
Quantification of the Influencing Factors of Stand Productivity of Subtropical Natural Broadleaved Forests in Eastern China Using an Explainable Machine Learning Framework

Abstract
1. Introduction
2. Materials and Methods
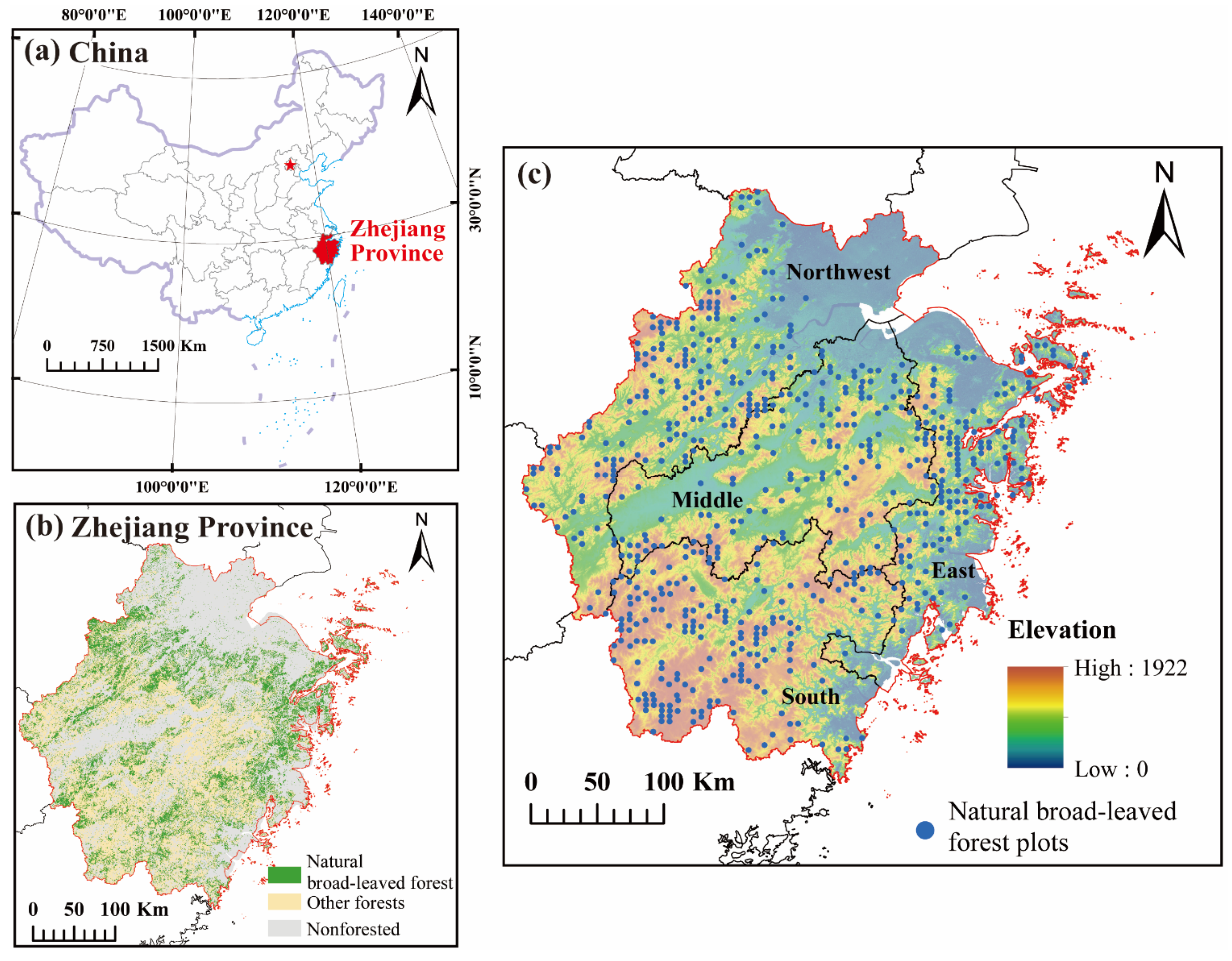
2.1. Study Area
2.2. Data Collection and Processing
2.2.1. Stand Productivity Calculation
2.2.2. Influencing Factor Data
2.2.3. Influencing Factor Screening Based on Boruta’s Algorithm
2.3. Machine Learning Models
2.4. SHAP (Shapley Additive Explanation) Algorithm
3. Results
3.1. Importance Analysis Screening of Influencing Factors
3.2. Forecasting Performances of Machine Learning Models
3.3. Model Interpretability Analysis
3.3.1. Global and Local Model Interpretability
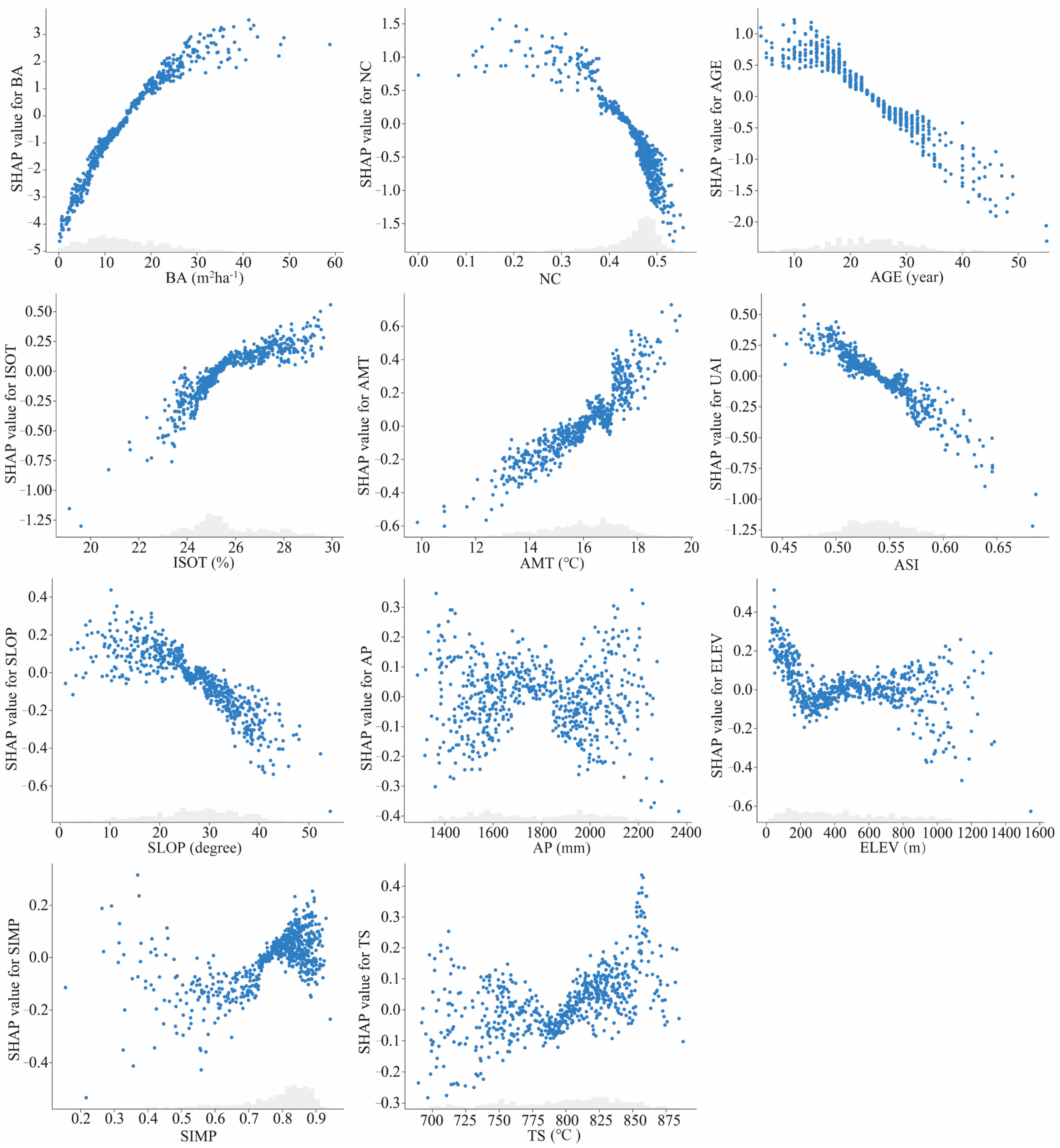
3.3.2. Factor Dependence Analysis
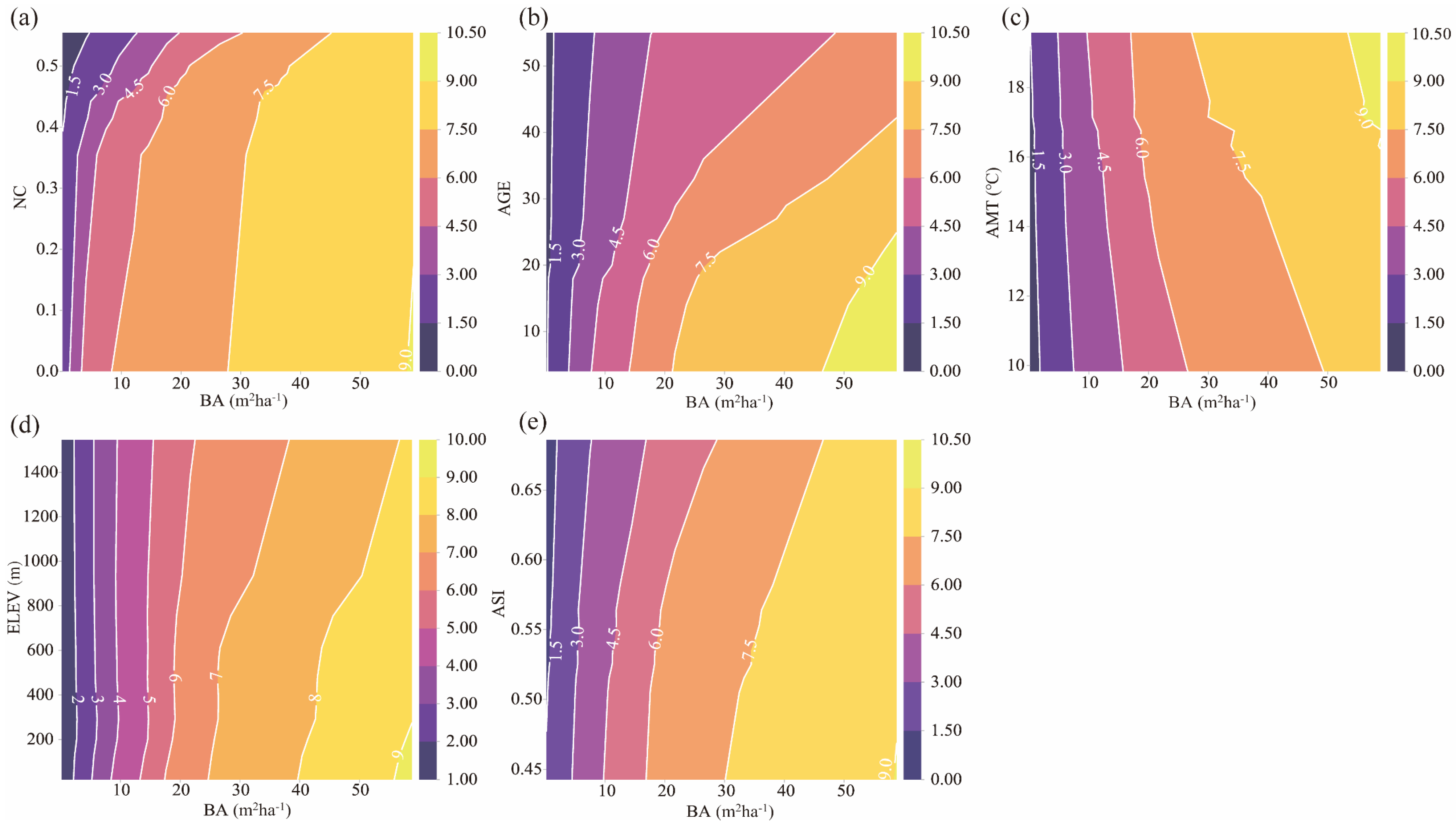
3.3.3. Two-Factor Interaction Analysis
4. Discussion
4.1. Tree Volume Model and Stand Productivity Parameter Selection
4.2. Machine Learning Model and Influencing Factor Selection
4.3. Basal Area Had Strongest Influence on Stand Productivity
4.4. Stand Spatial Structure: The Main Influencer of Stand Productivity
4.5. Increasing Stand Productivity Through Effective Forest Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Souza, C.R.; Mariano, R.F.; Maia, V.A.; Pompeu, P.V.; Santos, R.M.D.; Fontes, M.A.L. Carbon stock and uptake in the high-elevation tropical montane forests of the threatened Atlantic Forest hotspot: Ecosystem function and effects of elevation variation. Sci. Total Environ. 2023, 882, 163503. [Google Scholar] [CrossRef]
- Crow, T.R.; Dey, D.C.; Riemenschneider, D. Forest Productivity: Producing Goods and Services for People; U.S. Department of Agriculture, Forest Service, North Central Research Station: Madison, WI, USA, 2006. [Google Scholar]
- Mamo, N.; Sterba, H. Site index functions for Cupressus lusitanica at Munesa Shashemene, Ethiopia. For. Ecol. Manag. 2006, 237, 429–435. [Google Scholar] [CrossRef]
- Herraiz, A.D.; Salazar-Zarzosa, P.C.; Mesas, F.J.; Arenas-Castro, S.; Ruiz-Benito, P.; Villar, R. Modelling aboveground biomass and productivity and the impact of climate change in Mediterranean forests of South Spain. Agric. For. Meteorol. 2023, 337, 109498. [Google Scholar] [CrossRef]
- Sotomayor, L.N.; Cracknell, M.J.; Musk, R. Supervised machine learning for predicting and interpreting dynamic drivers of plantation forest productivity in northern Tasmania, Australia. Comput. Electron. Agric. 2023, 209, 107804. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Wang, X.; Xiao, W.; Chen, L.; Li, S.; Sun, H.; Deng, X.; Forrester, D.I.; Zeng, L.; et al. Effects of stand age, richness and density on productivity in subtropical forests in China. J. Ecol. 2019, 107, 2266–2277. [Google Scholar] [CrossRef]
- Liang, J.; Crowther, T.W.; Picard, N.; Wiser, S.; Zhou, M.; Alberti, G.; Schulze, E.-D.; McGuire, A.D.; Bozzato, F. Positive biodiversity-productivity relationship predominant in global forests. Science 2016, 354, 196. [Google Scholar] [CrossRef]
- Kweon, D.; Comeau, P.G. Factors influencing productivity of pine-dominated stands in South Korea. J. Environ. Manag. 2023, 330, 117250. [Google Scholar] [CrossRef] [PubMed]
- Salazar Zarzosa, P.; Diaz Herraiz, A.; Olmo, M.; Ruiz-Benito, P.; Barron, V.; Bastias, C.C.; de la Riva, E.G.; Villar, R. Linking functional traits with tree growth and forest productivity in Quercus ilex forests along a climatic gradient. Sci. Total Environ. 2021, 786, 147468. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Chhin, S.; Zhang, J.; Duan, A. Disentangling the effects of stand and climatic variables on forest productivity of Chinese fir plantations in subtropical China using a random forest algorithm. Agric. For. Meteorol. 2021, 304, 108412. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, J.; Wang, J.; Gu, Y.; Han, S. A linear positive relationship between tree species diversity and forest productivity across forest-dominated natural reserves on a large spatial scale. For. Ecol. Manag. 2023, 548, 121409. [Google Scholar] [CrossRef]
- Kara, F.; Keleş, S.Ö. Tree species richness influence productivity and anatomical characteristics in mixed fir-pine-beech forests. Plant Ecol. 2023, 224, 479–489. [Google Scholar] [CrossRef]
- Chetelat, R.T.; Pertuzé, R.A.; Faúndez, L.; Graham, E.B.; Jones, C.M. Distribution, ecology and reproductive biology of wild tomatoes and related nightshades from the Atacama Desert region of northern Chile. Euphytica 2008, 167, 77–93. [Google Scholar] [CrossRef]
- Rosas, T.; Mencuccini, M.; Barba, J.; Cochard, H.; Saura-Mas, S.; Martinez-Vilalta, J. Adjustments and coordination of hydraulic, leaf and stem traits along a water availability gradient. New Phytol. 2019, 223, 632–646. [Google Scholar] [CrossRef]
- Oboite, F.O.; Comeau, P.G. Competition and climate influence growth of black spruce in western boreal forests. For. Ecol. Manag. 2019, 443, 84–94. [Google Scholar] [CrossRef]
- Marcatti, G.E.; Resende, R.T.; Resende, M.D.V.; Ribeiro, C.A.A.S.; dos Santos, A.R.; da Cruz, J.P.; Leite, H.G. GIS-based approach applied to optimizing recommendations of Eucalyptus genotypes. For. Ecol. Manag. 2017, 392, 144–153. [Google Scholar] [CrossRef]
- Ameray, A.; Cavard, X.; Bergeron, Y. Climate change may increase Quebec boreal forest productivity in high latitudes by shifting its current composition. Front. For. Glob. Chang. 2023, 6, 1020305. [Google Scholar] [CrossRef]
- Ammer, C. Diversity and forest productivity in a changing climate. New Phytol. 2019, 221, 50–66. [Google Scholar] [CrossRef]
- Baach, E.; Himes, A.; Polinko, A.; Granger, J.J.; Zhou, Q. Diversity-productivity relationships in forests of the southeastern United States: Leveraging national inventory data and tree functional traits. For. Ecol. Manag. 2022, 521, 120426. [Google Scholar] [CrossRef]
- Kohyama, T.I.; Sheil, D.; Sun, I.F.; Niiyama, K.; Suzuki, E.; Hiura, T.; Nishimura, N.; Hoshizaki, K.; Wu, S.H.; Chao, W.C.; et al. Contribution of tree community structure to forest productivity across a thermal gradient in eastern Asia. Nat. Commun. 2023, 14, 1113. [Google Scholar] [CrossRef]
- Ma, S.; Wang, X.; Miao, W.; Wang, X.; Sun, H.; Guo, Z. Relative influence of environmental, stand factors and functional traits on allocation of forest productivity during the restoration of subtropical forests in central China. For. Ecol. Manag. 2021, 482, 118814. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J. A Review of Processes Behind Diversity—Productivity Relationships in Forests. Curr. For. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef]
- Forrester, D.I. Linking forest growth with stand structure: Tree size inequality, tree growth or resource partitioning and the asymmetry of competition. For. Ecol. Manag. 2019, 447, 139–157. [Google Scholar] [CrossRef]
- Forrester, D.I.; Kohnle, U.; Albrecht, A.T.; Bauhus, J. Complementarity in mixed-species stands of Abies alba and Picea abies varies with climate, site quality and stand density. For. Ecol. Manag. 2013, 304, 233–242. [Google Scholar] [CrossRef]
- Garber, S.M.; Maguire, D.A. Stand Productivity and Development in Two Mixed-Species Spacing Trials in the Central Oregon Cascades. For. Sci. 2004, 50, 92–105. [Google Scholar] [CrossRef]
- John, R.; Chen, J.; Kim, Y.; Ou-yang, Z.-t.; Xiao, J.; Park, H.; Shao, C.; Zhang, Y.; Amarjargal, A.; Batkhshig, O.; et al. Differentiating anthropogenic modification and precipitation-driven change on vegetation productivity on the Mongolian Plateau. Landsc. Ecol. 2015, 31, 547–566. [Google Scholar] [CrossRef]
- Zhang, M.; Fan, X.; Yue, Q.; Han, Z.; Huang, Y. Effects of Biotic and Abiotic Factors on Productivity of Coniferous and Broad-Leaved Mixed Forest in Jiaohe, Jilin Province. Sci. Silvae Sin. 2023, 59, 71–77. [Google Scholar]
- Zou, J.; Luo, Y.; Seidl, R.; Thom, D.; Liu, J.; Geres, L.; Richter, T.; Ye, L.; Zheng, W.; Ma, L.; et al. No generality in biodiversity-productivity relationships along elevation in temperate and subtropical forest landscapes. For. Ecosyst. 2024, 11, 100187. [Google Scholar] [CrossRef]
- Rocha, S.J.S.S.d.; Torres, C.M.M.E.; Villanova, P.H.; Tavares Júnior, I.d.S.; Rufino, M.P.M.X.; Romero, F.M.B.; Jacovine, L.A.G.; de Morais Junior, V.T.M.; França, L.C.d.J.; Schettini, B.L.S.; et al. Machine learning methods: Modeling net growth in the Atlantic Forest of Brazil. Ecol. Inform. 2024, 81, 102564. [Google Scholar] [CrossRef]
- Dong, L.; Lin, X.; Bettinger, P.; Liu, Z. The contributions of stand characteristics on carbon sequestration potential are triple that of climate variables for Larix spp. plantations in northeast China. Sci. Total Environ. 2024, 911, 168726. [Google Scholar] [CrossRef]
- Becknell, J.M.; Powers, J.S. Stand age and soils as drivers of plant functional traits and aboveground biomass in secondary tropical dry forest. Can. J. For. Res. 2014, 44, 604–613. [Google Scholar] [CrossRef]
- Qiao, X.; Hautier, Y.; Geng, Y.; Wang, S.; Wang, J.; Zhang, N.; Zhang, Z.; Zhang, C.; Zhao, X.; von Gadow, K. Biodiversity contributes to stabilizing ecosystem productivity across spatial scales as much as environmental heterogeneity in a large temperate forest region. For. Ecol. Manag. 2023, 529, 120695. [Google Scholar] [CrossRef]
- Hu, J.; Xu, J.; Li, M.; Jiang, Z.; Mao, J.; Feng, L.; Miao, K.; Li, H.; Chen, J.; Bai, Z.; et al. Identification and validation of an explainable prediction model of acute kidney injury with prognostic implications in critically ill children: A prospective multicenter cohort study. EClinicalMedicine 2024, 68, 102409. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, R.; Song, G.; Zhang, K.; Zhao, Z.; Wang, J. Interpretable ensemble prediction for anaerobic digestion performance of hydrothermal carbonization wastewater. Sci. Total Environ. 2024, 908, 168279. [Google Scholar] [CrossRef]
- Museru, M.L.; Nazari, R.; Giglou, A.N.; Opare, K.; Karimi, M. Advancing flood damage modeling for coastal Alabama residential properties: A multivariable machine learning approach. Sci. Total Environ. 2024, 907, 167872. [Google Scholar] [CrossRef]
- Liu, A. Mathematical modeling of volume of living tree in Zhejiang Province. J. Zhejiang For. Sci. Technol. 1986, 4, 25–30. [Google Scholar]
- Hui, G.; Gadow, K.v.; Albert, M. A new parameter for stand spatial structure neighbourhood comparison. For. Res. 1999, 12, 1–6. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Erickson, N.; Mueller, J.; Shirkov, A.; Zhang, H.; Larroy, P.; Li, M.; Smola, A. AutoGluon-Tabular: Robust and Accurate AutoML for Structured Data. arXiv 2020, arXiv:2003.06505. [Google Scholar]
- Qi, W.; Xu, C.; Xu, X. AutoGluon: A revolutionary framework for landslide hazard analysis. Nat. Hazards Res. 2021, 1, 103–108. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Y.; Feng, Y.; Yu, Y.; Li, Y.; Li, J.; Ren, Z.; Chen, S.; Feng, L.; Pan, J.; et al. Novel Intelligent System Based on Automated Machine Learning for Multiobjective Prediction and Early Warning Guidance of Biogas Performance in Industrial-Scale Garage Dry Fermentation. ACS EST Eng. 2023, 4, 139–152. [Google Scholar] [CrossRef]
- Lin, X.; Hou, J.; Wu, X.; Lin, D. Elucidating the impacts of microplastics on soil greenhouse gas emissions through automatic machine learning frameworks. Sci. Total Environ. 2024, 916, 170308. [Google Scholar] [CrossRef]
- Lai, J.; Zou, Y.; Zhang, S.; Zhang, X.; Mao, L.; Zhang, W.H. glmm.hp: An R package for computing individual effect of predictors in generalized linear mixed models. J. Plant Ecol. 2022, 15, 1302–1307. [Google Scholar] [CrossRef]
- Li, Z. Extracting spatial effects from machine learning model using local interpretation method: An example of SHAP and XGBoost. Comput. Environ. Urban Syst. 2022, 96, 101845. [Google Scholar] [CrossRef]
- Shapley, L.S. A Value for n-Person Games; Princeton University Press: Princeton, NJ, USA, 1953. [Google Scholar]
- Xu, Y.; Zhang, D.; Lin, J.; Peng, Q.; Lei, X.; Jin, T.; Wang, J.; Yuan, R. Prediction of phytoplankton biomass and identification of key influencing factors using interpretable machine learning models. Ecol. Indic. 2024, 158, 111320. [Google Scholar] [CrossRef]
- Lundberg, S.M.; Lee, S.-I. A unified approach to interpreting model predictions. Neural Inf. Process. Syst. 2017, 30, 4768–4777. [Google Scholar]
- Ji, B.; Tao, J.; Zhang, G.; Du, Q.; Yao, H.; Xu, J. Zhejiang Province’s forest vegetation biomass assessment for guaranteed accuracy. J. Zhejiang AF Univ. 2012, 29, 328–334. [Google Scholar]
- Ji, B.; Zhang, G.; Zhao, G.; Lai, J. A Technical Method of Dynamic Forest Resources Monitoring at the County Level Based on the Permanent Plots. For. Resour. Manag. 2009, 5, 50–53. [Google Scholar]
- Sun, K.; Wu, D.; Sun, H. Effects of Mixed Betula luminifera and Phyllostachys edulis with Different Proportions on the Growth of Cunninghamia lanceolata after Thinning. For. Grassl. Resour. Res. 2024, 2, 124–132. [Google Scholar]
- Brickell, J.E. More on Diameter Tape and Calipers. J. For. 1970, 68, 169–170. [Google Scholar]
- Mao, Z. Compilation of the two-way volume model for Zhejiang Province. J. Zhejiang AF Univ. 1988, 5, 75–80. [Google Scholar]
- Inoue, A.; Sakamoto, S.; Suga, H.; Kitazato, H.; Sakuta, K. Construction of one-way volume table for the three major useful bamboos in Japan. J. For. Res. 2013, 18, 323–334. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, L.; Lu, C.; Fu, Y.; Gu, Z.; Chen, Y.; Zhang, G.; Zhou, X. Drought- induced decrease in tree productivity mainly mediated by the maximum growth rate and growing-season length in a subtropical forest. For. Ecol. Manag. 2024, 563, 121985. [Google Scholar] [CrossRef]
- Hisano, M.; Ghazoul, J.; Chen, X.; Chen, H.Y.H. Functional diversity enhances dryland forest productivity under long-term climate change. Sci. Adv. 2024, 10, eadn4152. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2010, 20, 170–180. [Google Scholar] [CrossRef]
- Zhang, X.; Su, J.; Ji, Y.; Zhao, J.; Gao, J. Nitrogen deposition affects the productivity of planted and natural forests by modulating forest climate and community functional traits. For. Ecol. Manag. 2024, 563, 121970. [Google Scholar] [CrossRef]
- Jain, S.; Jain, S.; Wolf, I.T.; Lee, J.; Tong, Y.W. A comprehensive review on operating parameters and different pretreatment methodologies for anaerobic digestion of municipal solid waste. Renew. Sustain. Energy Rev. 2015, 52, 142–154. [Google Scholar] [CrossRef]
- Madrigal-González, J.; Zavala, M.A. Competition and tree age modulated last century pine growth responses to high frequency of dry years in a water limited forest ecosystem. Agric. For. Meteorol. 2014, 192, 18–26. [Google Scholar] [CrossRef]
- Calama, R.; Conde, M.; de-Dios-García, J.; Madrigal, G.; Vázquez-Piqué, J.; Gordo, F.J.; Pardos, M. Linking climate, annual growth and competition in a Mediterranean forest: Pinus pinea in the Spanish Northern Plateau. Agric. For. Meteorol. 2019, 264, 309–321. [Google Scholar] [CrossRef]
- Beyer, R.M.; Krapp, M.; Manica, A. High-resolution terrestrial climate, bioclimate and vegetation for the last 120,000 years. Sci. Data 2020, 7, 236. [Google Scholar] [CrossRef]
- Wang, Y.; Pederson, N.; Ellison, A.M.; Buckley, H.L.; Case, B.S.; Liang, E.; Julio Camarero, J. Increased stem density and competition may diminish the positive effects of warming at alpine treeline. Ecology 2016, 97, 1668–1679. [Google Scholar] [CrossRef]
- Pretzsch, H. Analysis and modeling of spatial stand structures. Methodological considerations based on mixed beech-larch stands in Lower Saxony. For. Ecol. Manag. 1997, 97, 237–253. [Google Scholar] [CrossRef]
- Huang, X.; Chen, Y.; Tan, H.; Zhang, Y.; Yu, S.; Chen, X.; Yu, K.; Liu, J. Extraction of the spatial structure of Chinese fir plantations stands based on unmanned aerial vehicle and its effect on AGB. For. Ecol. Manag. 2024, 558, 121800. [Google Scholar] [CrossRef]
- Yang, B.; Ma, R.; Zhai, J.; Du, J.; Bai, J.; Zhang, W. Stand spatial structure is more important than species diversity in enhancing the carbon sink of fragile natural secondary forest. Ecol. Indic. 2024, 558, 121800. [Google Scholar] [CrossRef]
- Wiedemann, E. Die Rotbuche 1931. Fortführung des Berichtes von Geheimrat Schwappach 1911 über die Preussischen BuchenVersuchsfläche; Schaper: Niedersachsen, Germany, 1932. [Google Scholar]
- Langsaeter, A. Om tynning i enaldret gran-og furuskog (About thinning in even-aged stands of spruce). Nor. Skogforsokresen 1941, 8, 131–216. [Google Scholar]
- Assmann, E. Grundflächen- und Volumzuwachs der Rotbuche bei verschiedenen Durchforstungsgraden. Forstwiss. Cent. 1950, 69, 256–286. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dauber, E. Long-term stand dynamics of managed spruce–fir–beech mountain forests in Central Europe: Structure, productivity and regeneration success. Forestry 2015, 88, 407–428. [Google Scholar] [CrossRef]
- Allen, M.G.; Burkhart, H.E. Growth-Density Relationships in Loblolly Pine Plantations. For. Sci. 2019, 65, 250–264. [Google Scholar] [CrossRef]
- Gizachew, B.; Brunner, A. Density–growth relationships in thinned and unthinned Norway spruce and Scots pine stands in Norway. Scand. J. For. Res. 2011, 26, 543–554. [Google Scholar] [CrossRef]
- Nilsson, U.; Agestam, E.; Ekö, P.-M.; Elfving, B.; Fahlvik, N.; Johansson, U.; Karlsson, K.; Lundmark, T.; Wallentin, C. Thinning of Scots pine and Norway spruce monocultures in Sweden: Effects of different thinning programmes on stand level gross- and net stem volume production. Stud. For. Suec. 2010, 219, 1–46. [Google Scholar]
- del Río, M.; Sterba, H. Comparing volume growth in pure and mixed stands of Pinus sylvestris and Quercus pyrenaica. Ann. For. Sci. 2009, 66, 502. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Martinez-Ramos, M.; Bongers, F. Biomass is the main driver of changes in ecosystem process rates during tropical forest succession. Ecology 2015, 96, 1242–1252. [Google Scholar] [CrossRef]
- Ali, A.; Lin, S.L.; He, J.K.; Kong, F.M.; Yu, J.H.; Jiang, H.S. Big-sized trees overrule remaining trees’ attributes and species richness as determinants of aboveground biomass in tropical forests. Glob. Chang. Biol. 2019, 25, 2810–2824. [Google Scholar] [CrossRef] [PubMed]
- Manuel Villa, P.; Ali, A.; Venâncio Martins, S.; Nolasco de Oliveira Neto, S.; Cristina Rodrigues, A.; Teshome, M.; Alvim Carvalho, F.; Heringer, G.; Gastauer, M. Stand structural attributes and functional trait composition overrule the effects of functional divergence on aboveground biomass during Amazon forest succession. For. Ecol. Manag. 2020, 477, 118481. [Google Scholar] [CrossRef]
- Miao, W.; Ma, S.; Guo, Z.; Sun, H.; Wang, X.; Lyu, Y.; Wang, X.; Ma, K. Effects of biodiversity, stand factors and functional identity on biomass and productivity during the restoration of subtropical forests in Central China. J. Plant Ecol. 2022, 15, 385–398. [Google Scholar] [CrossRef]
- Xu, K.; Wang, X.; Liang, P.; Wu, Y.; An, H.; Sun, H.; Wu, P.; Wu, X.; Li, Q.; Guo, X.; et al. A new tree-ring sampling method to estimate forest productivity and its temporal variation accurately in natural forests. For. Ecol. Manag. 2019, 433, 217–227. [Google Scholar] [CrossRef]
- Yuan, Z.; Ali, A.; Wang, S.; Gazol, A.; Freckleton, R.; Wang, X.; Lin, F.; Ye, J.; Zhou, L.; Hao, Z.; et al. Abiotic and biotic determinants of coarse woody productivity in temperate mixed forests. Sci. Total Environ. 2018, 630, 422–431. [Google Scholar] [CrossRef]
- van der Sande, M.T.; Peña-Claros, M.; Ascarrunz, N.; Arets, E.J.M.M.; Licona, J.C.; Toledo, M.; Poorter, L.; Hector, A. Abiotic and biotic drivers of biomass change in a Neotropical Forest. J. Ecol. 2017, 105, 1223–1234. [Google Scholar] [CrossRef]
- Yang, Q.; Ding, J.; Siemann, E.; Godoy, O. Biogeographic variation of distance-dependent effects in an invasive tree species. Funct. Ecol. 2019, 33, 1135–1143. [Google Scholar] [CrossRef]
- Wu, C.; Yuan, W.; Sheng, W.; Huan, Y.; Chen, Q.; Shen, A.; Zhu, J.; Jiang, B. Spatial distribution patterns and associations of tree species in typical natural secondary forest communities in Zhejiang Province. Acta Ecol. Sin. 2018, 38, 537–549. [Google Scholar]
- Zhong, L.; Liu, J.; Ding, W.; Tian, Y.; Chen, J.; Li, M.; Yu, M. Comparative research of the structures of plant functional groups in different successional stages of lowland secondary forests in Zhejiang Province. J. Zhejiang Univ. (Sci. Ed.) 2014, 41, 593–599. [Google Scholar]
- Yuan, Z.; Ali, A.; Sanaei, A.; Ruiz-Benito, P.; Jucker, T.; Fang, L.; Bai, E.; Ye, J.; Lin, F.; Fang, S.; et al. Few large trees, rather than plant diversity and composition, drive the above-ground biomass stock and dynamics of temperate forests in northeast China. For. Ecol. Manag. 2021, 481, 118698. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Ruger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, X.; Zhang, X.; Booth, T.H.; Lu, F. Root:shoot ratios across China’s forests: Forest type and climatic effects. For. Ecol. Manag. 2012, 269, 19–25. [Google Scholar] [CrossRef]
- Mokany, K.; Raison, R.J.; Prokushkin, A.S. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. 2005, 12, 84–96. [Google Scholar] [CrossRef]
- Sun, H.; Diao, S.; Liu, R.; Forrester, D.; Soares, A.; Saito, D.; Dong, R.; Jiang, J. Relationship between size inequality and stand productivity is modified by self-thinning, age, site and planting density in Sassafras tzumu plantations in central China. For. Ecol. Manag. 2018, 422, 199–206. [Google Scholar] [CrossRef]
- Wang, J.; Xu, S.; Ji, B.; Du, Q. Effects of topography and stand spatial structure on the diameter at breast height growth of major pioneer tree species of natural broad-leaved mixed forests in Zhejiang Province, China. Chin. J. Appl. Ecol. 2024, 35, 298–306. [Google Scholar]
- Soares, A.A.V.; Leite, H.G.; Souza, A.L.; Silva, S.R.; Lourenço, H.M.; Forrester, D.I. Increasing stand structural heterogeneity reduces productivity in Brazilian Eucalyptus monoclonal stands. For. Ecol. Manag. 2016, 373, 26–32. [Google Scholar] [CrossRef]
- Luu, T.C.; Binkley, D.; Stape, J.L. Neighborhood uniformity increases growth of individual Eucalyptus trees. For. Ecol. Manag. 2013, 289, 90–97. [Google Scholar] [CrossRef]
- Zhu, Y.; Mi, X.; Ren, H.; Ma, K. Density dependence is prevalent in a heterogeneous subtropical forest. Oikos 2010, 119, 109–119. [Google Scholar] [CrossRef]
- Zeller, L.; Pretzsch, H. Effect of forest structure on stand productivity in Central European forests depends on developmental stage and tree species diversity. For. Ecol. Manag. 2019, 434, 193–204. [Google Scholar] [CrossRef]
- Vanhellemont, M.; Bijlsma, R.-J.; De Keersmaeker, L.; Vandekerkhove, K.; Verheyen, K. Species and structural diversity affect growth of oak, but not pine, in uneven-aged mature forests. Basic Appl. Ecol. 2018, 27, 41–50. [Google Scholar] [CrossRef]
- Liu, L.; Zeng, F.; Song, T.; Wang, K.; Du, H. Stand Structure and Abiotic Factors Modulate Karst Forest Biomass in Southwest China. Forests 2020, 11, 443. [Google Scholar] [CrossRef]
- Coomes, D.A.; Holdaway, R.J.; Kobe, R.K.; Lines, E.R.; Allen, R.B. A general integrative framework for modelling woody biomass production and carbon sequestration rates in forests. J. Ecol. 2011, 100, 42–64. [Google Scholar] [CrossRef]
- Curtis, R.O. Technical Commentary: A New Look at an Old Question—Douglas-Fir Culmination Age. West. J. Appl. For. 1992, 7, 97–99. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Chai, Z. An R Package for Forest Spatial Structure Analysis. 2016. Available online: https://github.com/Zongzheng/forestSAS (accessed on 18 February 2024).
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; Association for Computing Machinery: New York, NY, USA, 2016; pp. 785–794. [Google Scholar]
- Cortes, C.; Vapnik, V. Support-vector networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Dorogush, A.V.; Ershov, V.; Gulin, A. CatBoost: Gradient boosting with categorical features support. arXiv 2018, arXiv:1810.11363. [Google Scholar]
- Friedman, J.H. Stochastic gradient boosting. Comput. Stat. Data Anal. 2002, 38, 367–378. [Google Scholar] [CrossRef]
- Hui, G.; Gadow, K.v.; Albert, M. A new structure parameter for describing distribution of forest tree position. Sci. Silvae Sin. 1999, 35, 37–42. [Google Scholar]
- Ke, G.; Meng, Q.; Finley, T.; Wang, T.; Chen, W.; Ma, W.; Ye, Q.; Liu, T.Y. LightGBM: A highly efficient gradient boosting decision tree. In Proceedings of the 31st International Conference on Neural Information Processing Systems, Long Beach, CA, USA, 4–9 December 2017; pp. 3149–3157. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | Influencing Factors | Mean | Std | Min | Max |
|---|---|---|---|---|---|
| Dependent variable | Stand productivity (m3ha−1year−1) | 4.9090 | 2.2403 | 0.2475 | 13.0550 |
| Species diversity | SIMP (unitless) | 0.7590 | 0.1385 | 0.1548 | 0.9422 |
| Structure diversity | NC (unitless) | 0.4406 | 0.0749 | 0.0000 | 0.5543 |
| Spatial distribution diversity | ASI (unitless) | 0.5412 | 0.0340 | 0.4427 | 0.6863 |
| Stand density | BA (m2ha−1) | 14.8948 | 9.2947 | 0.1669 | 58.8061 |
| Stand age | AGE (year) | 23.7913 | 8.6054 | 4.0000 | 55.0000 |
| Geographical condition | SLOP (degree) | 27.1391 | 9.3323 | 1.0625 | 54.2776 |
| ELEV (m) | 489.2547 | 313.8188 | 21.5299 | 1546.7600 | |
| ASPE | 2.5140 | 1.1285 | 1.0000 | 4.0000 | |
| Meteorological condition | AMT (°C) | 15.9507 | 1.4851 | 9.8313 | 19.5735 |
| AP (mm) | 1782.0610 | 245.1971 | 1287.0200 | 2365.7500 | |
| ISOT (unitless) | 25.7051 | 1.6273 | 19.1231 | 29.9280 | |
| TS (Std. 0.01 °C) | 796.8296 | 45.1620 | 689.4220 | 887.6100 | |
| PWQ (mm) | 555.8162 | 84.6708 | 385.0000 | 775.0000 |
| Model | R2 | RMSE (m3ha−1year−1) | rRMSE (%) |
|---|---|---|---|
| RF | 0.4763 | 1.6144 | 32.9008 |
| GBR | 0.4459 | 1.6572 | 33.7654 |
| XGBoost | 0.3794 | 1.7559 | 35.7458 |
| CatBoost | 0.4568 | 1.6423 | 33.4356 |
| Light GBM | 0.4613 | 1.6363 | 33.3490 |
| SVM | 0.4731 | 1.6163 | 32.8809 |
| Stacking | 0.4698 | 1.6219 | 33.0079 |
| Voting | 0.4855 | 1.5992 | 32.5715 |
| Autogluon | 0.5039 | 1.5710 | 31.9992 |
| LR | 0.3892 | 1.7416 | 35.4517 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Q.; Zhu, C.; Ji, B.; Xu, S.; Xie, B.; Wang, J.; Wang, Z. Quantification of the Influencing Factors of Stand Productivity of Subtropical Natural Broadleaved Forests in Eastern China Using an Explainable Machine Learning Framework. Forests 2025, 16, 95. https://doi.org/10.3390/f16010095
Du Q, Zhu C, Ji B, Xu S, Xie B, Wang J, Wang Z. Quantification of the Influencing Factors of Stand Productivity of Subtropical Natural Broadleaved Forests in Eastern China Using an Explainable Machine Learning Framework. Forests. 2025; 16(1):95. https://doi.org/10.3390/f16010095
Chicago/Turabian StyleDu, Qun, Chenghao Zhu, Biyong Ji, Sen Xu, Binglou Xie, Jianwu Wang, and Zhengyi Wang. 2025. "Quantification of the Influencing Factors of Stand Productivity of Subtropical Natural Broadleaved Forests in Eastern China Using an Explainable Machine Learning Framework" Forests 16, no. 1: 95. https://doi.org/10.3390/f16010095
APA StyleDu, Q., Zhu, C., Ji, B., Xu, S., Xie, B., Wang, J., & Wang, Z. (2025). Quantification of the Influencing Factors of Stand Productivity of Subtropical Natural Broadleaved Forests in Eastern China Using an Explainable Machine Learning Framework. Forests, 16(1), 95. https://doi.org/10.3390/f16010095