
Nitrogen Addition Promotes Soil Carbon Sequestration and Alters Carbon Pool Stability by Affecting Particulate Organic Carbon in a Karst Plantation

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
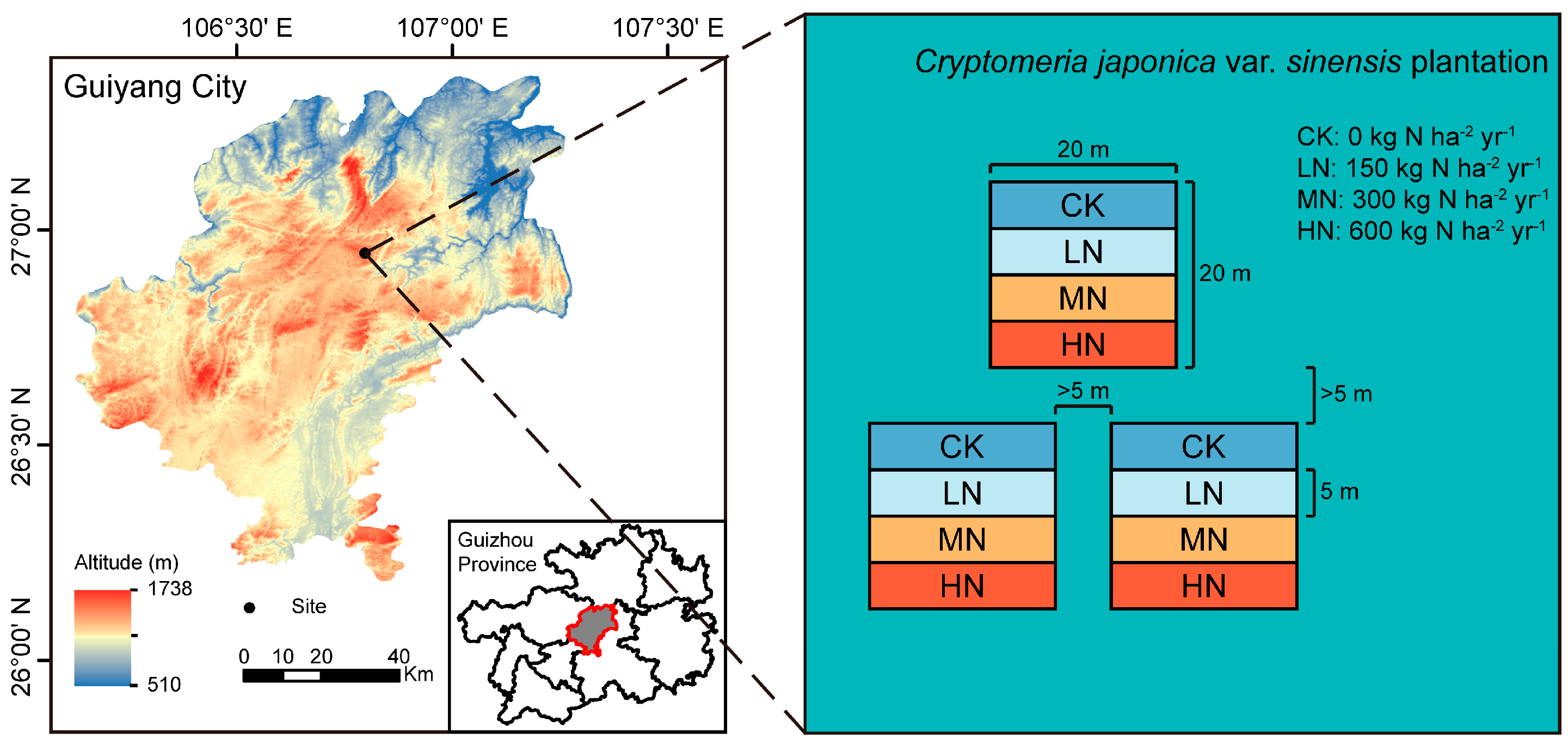
2.1. Overview of the Study Area
2.2. Experimental Design
2.3. Sample Collection and Processing
2.4. Determination of Sample Indices
2.5. Carbon Pool Lability Calculation
2.6. Sensitivity Index Calculation
2.7. Statistical Analysis
3. Results
3.1. Soil Organic Carbon
3.2. Soil Organic Carbon Fraction
3.3. Soil Carbon Pool Lability
3.4. Soil Chemical Properties
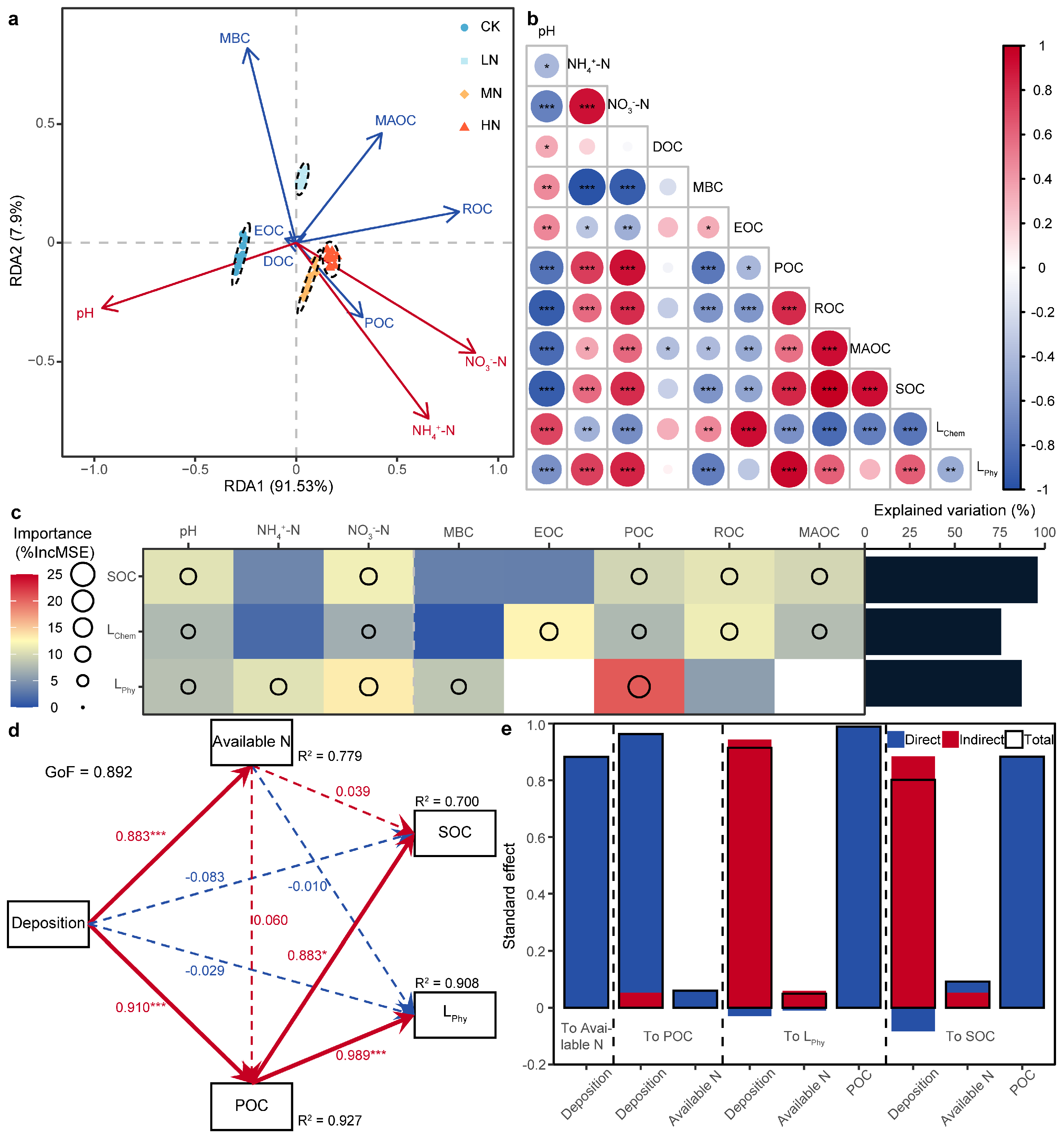
3.5. Driving Factors of Soil Organic Carbon and Carbon Pool Lability
4. Discussion
4.1. Effects of Nitrogen Addition on Soil Organic Carbon
4.2. Effects of Nitrogen Addition on Soil Organic Carbon Fractions and Carbon Pool Lability
4.3. Factors Affecting Soil Organic Carbon and Carbon Pool Lability Under Nitrogen Addition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Galloway, J.N.; Townsend, A.R.; Erisman, J.W.; Bekunda, M.; Cai, Z.C.; Freney, J.R.; Martinelli, L.A.; Seitzinger, S.P.; Sutton, M.A. Transformation of the nitrogen cycle: Recent trends, questions, and potential solutions. Science 2008, 320, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Galloway, J.N.; Cowling, E.B.; Seitzinger, S.P.; Socolow, R.H. Reactive nitrogen: Too much of a good thing? AMBIO J. Hum. Environ. 2002, 31, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Guenet, B.; Gabrielle, B.; Chenu, C.; Arrouays, D.; Balesdent, J.; Bernoux, M.; Bruni, E.; Caliman, J.-P.; Cardinael, R.; Chen, S.C.; et al. Can N2O emissions offset the benefits from soil organic carbon storage? Glob. Change Biol. 2020, 27, 237–256. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Eastman, B.A.; Adams, M.B.; Brzostek, E.R.; Burnham, M.B.; Carrara, J.E.; Kelly, C.; McNeil, B.E.; Walter, C.A.; Peterjohn, W.T. Altered plant carbon partitioning enhanced forest ecosystem carbon storage after 25 years of nitrogen additions. New Phytol. 2021, 230, 1435–1448. [Google Scholar] [CrossRef]
- Chen, Z.J.; Geng, S.C.; Zhou, X.Y.; Gui, H.R.; Zhang, L.L.; Huang, Z.Q.; Wang, M.H.; Zhang, J.H.; Han, S.J. Nitrogen addition decreases soil aggregation but enhances soil organic carbon stability in a temperate forest. Geoderma 2022, 426, 116112. [Google Scholar] [CrossRef]
- Yu, M.X.; Wang, Y.P.; Baldock, J.A.; Jiang, J.; Mo, J.M.; Zhou, G.Y.; Yan, J.H. Divergent responses of soil organic carbon accumulation to 14 years of nitrogen addition in two typical subtropical forests. Sci. Total Environ. 2020, 707, 136104. [Google Scholar] [CrossRef]
- Lu, X.F.; Ren, W.D.; Hou, E.Q.; Zhang, L.L.; Wen, D.Z.; Liu, Z.F.; Lin, Y.B.; Wang, J.; Kuang, Y.W. Negative effects of canopy N addition on soil organic carbon in wet season are primarily detected in uppermost soils of a subtropical forest. Glob. Ecol. Conserv. 2019, 17, e00543. [Google Scholar] [CrossRef]
- Ma, T.Y.; Liu, X.Y.; Xu, S.Q.; Guo, H.R.; Huang, H.; Hu, C.C.; Wu, D.; Sun, Z.C.; Chen, C.J.; Song, W. Levels and variations of soil organic carbon and total nitrogen among forests in a hotspot region of high nitrogen deposition. Sci. Total Environ. 2020, 713, 136620. [Google Scholar] [CrossRef]
- Xie, J.Y.; Chen, M.Y.; Zhang, X.; Wang, S.L.; Fang, X.M.; Xie, M.Y.; Zhang, L. Understory vegetation altered soil CO2 and N2O emissions and the correlation with plant and soil stoichiometry following N and P addition in Chinese fir plantations. Plant Soil 2023, 501, 155–170. [Google Scholar] [CrossRef]
- Tang, B.; Rocci, K.S.; Lehmann, A.; Rillig, M.C. Nitrogen increases soil organic carbon accrual and alters its functionality. Glob. Change Biol. 2023, 29, 1971–1983. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Piegholdt, C.; Andruschkewitsch, R.; Linsler, D.; Koch, H.J.; Ludwig, B. Impact of tillage intensity on carbon and nitrogen pools in surface and sub-surface soils of three long-term field experiments. Eur. J. Soil Sci. 2014, 65, 499–509. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, A.K.; Ghosh, P. Distribution of soil organic carbon and glomalin related soil protein in reclaimed coal mine-land chronosequence under tropical condition. Sci. Total Environ. 2018, 625, 1341–1350. [Google Scholar] [CrossRef]
- He, H.; Peng, M.W.; Lu, W.D.; Ru, S.B.; Hou, Z.N.; Li, J.H. Organic fertilizer substitution promotes soil organic carbon sequestration by regulating permanganate oxidizable carbon fractions transformation in oasis wheat fields. Catena 2023, 221, 106784. [Google Scholar] [CrossRef]
- Yang, X.; Meng, J.; Lan, Y.; Chen, W.F.; Yang, T.X.; Yuan, J.; Liu, S.N.; Han, J. Effects of maize stover and its biochar on soil CO2 emissions and labile organic carbon fractions in Northeast China. Agric. Ecosyst. Environ. 2017, 240, 24–31. [Google Scholar] [CrossRef]
- Blair, G.; Lefroy, R.; Lisle, L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Aust. J. Agric. Res. 1995, 46, 1459. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Samson, M.-E.; Chantigny, M.H.; Vanasse, A.; Menasseri-Aubry, S.; Royer, I.; Angers, D.A. Management practices differently affect particulate and mineral-associated organic matter and their precursors in arable soils. Soil Biol. Biochem. 2020, 148, 107867. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Cui, S.; Liang, G.P.; Zhang, Q.P. Microbial-derived carbon components are critical for enhancing soil organic carbon in no-tillage croplands: A global perspective. Soil Tillage Res. 2021, 205, 104758. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Change Biol. 2019, 26, 261–273. [Google Scholar] [CrossRef]
- Lugato, E.; Lavallee, J.M.; Haddix, M.L.; Panagos, P.; Cotrufo, M.F. Different climate sensitivity of particulate and mineral-associated soil organic matter. Nat. Geosci. 2021, 14, 295–300. [Google Scholar] [CrossRef]
- Fu, R.X.; Yu, Y.C.; He, Y.; Xu, X.N.; Sun, X.; Yang, J.; Tao, X. The impacts of nitrogen addition on the active carbon pools in forest soils along an urban–rural gradient. Catena 2024, 236, 107769. [Google Scholar] [CrossRef]
- Zhong, X.L.; Li, J.T.; Li, X.J.; Ye, Y.C.; Liu, S.S.; Hallett, P.D.; Ogden, M.R.; Naveed, M. Physical protection by soil aggregates stabilizes soil organic carbon under simulated N deposition in a subtropical forest of China. Geoderma 2017, 285, 323–332. [Google Scholar] [CrossRef]
- Chen, X.M.; Li, Y.L.; Mo, J.M.; Otieno, D.; Tenhunen, J.; Yan, J.H.; Liu, J.X.; Zhang, D.Q. Effects of nitrogen deposition on soil organic carbon fractions in the subtropical forest ecosystems of S China. J. Plant Nutr. Soil Sci. 2012, 175, 947–953. [Google Scholar] [CrossRef]
- Chen, J.Q.; Zhang, Q.F.; Dai, H.; Feng, J.G.; Zeng, Q.X.; Sun, X.Q.; Peng, Y.Z.; Chen, W.W.; Zhu, B.; Chen, Y. Nitrogen addition promotes the accumulation of soil particulate organic carbon in a subtropical forest. Forests 2024, 15, 619. [Google Scholar] [CrossRef]
- Li, Y.D.; Wang, B.; Dou, S.; Shen, H.Y.; Mei, L.Y.; Zhang, Y.; Zeng, X.M.; Zhang, Y.Y.; Pei, Y.M.; Ren, H.Y.; et al. Divergent responses of soil carbon and nitrogen pools to short-term nitrogen addition between two plantations in Northeast China. Int. J. Agric. Biol. Eng. 2019, 12, 82–90. [Google Scholar] [CrossRef]
- Zhang, J.F.; Zhou, J.G.; Sayer, E.J.; Lambers, H.; Liu, Z.F.; Lu, X.K.; Li, Y.W.; Li, Y.X.; Li, H.; Wang, F.M. Nitrogen deposition enhances soil organic carbon and microbial residual carbon in a tropical forest. Plant Soil 2022, 484, 217–235. [Google Scholar] [CrossRef]
- Liu, X.M.; Wang, C.; Gao, J.X.; Yuan, J.F.; Huang, Y.; Wang, B.; Peng, Y. Approaches to carbon sequestration enhancement in China’s plantation ecosystem for carbon peaking and carbon neutrality goals. Acta Ecol. Sin. 2023, 43, 5662–5673. [Google Scholar]
- Cheng, K.; Yang, H.T.; Tao, S.L.; Su, Y.J.; Guan, H.C.; Ren, Y.; Hu, T.Y.; Li, W.K.; Xu, G.C.; Chen, M.X.; et al. Carbon Storage through China’s Planted Forest Expansion. Nat. Commun. 2024, 15, 4106. [Google Scholar] [CrossRef]
- Du, E.Z.; Terrer, C.; Pellegrini, A.F.A.; Ahlström, A.; van Lissa, C.J.; Zhao, X.; Xia, N.; Wu, X.H.; Jackson, R.B. Global patterns of terrestrial nitrogen and phosphorus limitation. Nat. Geosci. 2020, 13, 221–226. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.H.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022. [Google Scholar]
- Yuan, Y.S.; Gu, D.P.; Huang, Z.X.; Zhang, J.L.; Xia, G.W.; Chen, L.J. Plant roots and associated mycelia enhance soil N transformation through different mechanisms in a karst plantation. J. Soils Sediments 2023, 23, 1687–1697. [Google Scholar] [CrossRef]
- Lu, R.K. Analysis Methods of Soil Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Yuan, G.Y.; Huan, W.W.; Song, H.; Lu, D.J.; Chen, X.Q.; Wang, H.Y.; Zhou, J.M. Effects of straw incorporation and potassium fertilizer on crop yields, soil organic carbon, and active carbon in the rice–wheat system. Soil Tillage Res. 2021, 209, 104958. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Tang, Z.X.; You, Y.M.; Guo, X.W.; Wu, C.J.; Liu, S.R.; Sun, O.J. Differential effects of forest-floor litter and roots on soil organic carbon formation in a temperate oak forest. Soil Biol. Biochem. 2023, 180, 109017. [Google Scholar] [CrossRef]
- Peng, X.Y.; Huang, Y.; Duan, X.W.; Yang, H.; Liu, J.X. Particulate and mineral-associated organic carbon fractions reveal the roles of soil aggregates under different land-use types in a karst faulted basin of China. Catena 2023, 220, 106721. [Google Scholar] [CrossRef]
- Sheng, H.; Zhou, P.; Zhang, Y.Z.; Kuzyakov, Y.; Zhou, Q.; Ge, T.D.; Wang, C.H. Loss of labile organic carbon from subsoil due to land-use changes in subtropical China. Soil Biol. Biochem. 2015, 88, 148–157. [Google Scholar] [CrossRef]
- Hu, Y.L.; Deng, Q.; Kätterer, T.; Olesen, J.E.; Ying, S.C.; Ochoa-Hueso, R.; Mueller, C.W.; Weintraub, M.N.; Chen, J. Depth-dependent responses of soil organic carbon under nitrogen deposition. Glob. Change Biol. 2024, 30, e17247. [Google Scholar] [CrossRef]
- Rahman, M.M.; Bárcena, T.G.; Vesterdal, L. Tree species and time since afforestation drive soil C and N mineralization on former cropland. Geoderma 2017, 305, 153–161. [Google Scholar] [CrossRef]
- Xia, G.W.; Wang, D.F.; Zhu, T.; Jiang, X.H.; Chen, G.P.; Huang, C.L. Effects of nitrogen addition and litter removal on soil respiration of Cryptomeria fortunei plantation in Karst Area of central Guizhou Province. Acta Ecol. Sin. 2023, 43, 8587–8597. [Google Scholar]
- Yuan, Y.S.; Yin, Y.C.; Adamczyk, B.; Liang, D.; Gu, D.P.; Xia, G.W.; Zhang, J.L.; Zhang, Z.L. Nitrogen addition alters the relative importance of roots and mycorrhizal hyphae in regulating soil organic carbon accumulation in a karst forest. Soil Biol. Biochem. 2024, 195, 109471. [Google Scholar] [CrossRef]
- Aber, J.; McDowell, W.; Nadelhoffer, K.; Magill, A.; Berntson, G.; Kamakea, M.; McNulty, S.; Currie, W.; Rustad, L.; Fernandez, I. Nitrogen saturation in temperate forest ecosystems: Hypotheses revisited. Bioscience 1998, 48, 921–934. [Google Scholar] [CrossRef]
- Fatemi, F.R.; Fernandez, I.J.; Simon, K.S.; Dail, D.B. Nitrogen and phosphorus regulation of soil enzyme activities in acid forest soils. Soil Biol. Biochem. 2016, 98, 171–179. [Google Scholar] [CrossRef]
- Barantal, S.; Schimann, H.; Fromin, N.; Hättenschwiler, S. C, N and P fertilization in an amazonian rainforest supports stoichiometric dissimilarity as a driver of litter diversity effects on decomposition. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141682. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, H.Y.H.; Ruan, H.H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Bloom, A.J.; Field, C.B.; Waring, R.H. Plant responses to multiple environmental factors: Physiological ecology provides tools for studying how interacting environmental resources control plant growth. Bioscience 1987, 37, 49–57. [Google Scholar] [CrossRef]
- Mo, J.M.; Zhang, W.; Zhu, W.X.; Gundersen, P.; Fang, Y.T.; Li, D.J.; Wang, H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Change Biol. 2007, 14, 403–412. [Google Scholar] [CrossRef]
- Chen, J.G.; Xiao, W.; Zheng, C.Y.; Zhu, B. Nitrogen addition has contrasting effects on particulate and mineral-associated soil organic carbon in a subtropical forest. Soil Biol. Biochem. 2020, 142, 107708. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; He, T.X.; Liu, L.; Wu, J.B. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils. Soil Biol. Biochem. 2014, 71, 13–20. [Google Scholar] [CrossRef]
- Du, Y.H.; Guo, P.; Liu, J.Q.; Wang, C.Y.; Yang, N.; Jiao, Z.X. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Glob. Change Biol. 2014, 20, 3222–3228. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.Y.; Wang, Y.X.; Li, J.L. Effects of nitrogen addition on plant-soil carbon dynamics in terrestrial ecosystems of China. Acta Ecol. Sin. 2022, 42, 4823–4833. [Google Scholar]
- Deng, W.B.; Wang, X.; Hu, H.B.; Zhu, M.D.; Chen, J.Y.; Zhang, S.; Cheng, C.; Zhu, Z.Y.; Wu, C.M.; Zhu, L. Variation characteristics of soil organic carbon storage and fractions with stand age in north subtropical Quercus acutissima Carruth. forest in China. Forests 2022, 13, 1649. [Google Scholar] [CrossRef]
- Liu, J.; Wu, N.N.; Wang, H.; Sun, J.F.; Peng, B.; Jiang, P.; Bai, E. Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter. Ecology 2016, 97, 1796–1806. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.-A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef]
- Hu, X.B.; Ji, F.Y.; Li, S.; Zhou, G.M.; Yu, D.N.; Tan, X.M.; Yang, D.C.; Yu, B. Study of vibrational spectra of humic substance in soils from the Three Gorges Reservoir Area. Spectrosc. Spectr. Anal. 2010, 30, 1376–1380. [Google Scholar]
- Meng, D.Y.; Cheng, H.X.; Shao, Y.; Luo, M.; Xu, D.D.; Liu, Z.M.; Ma, L.L. Progress on the effect of nitrogen on transformation of soil organic carbon. Processes 2022, 10, 2425. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Hou, Y.H.; Zhou, S.R.; Zhu, B. Particulate organic carbon is more vulnerable to nitrogen addition than mineral-associated organic carbon in soil of an alpine meadow. Plant Soil 2019, 458, 93–103. [Google Scholar] [CrossRef]
- Wang, M.H.; Li, F.C.; Dong, L.L.; Wang, X.; Han, L.B.; Olesen, J.E. Effects of exogenous organic/inorganic nitrogen addition on carbon pool distribution and transformation in grassland soil. Sci. Total Environ. 2023, 858, 159919. [Google Scholar] [CrossRef]
- Hong, S.B.; Gan, P.; Chen, A.P. Environmental controls on soil pH in planted forest and its response to nitrogen deposition. Environ. Res. 2019, 172, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, F.E. Effect of fertilizer nitrogen on the release of soil nitrogen. Soil Sci. Soc. Am. J. 1965, 29, 692–696. [Google Scholar] [CrossRef]
- Lu, X.F.; Gilliam, F.S.; Guo, J.Y.; Hou, E.Q.; Kuang, Y.W. Decrease in soil pH has greater effects than increase in above-ground carbon inputs on soil organic carbon in terrestrial ecosystems of China under nitrogen enrichment. J. Appl. Ecol. 2021, 59, 768–778. [Google Scholar] [CrossRef]
- Riggs, C.E.; Hobbie, S.E.; Bach, E.M.; Hofmockel, K.S.; Kazanski, C.E. Nitrogen addition changes grassland soil organic matter decomposition. Biogeochemistry 2015, 125, 203–219. [Google Scholar] [CrossRef]
- Man, M.L.; Pierson, D.; Chiu, R.; Tabatabaei Anaraki, M.; vandenEnden, L.; Ye, R.X.; Lajtha, K.; Simpson, M.J. Twenty years of litter manipulation reveals that above-ground litter quantity and quality controls soil organic matter molecular composition. Biogeochemistry 2022, 159, 393–411. [Google Scholar] [CrossRef]
- Almeida, L.F.J.; Souza, I.F.; Hurtarte, L.C.C.; Teixeira, P.P.C.; Inagaki, T.M.; Silva, I.R.; Mueller, C.W. Forest litter constraints on the pathways controlling soil organic matter formation. Soil Biol. Biochem. 2021, 163, 108447. [Google Scholar] [CrossRef]



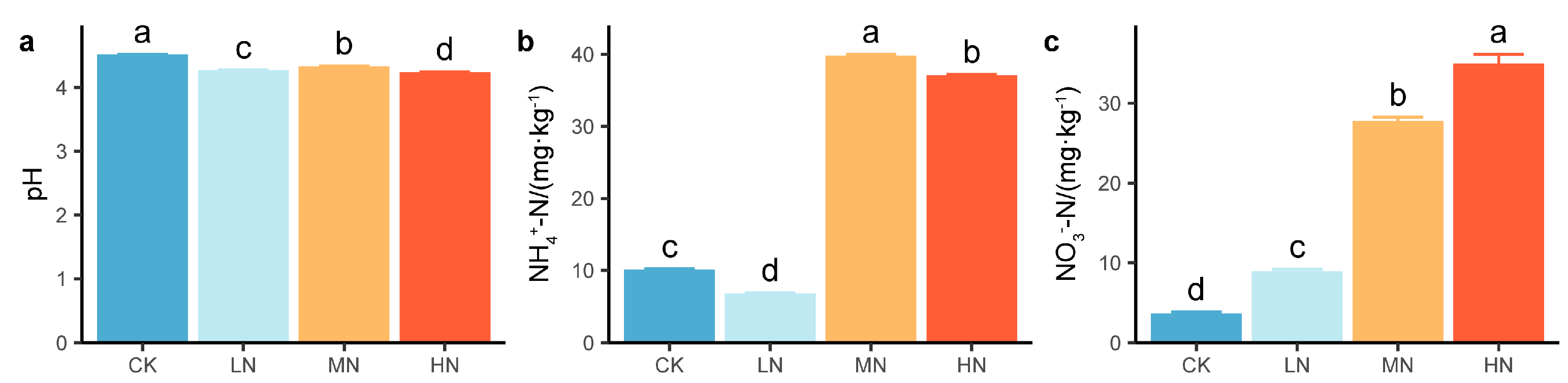

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | SOC | DOC | MBC | EOC | POC | ROC | MAOC |
|---|---|---|---|---|---|---|---|
| LN | 31.91 | −39.34 | 4.25 | −23.52 | 46.22 | 39.23 | 29.12 |
| MN | 33.06 | −22.19 | −80.17 | −29.23 | 72.49 | 41.29 | 25.38 |
| HN | 41.20 | −19.06 | −79.09 | −21.37 | 124.00 | 49.46 | 25.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, T.; Xia, G.; Yuan, Y.; Lu, Q.; Jiang, X.; Huang, C.; Zhou, W. Nitrogen Addition Promotes Soil Carbon Sequestration and Alters Carbon Pool Stability by Affecting Particulate Organic Carbon in a Karst Plantation. Forests 2025, 16, 730. https://doi.org/10.3390/f16050730
Zhu T, Xia G, Yuan Y, Lu Q, Jiang X, Huang C, Zhou W. Nitrogen Addition Promotes Soil Carbon Sequestration and Alters Carbon Pool Stability by Affecting Particulate Organic Carbon in a Karst Plantation. Forests. 2025; 16(5):730. https://doi.org/10.3390/f16050730
Chicago/Turabian StyleZhu, Tao, Guowei Xia, Yuanshuang Yuan, Qi Lu, Xiaohan Jiang, Chengling Huang, and Wei Zhou. 2025. "Nitrogen Addition Promotes Soil Carbon Sequestration and Alters Carbon Pool Stability by Affecting Particulate Organic Carbon in a Karst Plantation" Forests 16, no. 5: 730. https://doi.org/10.3390/f16050730
APA StyleZhu, T., Xia, G., Yuan, Y., Lu, Q., Jiang, X., Huang, C., & Zhou, W. (2025). Nitrogen Addition Promotes Soil Carbon Sequestration and Alters Carbon Pool Stability by Affecting Particulate Organic Carbon in a Karst Plantation. Forests, 16(5), 730. https://doi.org/10.3390/f16050730