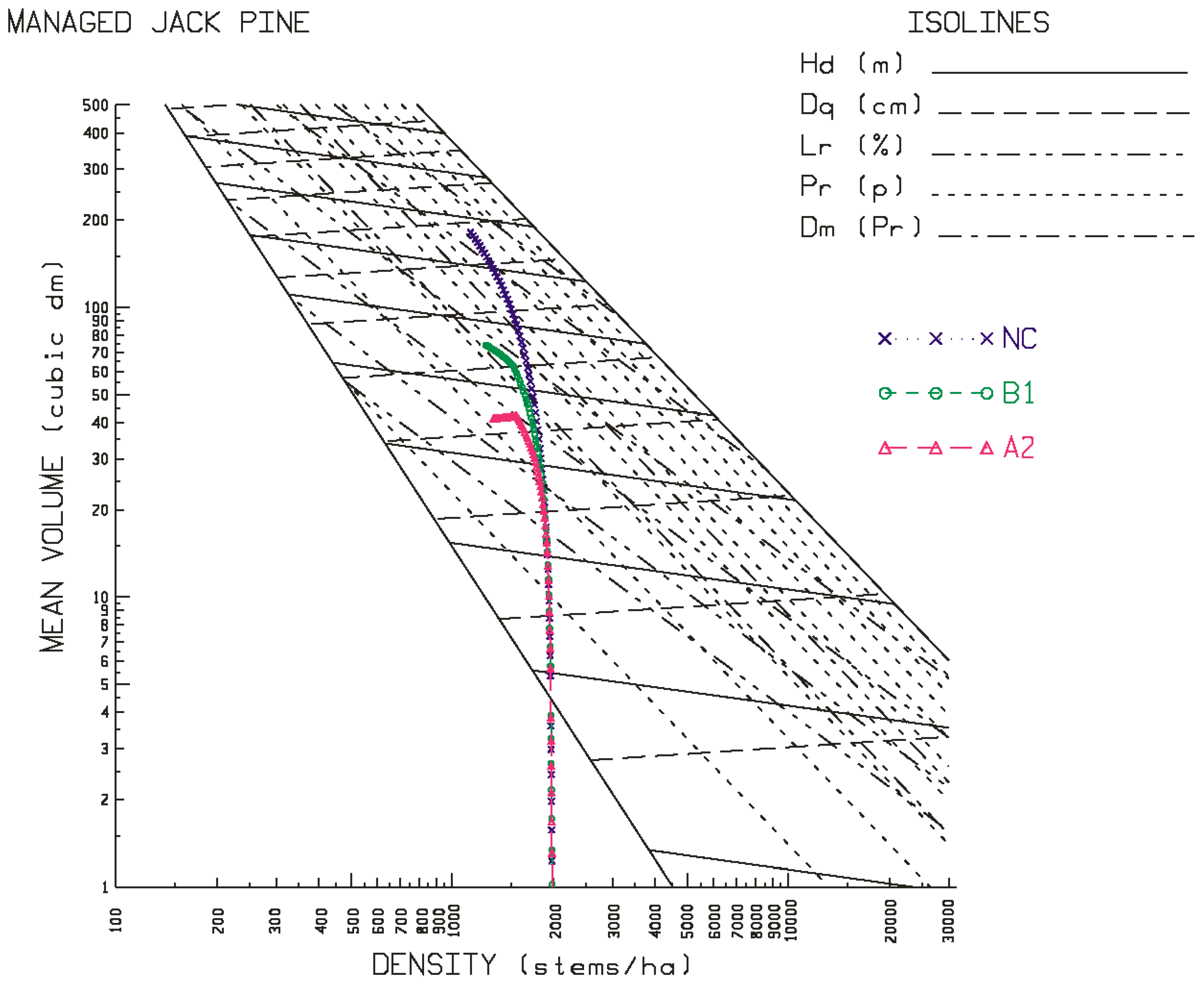
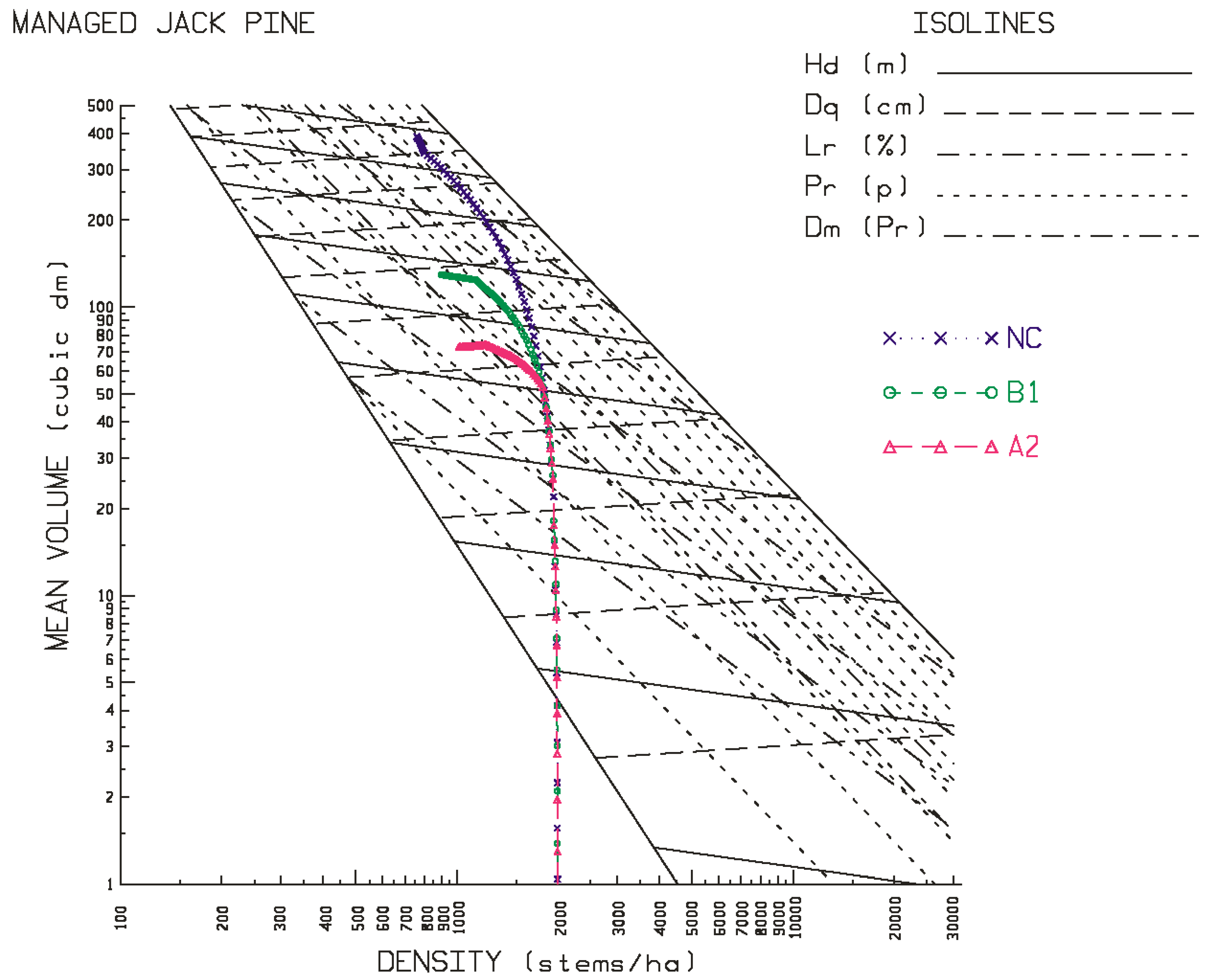
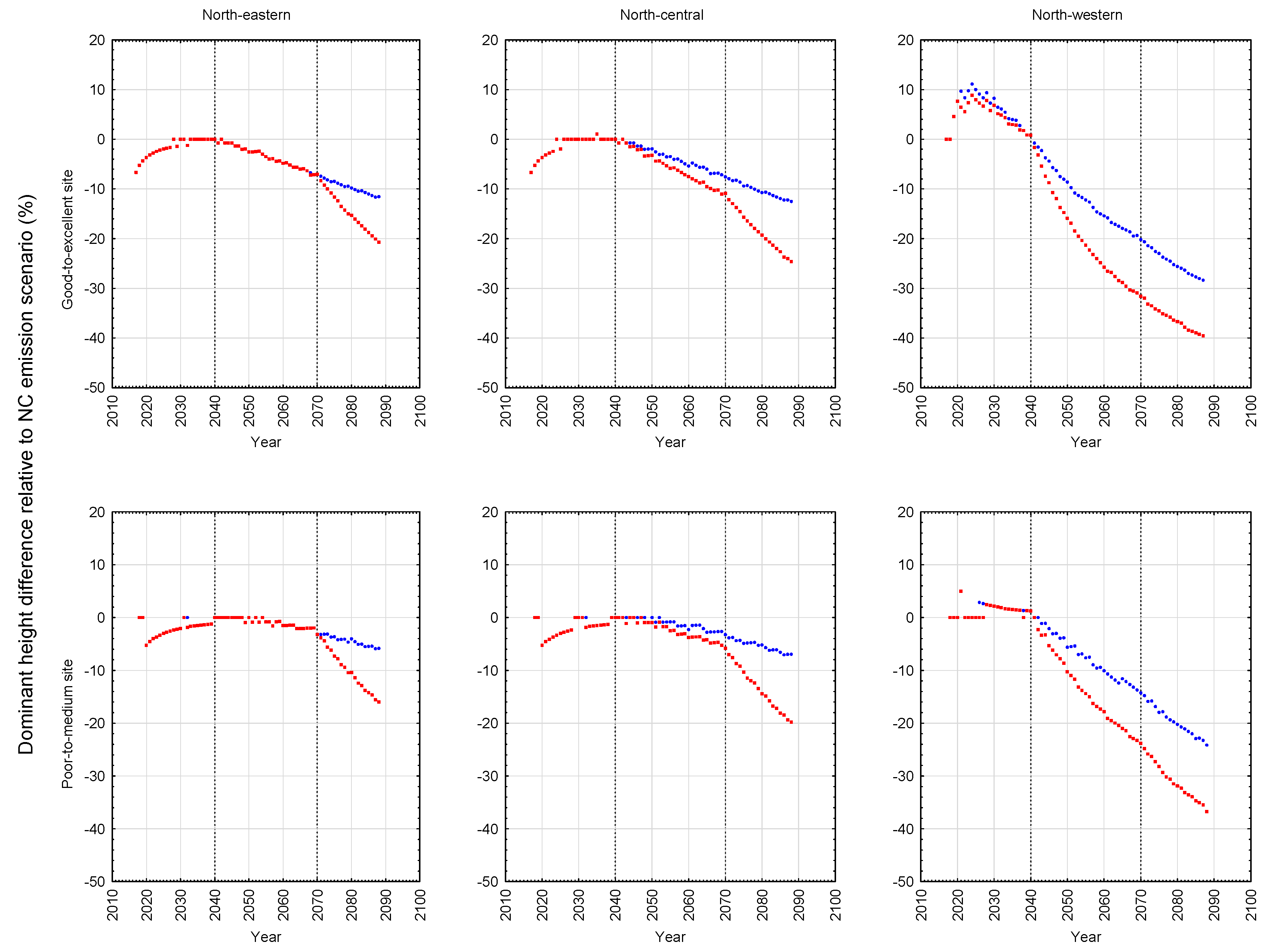
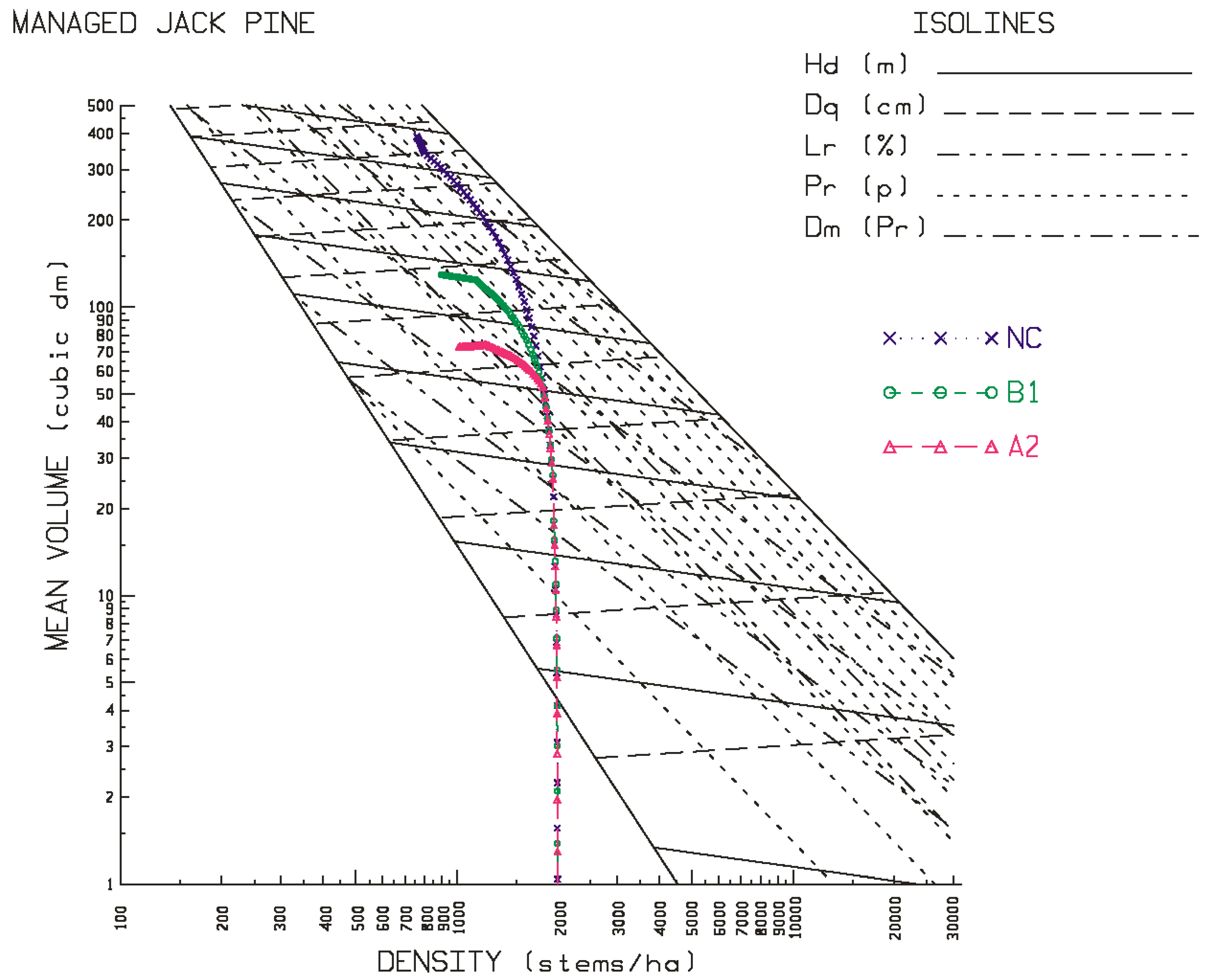
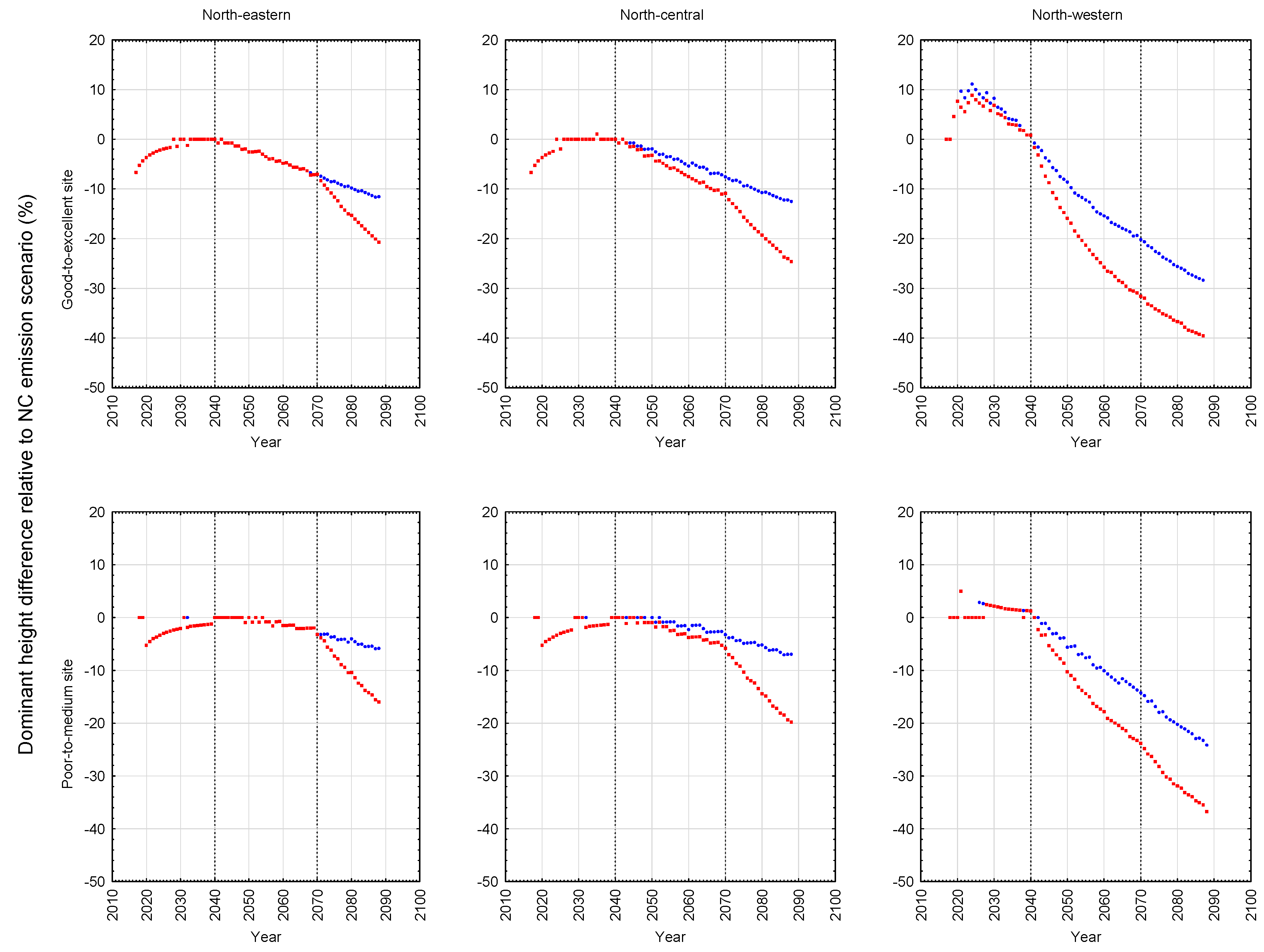
Contrary to the site-specific response patterns observed for black spruce, jack pine exhibited a consistent and orderly pattern of productivity declines on both site qualities (good-to-excellent > poor-to-medium) at all three locations (north-western > north-central > north-eastern) under both emission scenarios (A2 > B1). More specifically, yield reductions were evidenced in rotational mean sizes (quadratic mean diameters and mean volumes), volumetric, biomass and carbon yields, and end-products (increased production of pulp logs, decreased production of sawlogs, and reductions in chip and lumber recovery volumes irrespective of mill-type). Rotational site occupancy was also reduced as measured by basal area and relative density. Performance indices also indicated declining merchantable volumetric, biomass and carbon productivity, sawlog and lumber production, and economic worth. The negative effect of the A2 emission scenario was approximately 1.5 times greater than that of the B1 emission scenario. There was also a slight east-to-west increasing trend in the degree of the reductions. Although differences in responses in terms of stand stability, wood quality and time to operability status were evident among the emission scenarios, they were frequently negative and (or) largely inconsequential.
Although speculative, one plausible expectation for the negativity of the jack pine responses is that the additional moisture expected under climate change may not be made available given the low moisture retention ability of the sandy soils which jack pine traditionally occupies. Additionally, temperature-induced increased rates of evapotranspiration and tree respiration [
25] during the growing season may divert some of the photosynthate away from the production of new plant tissue. Inferences derived from physiological-based tree models have reported adverse climate change effects on jack pine arising from hydraulic limitations generated from temperature-induced vapour pressure deficits [
26]. Similar site-specific effects were reported by Loustau et al. [
27] who found that climate effects on the productivity of forests in western Europe were greatest on the higher fertility sites. The results of these simulations suggest that jack pine will not fare well under a changing climate and declines in rotational yields in terms of mean sizes, total and merchantable volume, biomass and carbon yields, end-products, and economic worth, may be the plausible consequences of wetter and warmer growing seasons. Conceptually, the growth response of a given species to climate change is largely a function of its ability to adapt to changing environmental conditions. Depending on the range of a species’ ecological amplitude, responses may be negative or positive [
28]. Although speculative, the results of this study suggest that black spruce may have a greater ability to adjust to a changing climate than does jack pine.
4.2. Modelling Approach and Limitations
As demonstrated in this study, for a given crop plan, site quality, set of climate variables specific to geographic locale, emission scenario, and commitment period, the modified SSDMMs were able to predict the consequences of climate change in terms of future productivity (e.g., mean annual volume, biomass and carbon increments), volumetric yields (e.g., total and merchantable volumes per unit area), log-product distributions (e.g., number of pulp and saw logs), biomass production and carbon sequestration outcomes (e.g., oven-dried masses of above-ground components and associated carbon equivalents), recoverable end-products and associated monetary values (e.g., volume and economic value of recovered chip and dimensional lumber products) by sawmill-type (stud and randomized length processing protocols), economic efficiency (e.g., land expectation value), structural stability, and fibre attributes (e.g., wood density). Most of these metrics and performance measures did vary by emission scenario, site quality, locale, and species with the exception of stand stability and wood density.
The SSDMMs do address suppression-related density-dependent mortality processes and density-independent mortality (governed by the operational adjustment factor inputted during the model initialization phase (e.g.,
Table 2)). However, climate change effects on survival probabilities are not explicitly accounted for in the current model formulations and thus mortality estimates are likely under-estimated. Conversely, any growth increases arising from increased carbon dioxide concentrations (CO
2 fertilization) are also not accounted for. Further research on the effects of a changing climate on a broader array of fibre attributes, and mortality and growth processes would have utility in informing future modelling efforts. Although this study assessed a single crop plan that was consistent with current operational silvicultural practice in the central portion of the Canadian Boreal Forest Region, the modified SSDMMs are fully capable of simulating a multitude of different crop plans including those that encompass thinning events for any nominal rotation length, site quality, or locale.
Employing the results from a variety of different models has been useful in delineating a range of plausible outcomes of climate change on boreal ecosystems (e.g., [
24,
29,
30,
31,
32,
33]). However, the availability of species-specific models which account for site variability and geographic location and have the ability to evaluate outcomes in terms of stand-level productivity metrics applicable to the forest management community, has been limited. The results of this study provide additional support for the model adaptation approach: i.e., modifying existing stand-level forecasting models and systems in order to estimate site-specific effects of climate change and thereby conserving outcome metrics which are applicable to operational forest management decision-making. More specifically, the site-based dominant height-age functions which govern the temporal rate of stand development and structural change within the SSDMM structure [
5,
6] are key relationships for incorporating external influences. Replacing conventional site-specific height-age equations with their biophysical equivalents that account for climatic influences through the introduction of climate-based predictor variables, enabled the evaluation of climate change effects at the stand-level by emission scenario, site quality, locale, and species. Given the similarities between the SSDMM structure and numerous other stand-level growth and yield forecasting systems with respect to their employment of site-specific height-age models, suggests that the biophysical functional approach may have broader applicability when attempting to modify existing models in order to account for climate change effects.
Based on the projection of climate change effects in terms of mean temperature and precipitation rates, the simulations used input values that extended beyond the range that were used to parameterize the biophysical models. Specifically, the biophysical equations were parameterized using data derived from historical or current climate measurements. Hence, similar to other models that attempt to project the effects of the future climate, the biophysical models are being used outside of the range of their underlying calibration data sets. Reviewing the values given in
Table 1 of Sharma and others [
16], indicates that the precipitation and temperature values for their sample sites ranged from a minimum of 403 mm to a maximum of 502 mm, and from 12.0 to 13.4 °C, respectively. Contrasting these values with those given in
Table 1 (490 to 577 mm for precipitation and 12.5 to 17.0 °C for temperature), suggest that there is a significant risk of projection error given the power exponent’s functional dependence on these variables (Equations (1) and (2)). Although employing the difference approach in terms of height accumulation estimates across the commitment periods assisted in minimizing these non-linear effects in addition to providing a continuous and logical height-based response curve, the degree of prediction error is largely unknown.
The relative differences of these predicted height development patterns for the B1 and A2 emission scenarios with respect to the NC emission scenario are graphically illustrated for black spruce plantations in
Figure 5 and jack pine plantations in
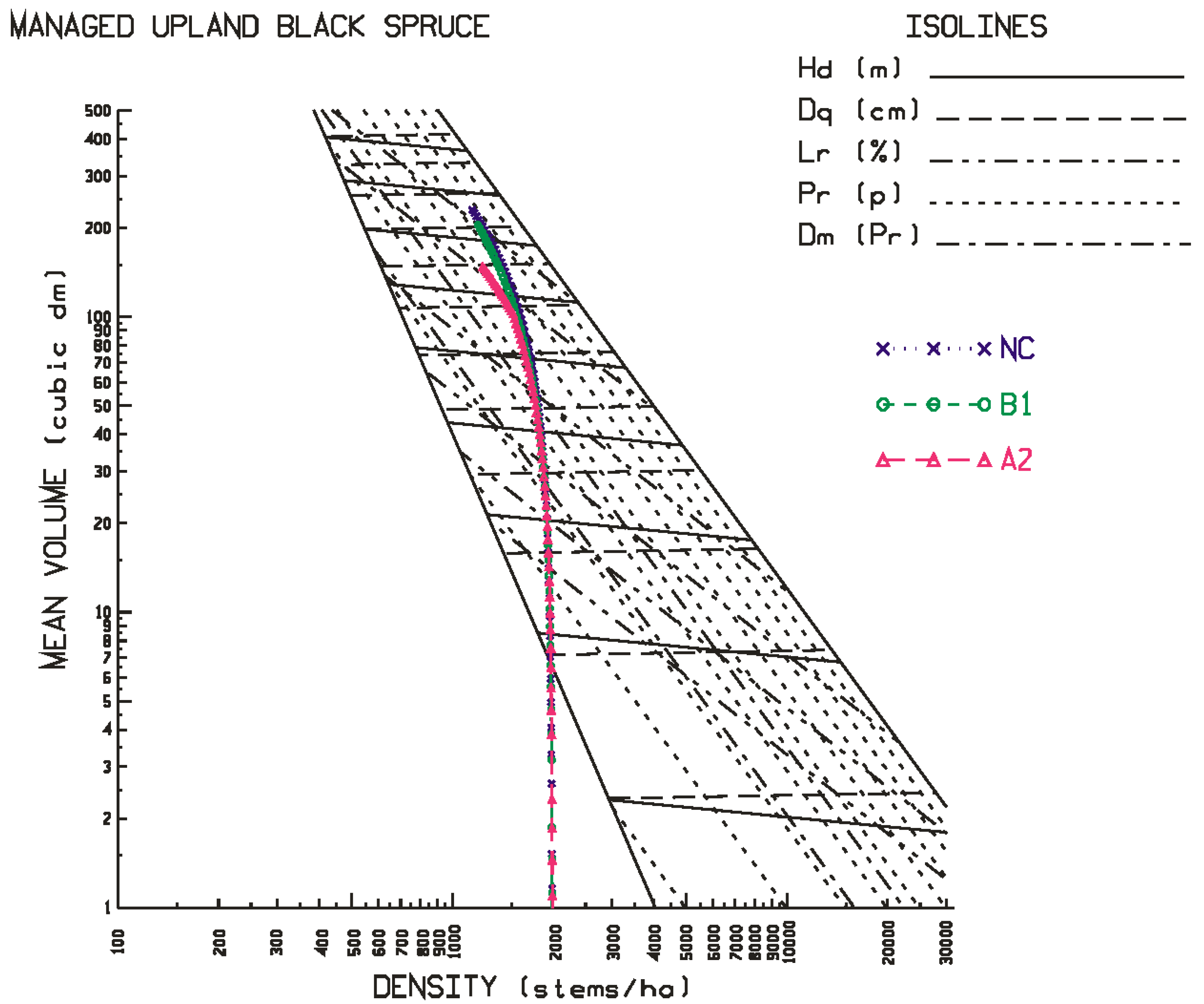
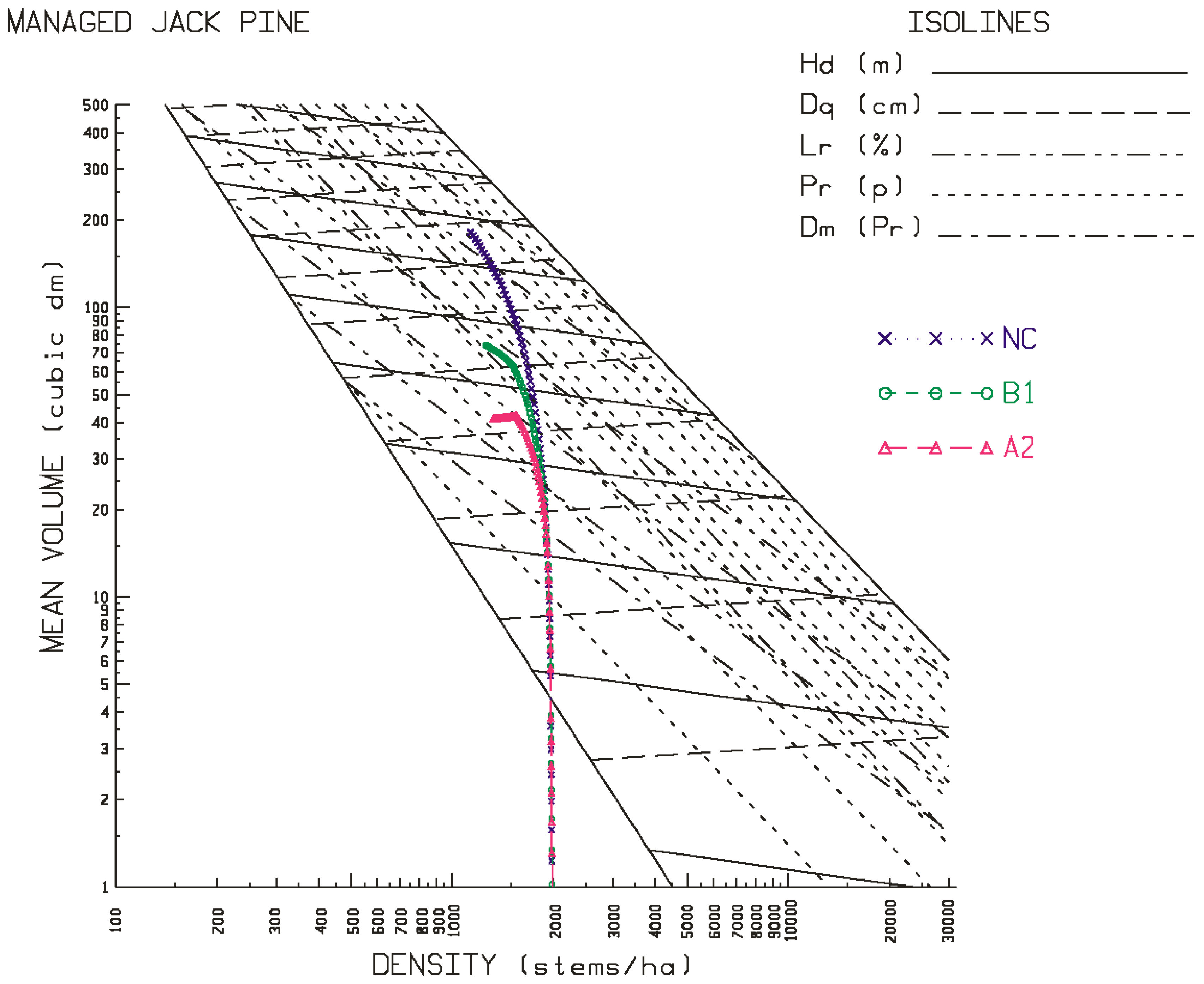
Figure 6 for the site qualities and locales examined in this study. As evident in the temporal patterns for black spruce, most of the positive and negative effects of climate change were initiated at the beginning of the second commitment period. Similar trends were evident for jack pine with the exception that the relative height growth reductions were greater and rate of decline accelerated during the third commitment period with respect to the A2 emission scenario. These response patterns were also consistent with the size-density trajectories illustrated in the SSDMM graphics (e.g.,
Figure 1 and
Figure 2 for black spruce and
Figure 3 and
Figure 4 for jack pine) and their rotational consequences in terms of the productivity measures and performance metrics as presented in
Table 3a,
Table 3b and
Table 3c for black spruce and
Table 4a,
Table 4b and
Table 4c for jack pine: e.g., the greater the relative reduction in dominant height growth of the B1 and A2 scenarios (
Figure 5 and
Figure 6), the lower the corresponding plantation is positioned within the size-density space at rotation (
Figure 1 and
Figure 2 for black spruce and
Figure 3 and
Figure 4 for jack pine), and the greater the reduction in rotational productivity outcomes (
Table 3a,
Table 3b and
Table 3c for black spruce and
Table 4a,
Table 4b and
Table 4c for jack pine).
Similar approaches have been used to predict the effects of climate change for other forest tree species: e.g., lodgepole pine (
Pinus contorta Douglas ex Louden) in western North America [
34], and Norway spruce (
Picea abies (L.) Karst.) and common beech (
Fagus sylvatica L.) in central Europe [
32]. The employment of the comprehensive modelling framework represented by the SSDMM conceptually parallels the previous efforts where large integrated decision-support models have been modified to account for climate change using a similar approach (e.g., FVS [
8] and TASS [
9]). Additionally, more elaborate biophysical models which directly account for GHG concentrations have also been developed and incorporated into stand-level decision-support systems (e.g., SILVA [
7]). With respect to the modified SSDMMs presented in this study, Sharma and others [
16] evaluated the significance of including additional climate variables into the biophysical height-age model specifications and concluded that adding variables such as soil-moisture related variables, would not significantly improve the explanatory performance of their models (e.g., final model forms explained over 98% of the variation in the dependent variables).
In a more general sense, these statistical quasi-causal approaches (e.g., [
28]) do not afford the ability to gain insight nor understanding of the underlying ecophysiological effects of climate change on tree and stand growth processes, as do process-based models. However, these mensurational-based approaches can empirically account for climate change effects by providing a plausible range of outcomes in metrics that are of utility in forest management planning and policy formulation. As demonstrated, the decline in the rate of stand development arising from deteriorating growing conditions under climate change translates into growth reductions and productivity declines at rotation. Although survival rates were largely unaffected due to the establishment year employed (2014), plantations established in later years may very well experience failure due to climate-driven density-independent factors which could negatively affect seedling survival. Consequently, species substitution and assisted migration (population migration, range expansion, and long-distance migration [
35]) are plausible silvicultural considerations when reforesting sites throughout the central portion of the Canadian Boreal Forest Region [
11].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}