
Previous Land Use and Invasive Species Impacts on Long-term Afforestation Success

Abstract
:1. Introduction
2. Experimental Section
2.1. Research Area
2.2. Sampling
2.3. Statistical Analysis
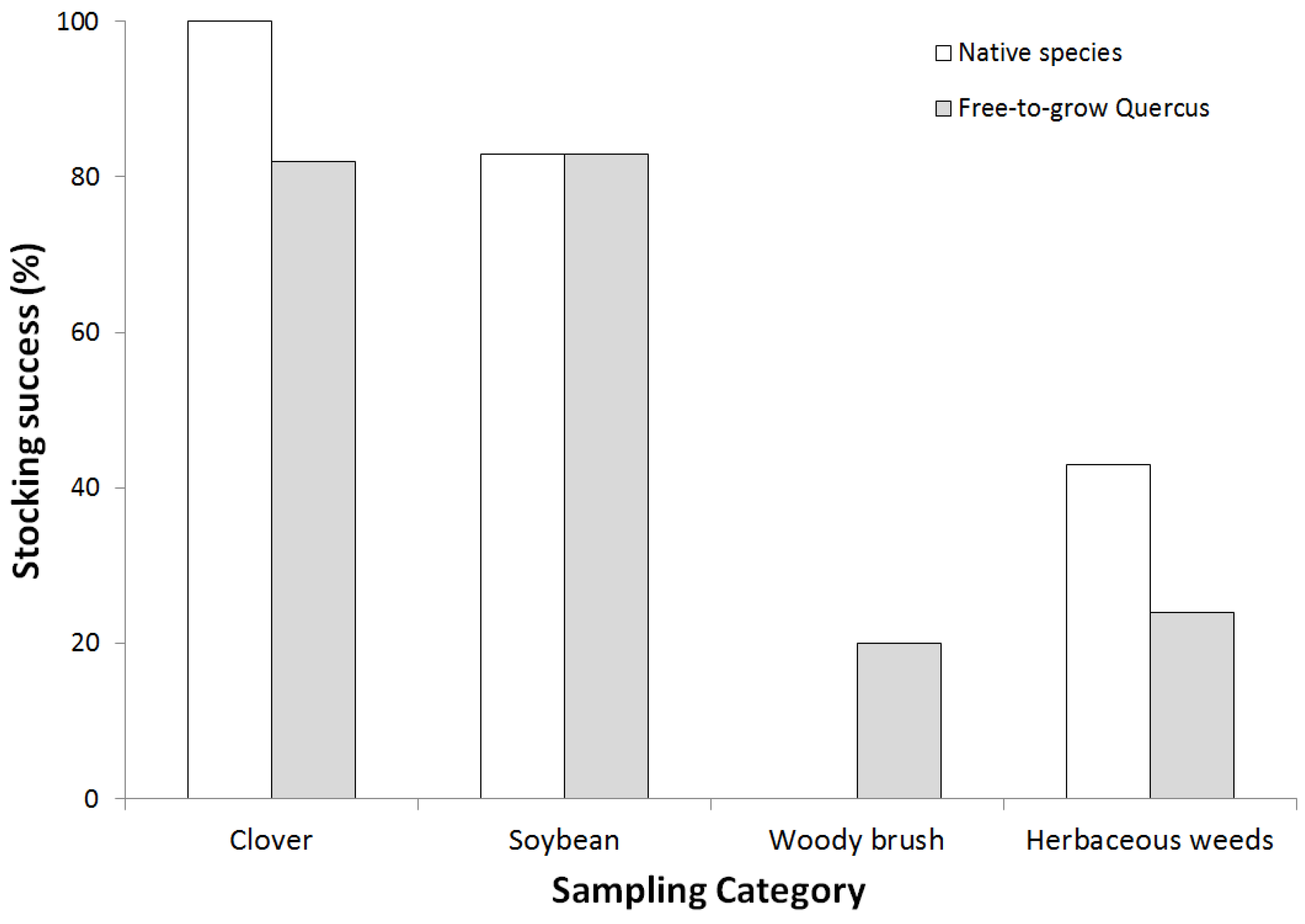
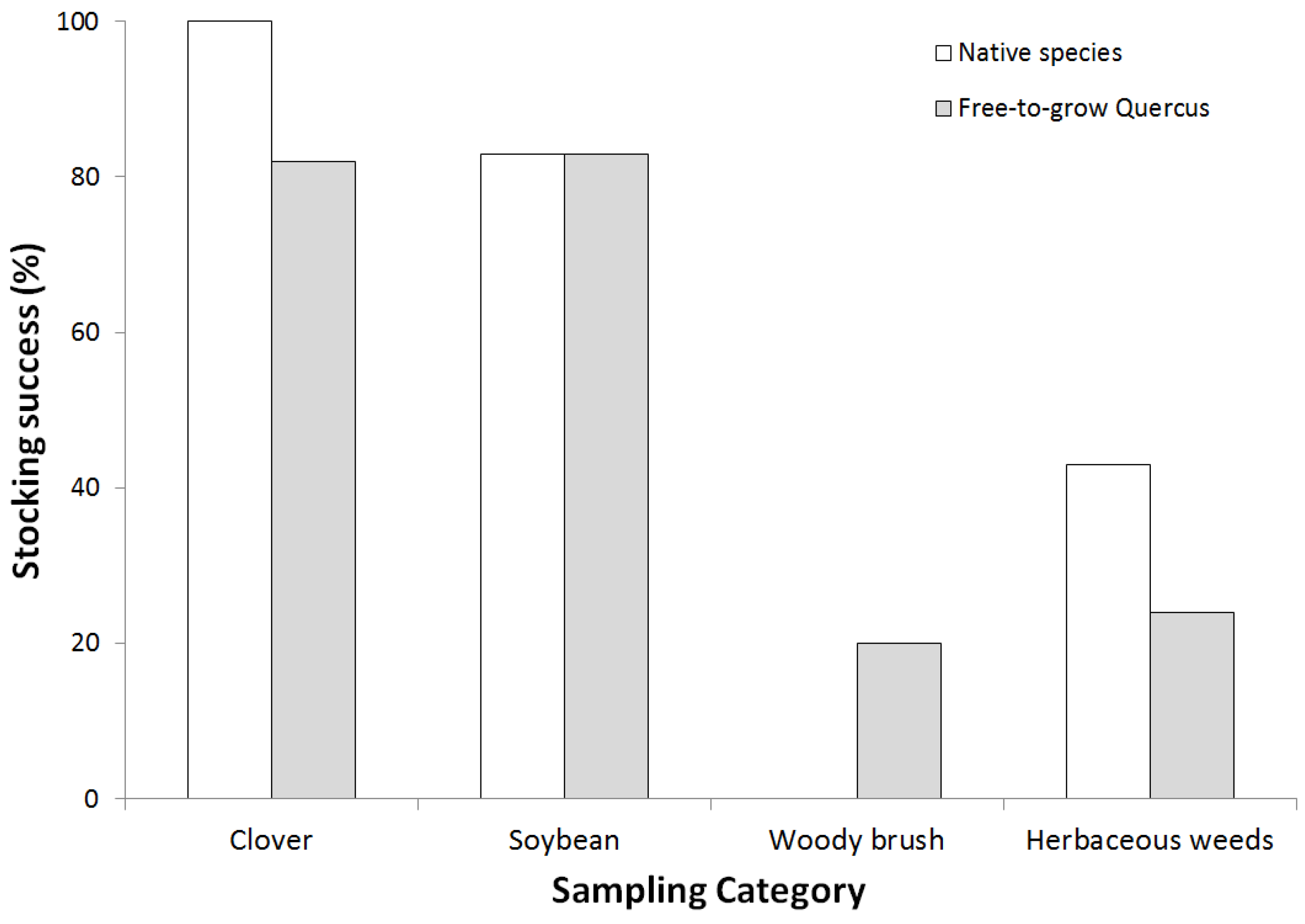
2.4. Afforestation Success
3. Results
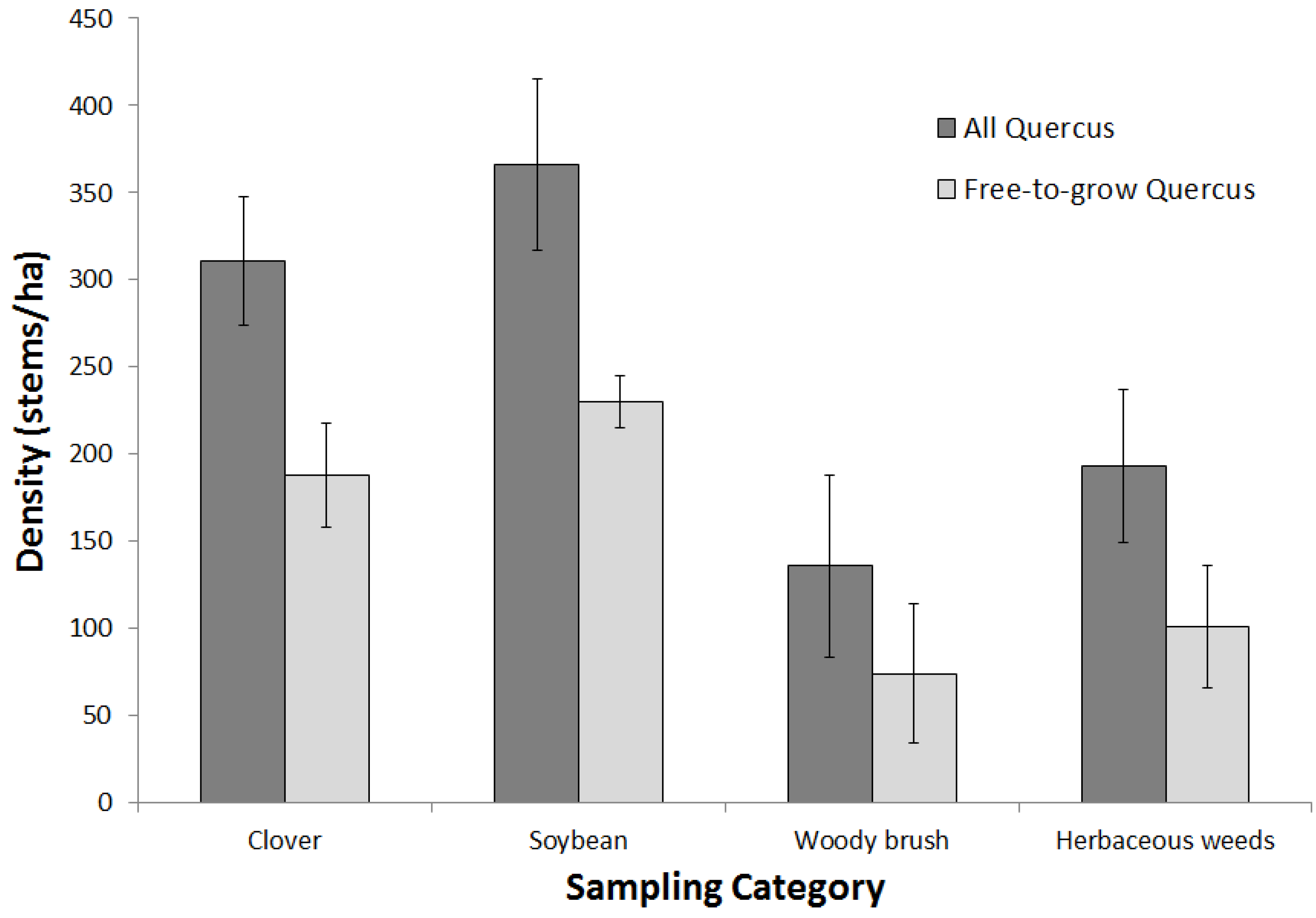
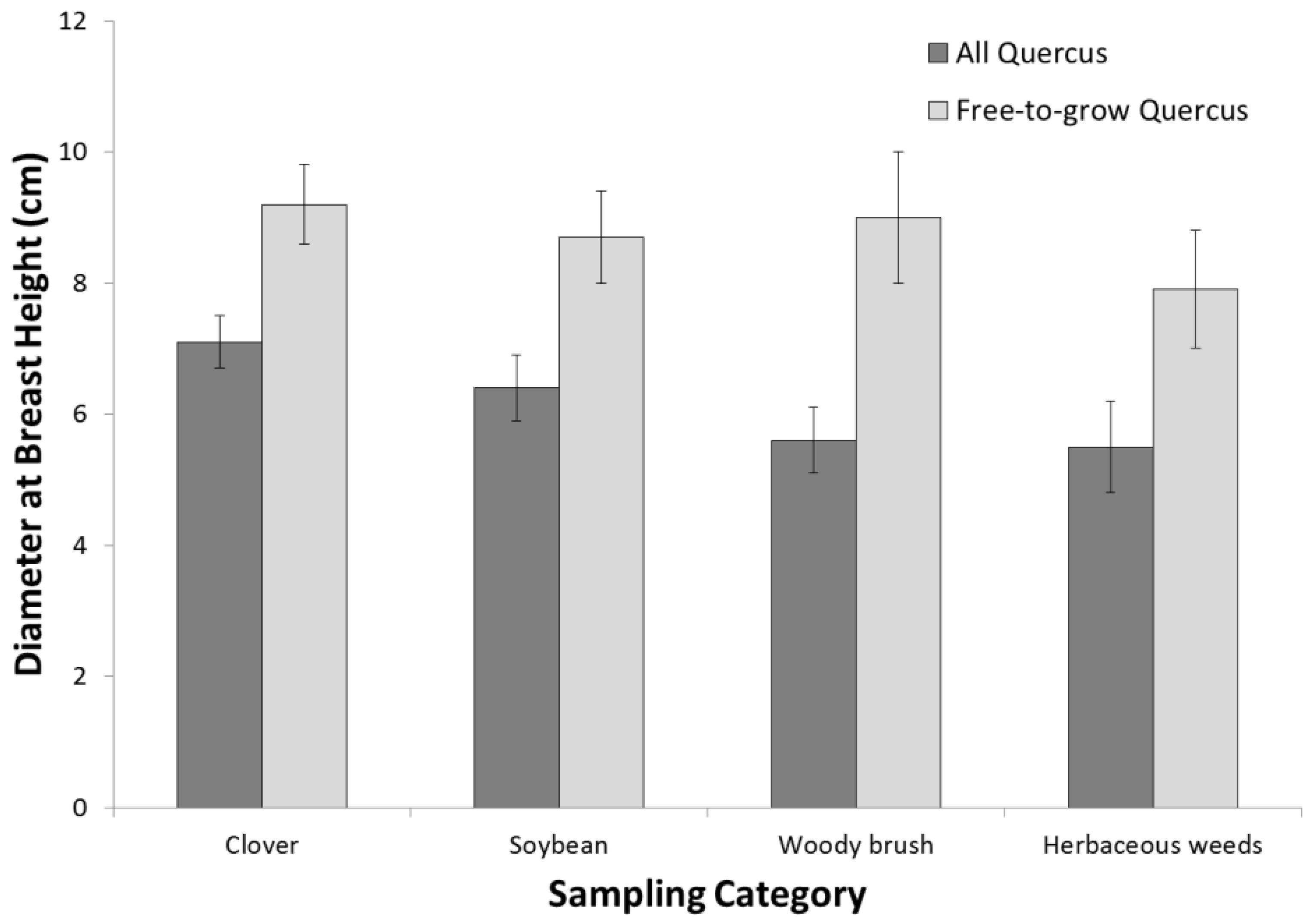
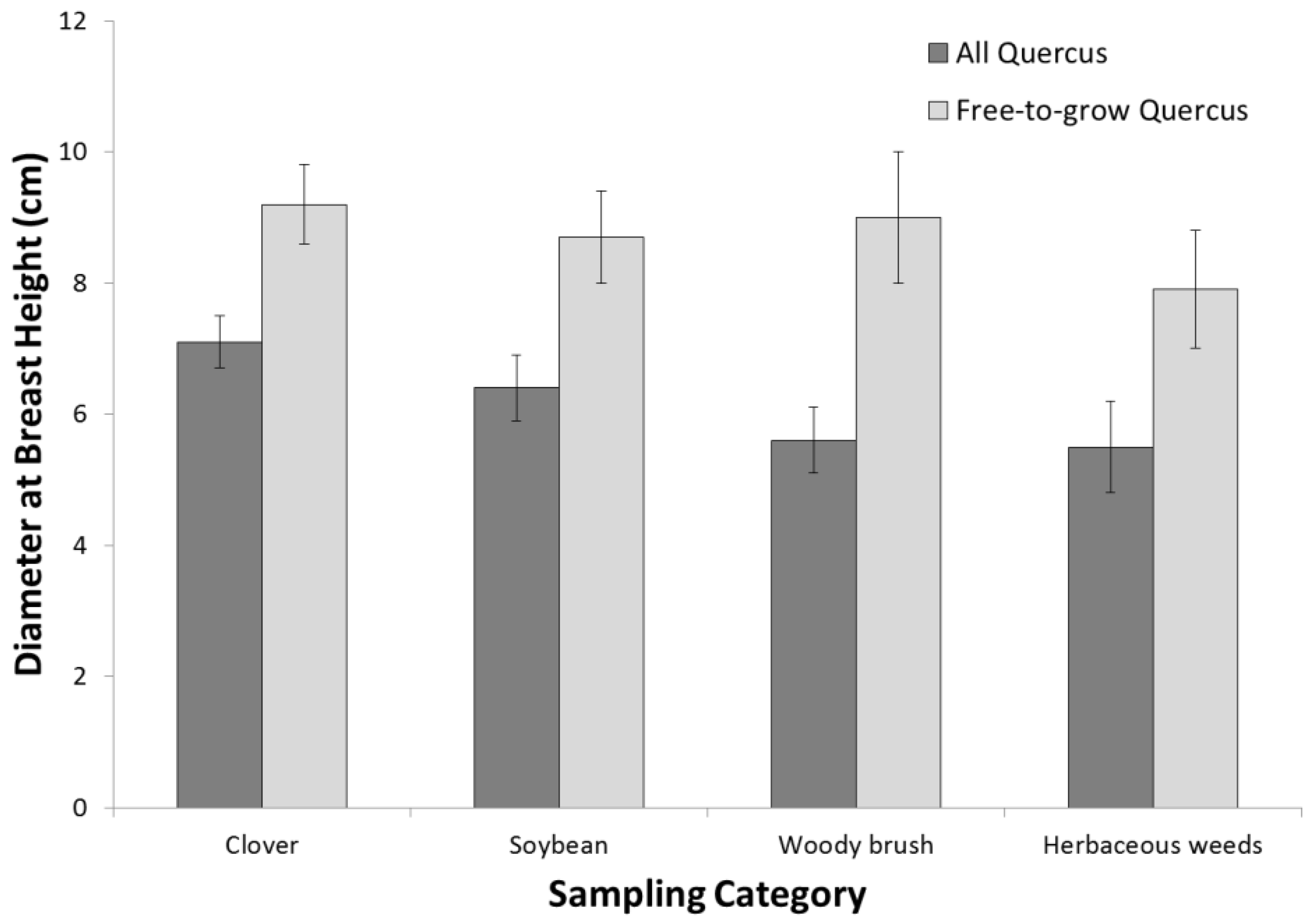
3.1. Tree Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | <2.5 cm | 2.5–7.6 cm | 7.6–12.7 cm | >12.7 cm | Total trees sampled | # of sites occupied |
|---|---|---|---|---|---|---|
| Q. alba | 20 | 43 | 27 | 9 | 411 | 26 |
| Q. palustris | 11 | 40 | 31 | 17 | 252 | 22 |
| Q. macrocarpa | 39 | 45 | 15 | 2 | 137 | 23 |
| Q. rubra | 13 | 52 | 28 | 7 | 127 | 13 |
| Q. bicolor | 3 | 26 | 40 | 30 | 99 | 8 |
| Q. velutina | 29 | 32 | 25 | 14 | 72 | 14 |
| Q. shumardii | 11 | 68 | 16 | 5 | 38 | 3 |
| Q. imbricaria | 42 | 32 | 26 | 0 | 31 | 10 |
| Q. muehlenbergii | 27 | 27 | 27 | 18 | 11 | 3 |
| Q. acutissima | 25 | 0 | 25 | 50 | 4 | 1 |
| Q. michauxii | 0 | 100 | 0 | 0 | 1 | 1 |
| Total | 19 | 42 | 27 | 12 | 1,183 | 28 |

| Species | <2.5 cm | 2.5–7.6 cm | 7.6–12.7 cm | >12.7 cm |
|---|---|---|---|---|
| Acer negundo | 3 | 3 | 6 | 4 |
| Acer rubrum | 9 | 13 | 9 | 3 |
| Diospyros virginiana | 2 | 3 | 7 | 0 |
| Fraxinus spp. | 16 | 17 | 11 | 4 |
| Juniperus virginiana | 0 | 6 | 9 | 4 |
| Liquidambar styraciflua | 6 | 8 | 20 | 36 |
| Rhus copallina | 3 | 5 | 0 | 0 |
| Ulmus alata | 5 | 4 | 0 | 0 |
| Ulmus americana | 49 | 31 | 18 | 11 |
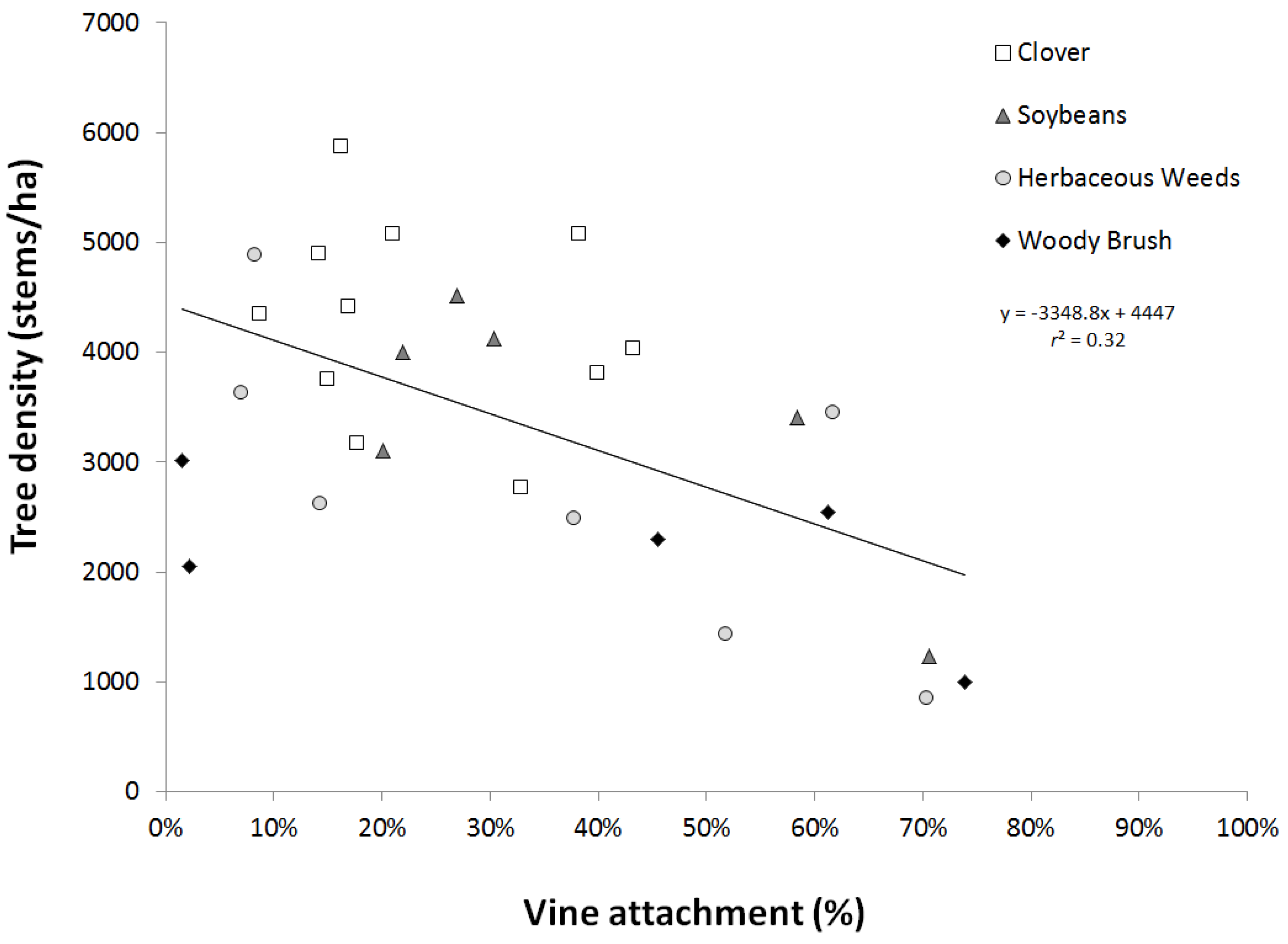
3.2. Afforestation Success
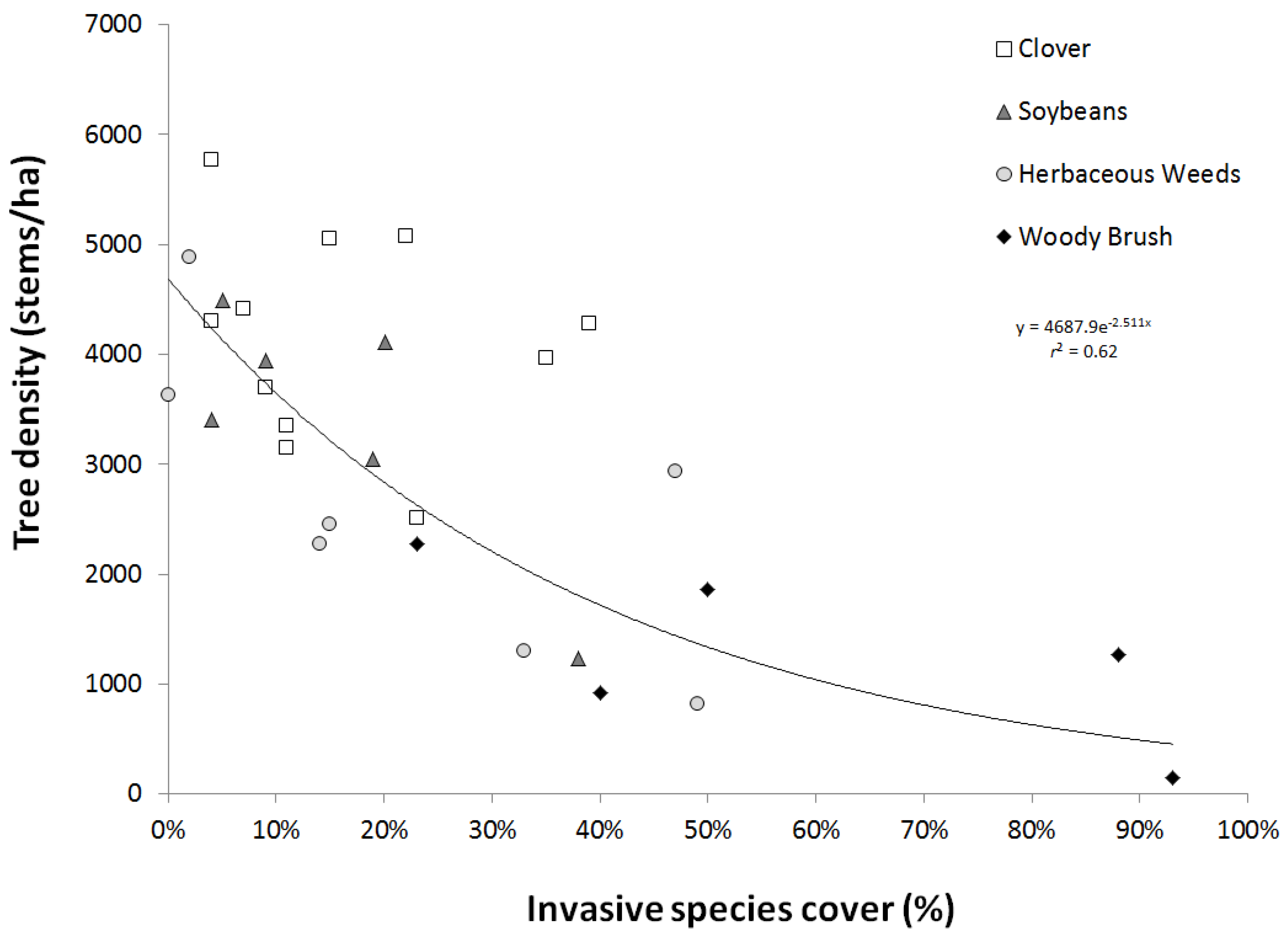
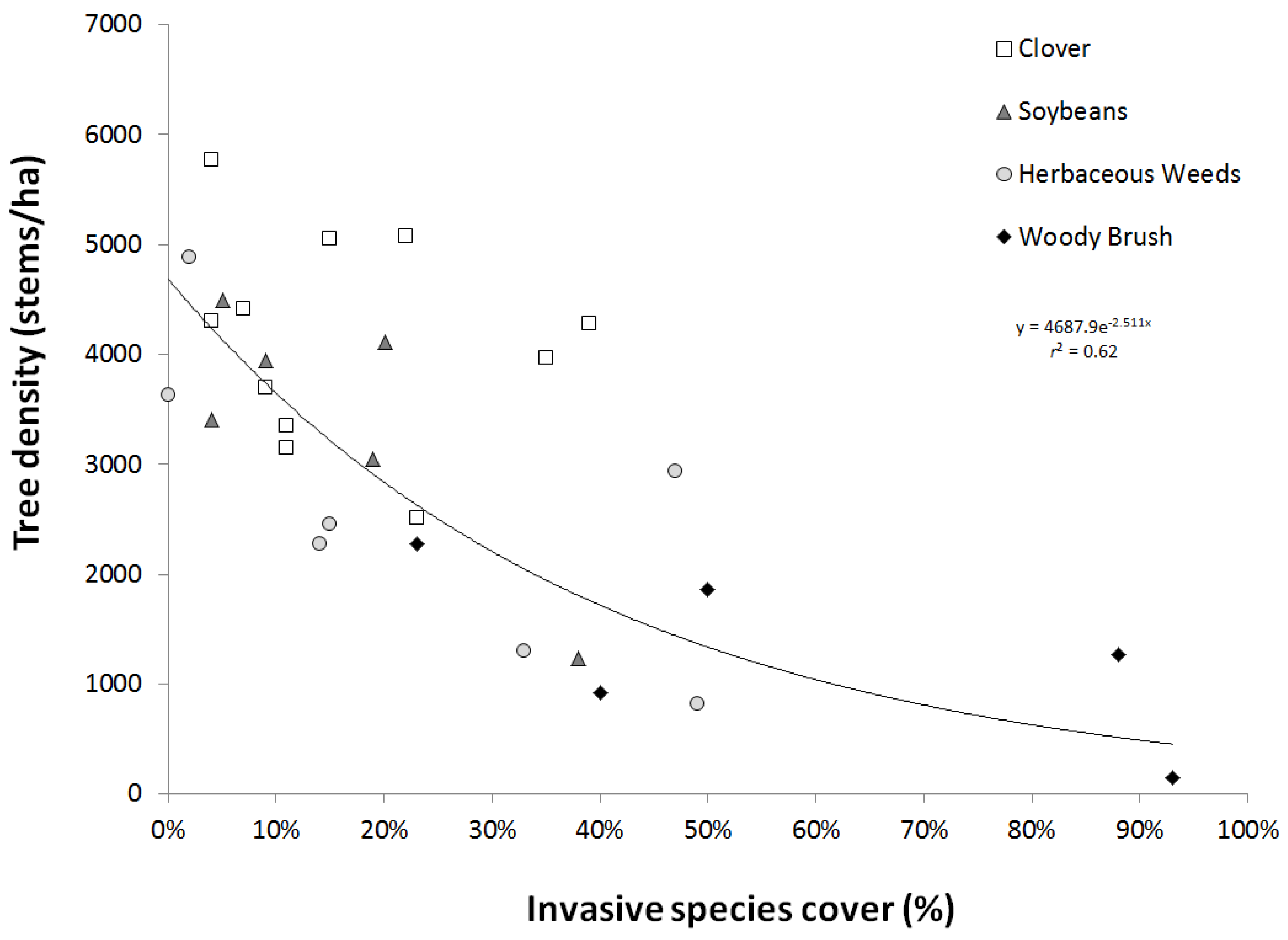
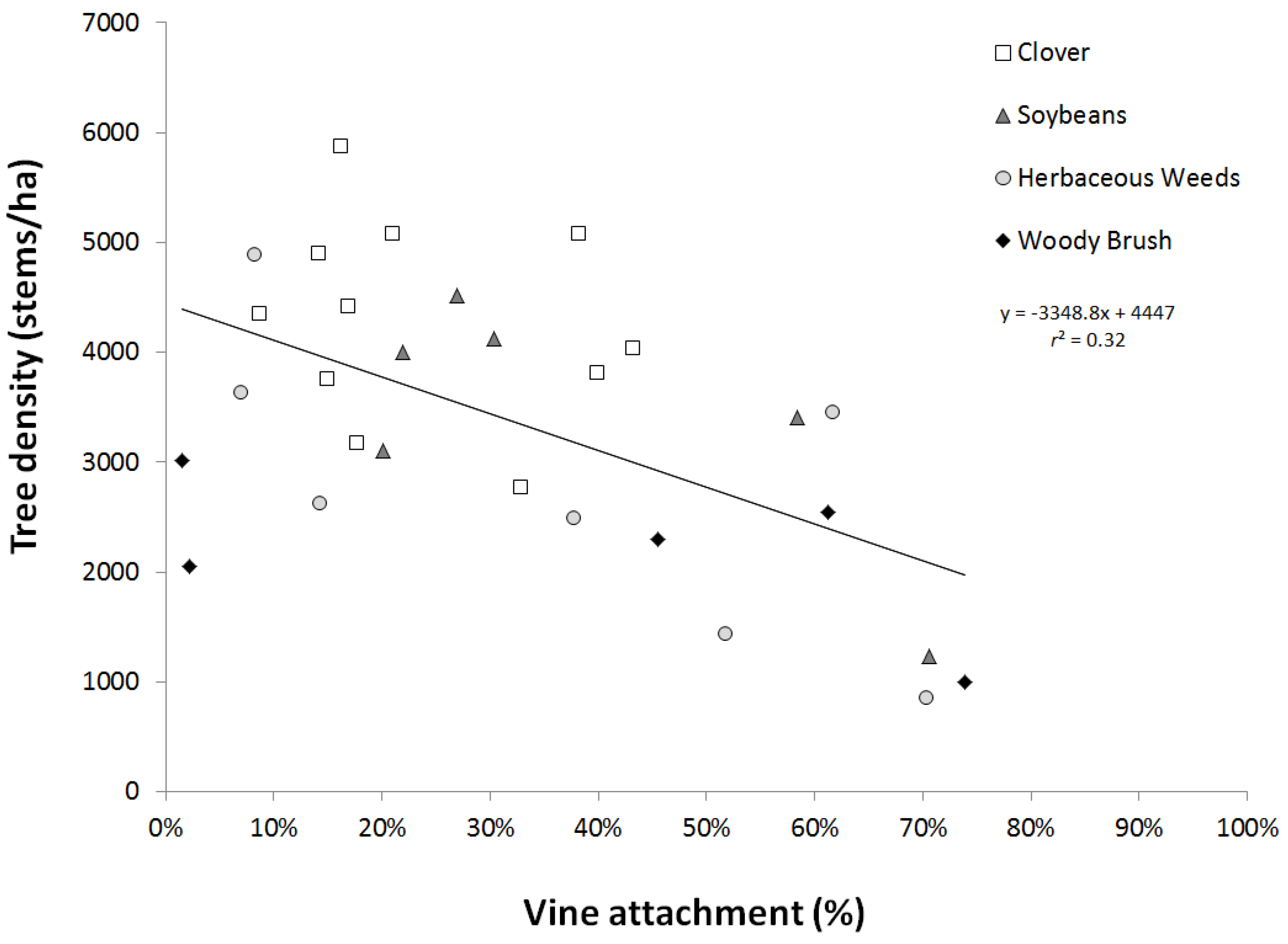
3.3. Invasive Species
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Vesterdal, L.; Ritter, E.; Gundersen, P. Change in soil organic carbon following afforestation of former arable land. For. Ecol. Manag. 2002, 169, 137–147. [Google Scholar] [CrossRef]
- Davis, A.S.; Jacobs, D.F. Afforestation in the central hardwood forest region of the USA. In The Thin Green Line: A Symposium on the State-of-the-Art in Reforestation, In Proceedings of Ontario Ministry of Natural Resources, Thunder Bay, ON, Canada, 2005; Colombo, S.J., Ed.; pp. 48–53.
- Xu, J.; Yin, R.; Li, Z.; Liu, C. China’s ecological rehabilitation: Unprecedented efforts, dramatic impacts, and requisite policies. Ecol. Econ. 2006, 57, 595–607. [Google Scholar] [CrossRef]
- Ramankutty, N.; Foley, J.A. Estimating historical changes in global land cover: Croplands from 1700 to 1992. Global Biochem. Cy. 1999, 13, 997–1027. [Google Scholar] [CrossRef]
- Benayas, J.R.; Martins, A.; Nicolau, J.M.; Schulz, J.J. Abandonment of agricultural land: An overview of drivers and consequences. CAB Rev. Perspect. Ag. Vet. Sci. Nutrit. Nat. Resour. 2007, 2, 1–14. [Google Scholar] [CrossRef]
- Ross-Davis, A.L.; Broussard, S.R.; Jacobs, D.F.; Davis, A.S. Afforestation motivations of private landowners: An examination of hardwood tree plantings in Indiana. North. J. Appl. For. 2005, 22, 149–153. [Google Scholar]
- Lima, A.M.; Silva, I.R.; Neves, J.C.; Novais, R.F.; Barros, N.F.; Mendonça, E.S.; Smyth, T.J.; Moreiraa, M.S.; Leite, F.P. Soil organic carbon dynamics following afforestation of degraded pastures with eucalyptus in southeastern Brazil. For. Ecol. Manag. 2006, 235, 219–231. [Google Scholar] [CrossRef]
- Niu, X.; Duiker, S.W. Carbon sequestration potential by afforestation of marginal agricultural land in the Midwestern US. For. Ecol. Manag. 2006, 223, 415–427. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Gaskins, M.D.; Mangun, J.C. A GIS approach to prioritizing habitat for restoration using Neotropical migrant songbird criteria. Environ. Manag. 2011, 48, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Fei, S.; Kong, N.; Steiner, K.C.; Moser, W.K.; Steiner, E.B. Change in oak abundance in the eastern United States from 1980 to 2008. For. Ecol. Manag. 2011, 262, 1370–1377. [Google Scholar] [CrossRef]
- Dey, D.C. Sustaining oak forests in eastern North America: Regeneration and recruitment, the pillars of sustainability. For. Sci. 2014, 60, 926–942. [Google Scholar] [CrossRef]
- Delate, K.; Holzmueller, E.; Davis Frederick, D.; Mize, C.; Brummer, C. Tree establishment and growth using forage ground covers in an alley-cropped system in Midwestern USA. Agroforest. Syst. 2005, 65, 3–52. [Google Scholar] [CrossRef]
- Dey, D.C.; Jacobs, D.; McNabb, K.; Miller, G.; Baldwin, V.; Foster, G. Artificial regeneration of major oak (Quercus) species in eastern United States—A review of the literature. For. Sci. 2008, 54, 77–106. [Google Scholar]
- Jacobs, D.F.; Ross-Davis, A.L.; Davis, A.S. Establishment success of conservation tree plantations in relation to silvicultural practices in Indiana, USA. New For. 2004, 28, 23–26. [Google Scholar] [CrossRef]
- Ruzicka, K.J.; Groninger, J.W.; Zaczek, J.J. Deer browsing, forest edge effects, and vegetation dynamics following bottomland forest restoration. Restor. Ecol. 2010, 18, 702–710. [Google Scholar] [CrossRef]
- Ceacero, C.J.; Díaz-Hernández, J.L.; del Campo, A.D.; Navarro-Cerrillo, R.M. Interactions between soil gravel content and neighboring vegetation control management in oak seedling establishment success in Mediterranean environments. For. Ecol. Manag. 2012, 271, 10–18. [Google Scholar] [CrossRef]
- Olarieta, J.R.; Rodríguez-Ochoa, R.; Ascaso, E. Soil gypsum and increased penetration resistance restrict early growth of Quercus ilex plantations. Arid Land Res. Manag. 2012, 26, 250–260. [Google Scholar] [CrossRef]
- Ebinger, J.E. Exotic shrubs: A potential problem in natural area management in Illinois. Nat. Area. J. 1983, 3, 3–6. [Google Scholar]
- Robertson, D.J. Trees, deer and non-native vines: Two decades of northern piedmont forest restoration. Ecolog. Restor. 2012, 30, 59–70. [Google Scholar] [CrossRef]
- Holzmueller, E.J.; Jose, S. Invasive plant conundrum: What makes the aliens so successful? J. Trop. Ag. 2009, 47, 18–29. [Google Scholar]
- Crab Orchard National Wildlife Refuge. Affected Environment. In Comprehensive Conservation Plan; Crab Orchard National Wildlife Refuge US FWS: Marion, IL, USA, 2005; pp. 120–168. [Google Scholar]
- Palmer, T.A. Reforestation Plan; Crab Orchard National Wildlife Refuge US FWS: Marion, IL USA, 1995; pp. 1–6. [Google Scholar]
- SAS SAS/STAT User Guide, Version 9.3, SAS Institute: Cary, NC, USA, 2011.
- Gingrich, S.F. Measuring and evaluation stocking and stand density in upland hardwood forests in the central states. For. Sci. 1967, 13, 38–53. [Google Scholar]
- Minckler, L.S.; Boggess, W.R.; Backler, W.R.; Campen, E.R.; Kladiva, J.; Culver, L.B.; Kurmes, E.A.; Fox, H.W.; Menzle, J.F.; Massie, G.E.; et al. Recommended Silviculture and Management Practices for Illinois Hardwood Forest Types; Illinois Technical Forestry Association: Springfield, IL, USA, 1972; pp. 1–53. [Google Scholar]
- Hosner, J.F.; Minckler, L.S. Bottomland hardwood forest of southern Illinois: Regeneration and succession. Ecology 1963, 44, 29–41. [Google Scholar] [CrossRef]
- Kruse, B.S.; Groninger, J.W. Vegetative Characteristics of Recently Reforested Bottomlands in the Lower Cache River Watershed, Illinois, USA. Restor. Ecol. 2003, 11, 273–280. [Google Scholar] [CrossRef]
- Clatterbuck, W.K.; Hodges, J.D. Development of cherrybarkoak and sweetgum in mixed, evenaged bottomland stands in central Mississippi, USA. Can. J. For. Res. 1988, 18, 12–18. [Google Scholar] [CrossRef]
- Lockhart, B.R.; Gardiner, E.; Leininger, T.; Stanturf, J. A stand-development approach to oak afforestation in the Lower Mississippi Alluvial Valley. South. J. Appl. For. 2008, 32, 120–129. [Google Scholar]
- Catling, P.M.; Oldham, M.J.; Sutherland, D.A.; Brownell, V.R.; Larson, B.M.H. The recent spread of autumn-olive, Elaeagnus umbellata, into Southern Ontario and its current status. Can. Field Nat. 1997, 111, 376–380. [Google Scholar]
- Meiners, S.J. Apparent competition: An impact of exotic shrub invasion on tree regeneration. Biol. Invas. 2007, 9, 849–855. [Google Scholar] [CrossRef]
- Moore, M.R.; Buckley, D.S.; Klingeman, W.E., III; Saxton, A.M. Distribution and growth of autumn olive in managed forest landscape. For. Ecol. Manag. 2013, 310, 589–599. [Google Scholar] [CrossRef]
- Evans, D.M.; Zipper, C.E.; Burger, J.A.; Strahm, B.D.; Villamagna, A.M. Reforestation practice for enhancement of ecosystem services on a compacted surface mine: Path toward ecosystem recovery. Ecol. Engin. 2013, 51, 16–23. [Google Scholar] [CrossRef]
- Blandier, P.; Frochot, H.; Sourisseau, A. Improvement of direct tree seeding with cover crops in afforestation: Microclimate and resource availability induced by vegetation composition. For. Ecol. Manag. 2009, 257, 1716–1724. [Google Scholar] [CrossRef]
- Gardiner, E.S.; Russell, D.R.; Oliver, M.; Dorris, L.C., Jr. Bottomland hardwood afforestation: State of the art. In Proceedings of a Conference on Sustainability of Wetlands and Water Resources; Holland, M.M., Warren, M.L., Stanturf, J.A., Eds.; Oxford: Stoneville, MS, USA, 2002; pp. 75–86. [Google Scholar]
- Bobsin, K.; Conn, W.; Deizman, P.; Edgington, J.; Groninger, J.; Hayek, J.; Sipp, S. We Planted All These Trees, Now What? A Field Guide for Tree Care; Illinois Forestry Association: Chatham, IL, USA, 2014; pp. 1–17. [Google Scholar]
- McLane, C.R.; Battaglia, L.L.; Gibson, D.J.; Groninger, J.W. Succession of exotic and native species assemblages across a chronosequence of restored floodplain forests: A test of the parallel dynamics hypothesis. Restor. Ecol. 2012, 20, 202–210. [Google Scholar]
- Battaglia, L.; Minchin, P.R.; Pritchett, D.W. Sixteen years of old-field succession and reestablishment of a bottomland hardwood forest in the Lower Mississippi Alluvial Valley. Wetlands 2002, 22, 1–17. [Google Scholar] [CrossRef]
- Martin, P.H.; Canham, C.D.; Marks, P.L. Why forests appear resistant to exotic plant invasions: Intentional introductions, stand dynamics, and the role of shade tolerance. Front. Ecol. Environ. 2009, 7, 142–149. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nickelson, J.B.; Holzmueller, E.J.; Groninger, J.W.; Lesmeister, D.B. Previous Land Use and Invasive Species Impacts on Long-term Afforestation Success. Forests 2015, 6, 3123-3135. https://doi.org/10.3390/f6093123
Nickelson JB, Holzmueller EJ, Groninger JW, Lesmeister DB. Previous Land Use and Invasive Species Impacts on Long-term Afforestation Success. Forests. 2015; 6(9):3123-3135. https://doi.org/10.3390/f6093123
Chicago/Turabian StyleNickelson, Joshua B., Eric J. Holzmueller, John W. Groninger, and Damon B. Lesmeister. 2015. "Previous Land Use and Invasive Species Impacts on Long-term Afforestation Success" Forests 6, no. 9: 3123-3135. https://doi.org/10.3390/f6093123
APA StyleNickelson, J. B., Holzmueller, E. J., Groninger, J. W., & Lesmeister, D. B. (2015). Previous Land Use and Invasive Species Impacts on Long-term Afforestation Success. Forests, 6(9), 3123-3135. https://doi.org/10.3390/f6093123