



Woody Species Composition, Stand Structure and Regeneration Status of Londiani Forest in Kenya

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Research Design
2.3. Data Collection
2.4. Data Analysis and Presentation
- 1.
- 2.
- 3.
- 4.
- Species Diversity
- 5.
- Evenness
- 6.
- Jaccard Similarity Index
- 7.
- The Importance Value Index (IVI) was used to determine the ecological importance of each tree species.
3. Results
3.1. Forest Stand Structure and Composition of Londiani Forest
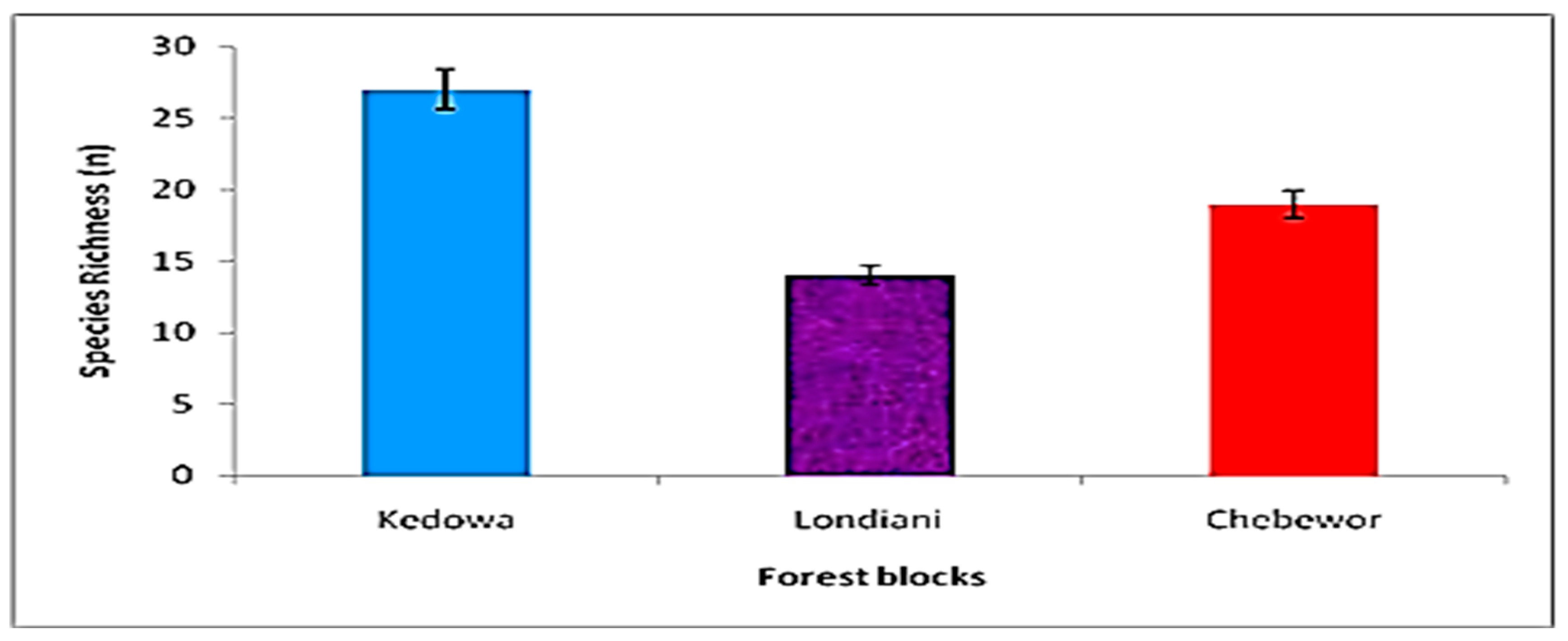
Woody Species Richness and Importance Value Indices
3.2. Diameter at Breast Height Distribution
3.3. Tree Height Distribution
3.4. Density and Relative Density
3.5. Basal Area, Dominance and Relative Dominance
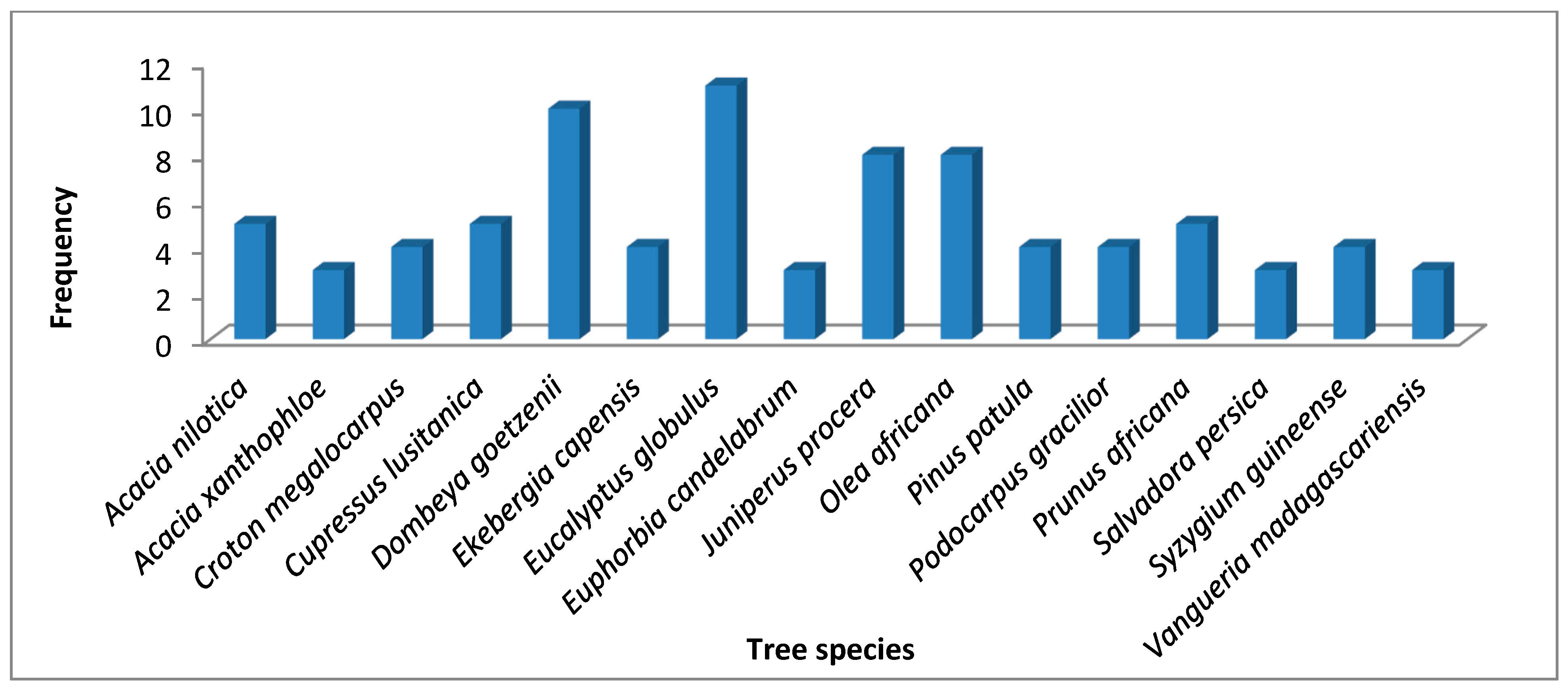
3.6. Frequency and Relative Frequency
3.7. Woody Species Diversity and Evenness
3.8. Similarity between Sites
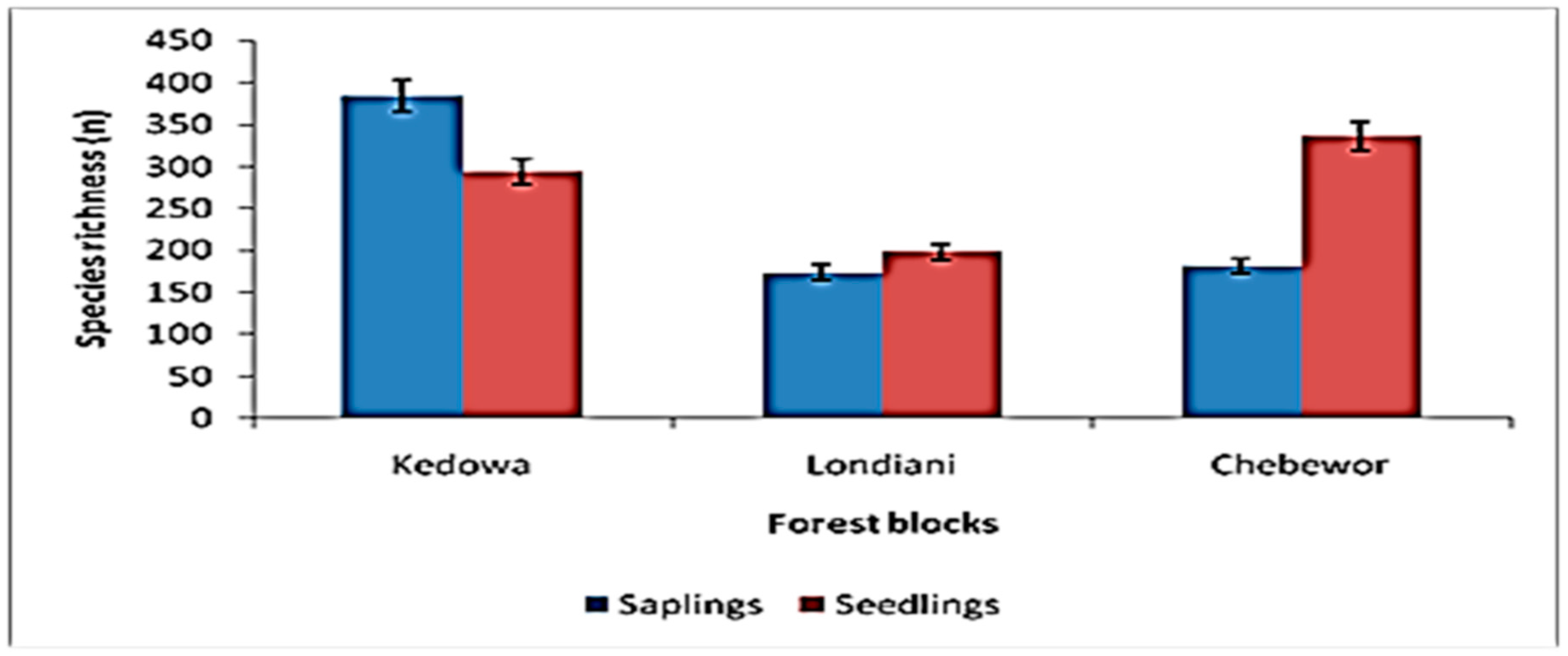
3.9. Regeneration Status of Londiani Forest
4. Discussion
4.1. Forest Stand Structure and Species Composition
4.2. Population Structure and Regeneration Status of Londiani Forest
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, G. Integrating Biodiversity Conservation and Sustainable Use: Lessons Learned from Ecological Networks; IUCN: Gland, Switzerland, 2004. [Google Scholar]
- Bharucha, Z.; Pretty, J. The roles and values of wild foods in agricultural systems. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2913–2926. [Google Scholar] [CrossRef]
- Blackman, A.; Veit, P. Titled Amazon indigenous communities cut forest carbon emissions. Ecol. Econ. 2018, 153, 56–67. [Google Scholar]
- Agevi, H.; Wabusya, M.; Tsingalia, H.M. Community Forest Associations and Community-Based Organizations: Redesigning their Roles in Forest Management and Conservation in Kenya. Int. J. Sci. Res. (IJSR) 2014, 3. [Google Scholar]
- Chomba, B.M.; Tembo, O.; Mutandi, K.; Mtongo, C.S.; Makano, A. Drivers of Deforestation, Identification of Threatened Forests and Forest Co-Benefits Other than Carbon from REDD+ Implementation in Zambia; A consultancy report prepared for the Forestry Department and the Food and Agriculture Organization of the United Nations under the national UN-REDD Programme. Ministry of Lands, Natural Resources and Environmental Protection: Lusaka, Zambia, 2014. Available online: http://landforlions.org/data/documents/drivers-deforestation-Zambia-WEB_final.pdf (accessed on 22 September 2021).
- Beatty, C.R.; Cox, N.A.; Kuzee, M.E. Biodiversity Guidelines for Forest Landscape Restoration Opportunities Assessments, 1st ed.; IUCN: Gland, Switzerland, 2018. [Google Scholar]
- Bastin, J.-F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The global tree restoration potential. Science 2019, 365, 76–79. [Google Scholar] [PubMed]
- Zhu, H.; Xu, Z.F.; Wang, H.; Li, B.G. Tropical rain forest fragmentation and its ecological and species diversity changes in southern Yunnan. Biodivers. Conserv. 2004, 13, 1355–1372. [Google Scholar] [CrossRef]
- Agrawal, A.; Chhatre, A.; Hardin, R. Changing governance of the world’s forests. Science 2008, 320, 1460–1462. [Google Scholar] [CrossRef]
- Chan, K.M.A.; Pringle, R.M.; Ranganathan, J.; Boggs, C.L.; Chan, Y.L.; Ehrlich, P.R.; Haff, P.K.; Heller, N.E.; Al-Khafaji, K.; Macmynowski, D.P. When agendas collide: Human welfare and biological conservation. Conserv. Biol. 2007, 21, 59–68. [Google Scholar] [CrossRef]
- Ahenkan, A.; Boon, E. Improving nutrition and health through non-timber forest products in Ghana. J. Health Popul. Nutr. 2011, 29, 141–148. [Google Scholar] [CrossRef]
- Barros, F.M.; Peres, C.A.; Pizo, M.A.; Ribeiro, M.C. Divergent flows of avian-mediated ecosystem services across forest-matrix interfaces in human-modified landscapes. Landsc. Ecol. 2019, 35, 879. [Google Scholar] [CrossRef]
- CBD. Linking Biodiversity Conservation and Poverty Alleviation: A State of Knowledge Review; CBD Technical Series No: 55; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 2010. [Google Scholar]
- Ding, H.; Veit, P.G.; Blackman, A.; Gray, E.; Reytar, K.; Altamirano, J.C.; Hodgdon, B. Climate Benefits, Tenure Costs: The Economic Case for Securing Indigenous Land Rights in the Amazon; WRI: Washington, DC, USA, 2016. [Google Scholar]
- Wenger, K.F. Forestry Handbook; John Wiley and Sons, Inc.: New York, NY, USA, 2013; 1335p. [Google Scholar]
- Dau, J.H.; Mati, A.; Dawaki, S.A. Role of Forest Inventory in Sustainable Forest Management: A Review. Int. J. For. Hortic. 2015, 1, 33–40. [Google Scholar]
- Monserud, R.A. Evaluating Forest Models in a Sustainable Forest Management Context; FBMIS, 2003; Volume 1, pp. 35–47. ISSN 1740-5955. Available online: http://www.fbmis.info/A/3_1_MonserudR_1 (accessed on 15 May 2023).
- Zerihun, A.; Yemir, T. Hawassa University: Wondo Genet Colleg of Forestry and Natural Resources. In Training Manual on: Forest Inventory and Management in the Context of SFM & REDD; International Journal of Forestry and Horticulture (IJFH) Volume 1, Issue 2, July–September 2015, PP 33-40 ISSN 2454-9487. Available online: www.arcjournals.org (accessed on 15 May 2023).
- Mendoza, G.A.; Prabhu, R. Qualitative multi-criteria approach to assessing indicators of sustainable forest resource management. For. Ecol. Manag. 2000, 131, 107–126. [Google Scholar] [CrossRef]
- Marengo, J. Drought, Floods, Climate Change, and Forest Loss in the Amazon Region: A Present and Future Danger? Front. Young Minds 2020, 8, 147. [Google Scholar] [CrossRef]
- Lawrence, D.; Vandecar, K. Effects of tropical deforestation on climate and agriculture. Nat. Clim. Chang. 2015, 5, 27–36. [Google Scholar] [CrossRef]
- Krupnick, G.A. Conservation of tropical plant biodiversity: What have we done, where are we going? Wiley Online libr. Biotropica 2013, 45, 693–708. [Google Scholar]
- Fischer, R.; Bohn, F.; de Paula, M.D.; Dislich, C.; Groeneveld, J.; Gutiérrez, A.G.; Kazmierczak, M.; Knapp, N.; Lehmann, S.; Paulick, S.; et al. Lessons learned from applying a forest gap model to understand the ecosystem and carbon dynamics of complex tropical forests. Ecol. Modell. 2016, 326, 124–133. [Google Scholar] [CrossRef]
- Aide, T.M. Tropical Forest Restoration; Restoration Ecology Special Issue; Encyclopedia of Life Support Systems (EOLSS); 2000; Volume 8, pp. 327–424. Available online: https://www.eolss.net/ebooklib/sc_cart.aspx (accessed on 18 June 2022).
- Chazdon, R.L.; Uriarte, M. Natural regeneration in the context of large-scale forest and landscape restoration in the tropics. Biotropica 2016, 48, 709–715. [Google Scholar] [CrossRef]
- Wangda, P. Forest Zonation along the Complex Altitudinal Gradients in a Dry Valley of Punatsang Chu. Master’s Thesis, Graduate School of Frontier Sciences, Laboratory of Biosphere Functions, University of Tokyo, Tokyo, Japan, 2003. [Google Scholar]
- Mori, A.; Takeda, H. Effects of Undisturbed Canopy Structure on Population Structure and Species Coexistence in an Old-growth Subalpine Forest in Central Japan. For. Ecol. Manag. 2004, 200, 89–100. [Google Scholar] [CrossRef]
- Peña-Claros, M.; Poorter, L.; Alarcón, A.; Blate, G.; Choque, U.; Fredericksen, T.S.; Justiniano, M.J.; Leaño, C.; Licona, J.C.; Pariona, W. Soil Effects on Forest Structure and Diversity in a Moist and a Dry Tropical Forest. Biotropica 2012, 44, 276–283. [Google Scholar] [CrossRef]
- Liira, J.; Sepp, T.; Kohv, K. The ecology of tree regeneration in mature and old forests: Combined knowledge for sustainable forest management. J. For. Res. 2011, 16, 184–193. [Google Scholar] [CrossRef]
- Chazdon, R.L. Tropical forest recovery: Legacies of human impact and natural disturbances. In Perspectives in Plant Ecology, Evolution and Systematics; Urban & Fischer Verlag: Munich, Germany, 2003; Volume 6, pp. 51–71. ISSN 1433-8319. [Google Scholar]
- Bazzaz, F. Regeneration of Tropical Forests: Physiological Responses of Pioneer and Secondary Species. In Rainforest Regeneration and Management; Gomez-pompa, A., Whitmore, T., Hadley, M., Eds.; Parthenon Publishing, UNES: Paris, France, 1991; pp. 91–118. [Google Scholar]
- Khumbongmayum, A.D.; Khan, M.; Tripathi, R. Biodiversity Conservation in Sacred Groves of Manipur, Northeast India: Population Structure and Regeneration Status of Woody Species. In Human Exploitation and Biodiversity Conservation; Springer: Berlin/Heidelberg, Germany, 2006; Volume 15, pp. 2439–2456. [Google Scholar]
- Strassburg, B.B.N.; Beyer, H.L.; Crouzeilles, R.; Iribarrem, A.; Barros, F.; de Siqueira, M.F.; Sánchez-Tapia, A.; Balmford, A.; Sansevero, J.B.B.; Brancalion, P.H.S.; et al. Strategic approaches to restoring ecosystems can triple conservation gains and halve costs. Nat. Ecol. Evol. 2019, 3, 62–70. [Google Scholar] [CrossRef]
- Teketay, D. Seed and Regeneration Ecology in Dry Afromontane Forests of Ethiopia: I. Seed Production-population Structures. Trop. Ecol. 2005, 46, 29–44. [Google Scholar]
- Wittmann, F.; Schöngart, J.; Montero, J.C.; Motzer, T.; Junk, W.J.; Pieadade, M.T.F.; Queiroz, H.L.; Worbes, M. Tree Species Composition and Diversity Gradients in the White-Water Forests across the Amazon Basin. J. Biogeogr. 2006, 33, 1334–1347. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Chao, A. Measuring and Estimating Species Richness, Species Diversity, and Biotic Similarity from Sampling Data, 2nd ed.; Levin, S.A., Ed.; Encyclopedia of Biodiversity; Academic Press: Waltham, MA, USA, 2013; Volume 5, pp. 195–211. [Google Scholar]
- Schleuning, M.; Farwig, N.; Peters, M.K.; Bergsdorf, T.; Bleher, B.; Brandl, R.; Dalitz, H.; Fischer, G.; Freund, W.; Gikungu, M.W.; et al. Forest fragmentation and selective logging have inconsistent effects on multiple animal-mediated ecosystem processes in a tropical forest. PLoS ONE 2011, 6, e27785. [Google Scholar] [CrossRef] [PubMed]
- Tarus, G.K.; Nadir, S.W. Effect of Forest Management Types on Soil Carbon Stocks in Montane Forests: A Case Study of Eastern Mau Forest in Kenya. Int. J. For. Res. 2020, 8862813. [Google Scholar] [CrossRef]
- [GoK] Government of Kenya. Ministry of livestock and fisheries. Agricultural Sector Development Support System (ASDSP) Volume I Household Baseline Survey Report- Kericho County 2014. Available online: www.asdsp.co.ke (accessed on 5 July 2022).
- Government of Kenya (GOK). Sessional Paper No. 1 of 2007 on Forest Policy; Government of Kenya Printers: Nairobi, Kenya, 2007.
- Government of Kenya (GOK). Kenya Gazette Supplement Acts: The Forest Act; Government of Kenya printers: Nairobi, Kenya, 2005.
- Kenya Forest Service. Londiani Participatory Forest Management Plan 2018–2022; CIFOR: Bogor, Indonesia, 2018. [Google Scholar]
- GOK. Plantation Establishment for Livelihood Improvement Scheme Guidelines; Government Printers: Nairobi, Kenya, 2007. [Google Scholar]
- Richardson, D.M.; Pysek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 3, 14–93. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Pyšek, P.; Bacher, S.; Carlton, J.T.; Duncan, R.P.; Jarošík, V.; Wilson, J.R.; Richardson, D.M. A proposed unified framework for biological invasions. Trends Ecol. Evol. 2011, 26, 333–339. [Google Scholar] [CrossRef]
- Roswell, M.; Dushoff, J.; Winfree, R. A Conceptual Guide to Measuring Species Diversity. Oikos Adv. Ecol. 2021, 130, 321–338. [Google Scholar] [CrossRef]
- Hancock, P.R. Book Reviews. Rev. Radic. Political Econ. 2004, 36, 260–263. [Google Scholar] [CrossRef]
- Mutiso, F.; Hitimana, J.; Kiyiapi, J.; Sang, F.; Eboh, E. Recovery of Kakamega tropical rainforest from anthropogenic disturbances. J. Trop. For. Sci. 2013, 25, 566–576. [Google Scholar]
- Girma, A.; Mosandl, R. Structure and potential regeneration of degraded secondary stands in Sameness-Shashemene Forest, Ethiopia. J. Trop. For. Sci. 2012, 24, 46–53. [Google Scholar]
- Teketay, D.; Kashe, K.; Madome, J.; Kabelo, M.; Neelo, J.; Mmusi, M.; Masamba, W. Enhancement of Diversity, Stand Structure and Regeneration of Woody Species through Area Exclosure: The Case of a Mopane Woodland in Northern Botswana. Ecol. Process. 2018, 7, 5. [Google Scholar] [CrossRef]
- Mutiso, F.M.; Mugo, M.J.; Cheboiwo, J.; Sang, F.; Tarus, G.K. Floristic Composition, Affinities and Plant Formations in Tropical Forests: A Case Study of Mau Forests in Kenya. Int. J. Agric. For. 2015, 5, 79–91. [Google Scholar] [CrossRef]
- Getaneh, G.; Teshome, S.; Tesfaye, B.; Demel, T. Species composition, stand structure, and regeneration status of tree species in dry Afromontane forests of Awi Zone, northwestern Ethiopia. Ecosyst. Health Sustain. 2019, 5, 199–215. [Google Scholar] [CrossRef]
- Kinyanjui, M.J.; Latva-Käyrä, P.; Bhuwneshwar, P.S.; Kariuki, P.; Gichu, A.; Wamichwe, K. An Inventory of the Above Ground Biomass in the Mau Forest Ecosystem, Kenya. Open J. Ecol. 2014, 4, 619–627. [Google Scholar] [CrossRef]
- Sapkota, I.; Tigabu, M.; Oden, P. Changes in tree species diversity and dominance across disturbance gradient in Napelese Sal (Shorea robusta Gaertn. f.). J. For. Res. 2010, 21, 25–32. [Google Scholar] [CrossRef]
- Kuria, N.C.; Balozi, K.; Kipkore, W. Growth and Yield Models for Plantation-Grown Cupressus lusitanica for Central Kenya. Afr. J. Educ. Sci. Technol. 2019, 5, 34–58. [Google Scholar]
- Mutangah, J.; Mwangangi, O.; Mwaura, P. Mau Forest Complex Vegetation. J. Ecol. 2014, 04, 619–627. [Google Scholar] [CrossRef]
- Omondi, F.S. Early Growth Performance and Survival of Pine Hybrids in Turbo, Kenya. East Afr. Agric. For. J. 2018, 84. Available online: https://www.kalro.org/www.eaafj.or.ke/index.php/path/article/view/301/475 (accessed on 5 July 2022).
- Agevi, H. Determination of Species Abundance, Diversity and Carbon Stocks in Kakamega Forest Ecosystem. Ph.D. Dissertation, Moi University, Eldoret, Kenya, 2020. Available online: http://ir.mu.ac.ke:8080/jspui/handle/123456789/3101 (accessed on 5 July 2022).
- Cheboiwo, K.; Joshua, M.; Robert, O.; Mbinga, J.; Festus, M. Potential Growth, Yields and Socioeconomic benefits of Four Indigenous Species for Restoration in Moist Forest, Mau Kenya. J. Environ. Earth Sci. 2015, 5. ISSN 2224-3216 (Paper) ISSN 2225-0948 (Online). Available online: www.iiste.org (accessed on 5 July 2022).
- KEFRI. National Tree Improvement Strategy 2018–2022; KEFRI: Muguga, Kenya, 2018. [Google Scholar]
- Gaaf, N.; Poels, H.; Rompaey, R. Effect of silvicultural treatment on growth and mortality of rainforest in Suriname over long periods. For. Ecol. Manag. 1999, 124, 123–135. [Google Scholar]
- Kagombe, J.K. Contribution of PELIS in Increasing Tree Cover and Community Livelihoods; KEFRI: Nairobi, Kenya, 2014; Available online: www.kefri.org (accessed on 5 July 2022).
- Gichuru, I. An Evaluation of Factors Contributing to the Success of PELIS Strategy in Forest Plantation Establishment in Mucheene Forest. Int. J. Soc. Sci. Proj. Plan. Manag. 2015, 1, 15–43. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Block | Abundance | Richness | Diversity (H′) | Evenness (H/Hmax) |
|---|---|---|---|---|
| Kedowa | 76 ± 0.033 | 6 ± 0.5 | 1 ± 0.14 | 0.57 ± 0.1 |
| Chebewor | 71 ± 0.4 | 7 ± 0.01 | 1 ± 0.64 | 1 ± 0.32 |
| Londiani | 72 ± 0.3 | 4 ± 0.21 | 0.77 ± 0.01 | 0.41 ± 0.04 |
| Forest | Jaccard’s Similarity Coefficient (S) | ||
|---|---|---|---|
| Kedowa | Chebewor | Londiani | |
| Chebewor | 0.47 | ||
| Londiani | 0.35 | ||
| Kedowa | 0.34 | ||
| Forest Blocks | Abundance | Richness | Diversity H’ | |||
|---|---|---|---|---|---|---|
| Saplings | Seedlings | Saplings | Seedlings | Saplings | Seedlings | |
| Kedowa | 384 | 294 | 17 | 22 | 2.01 | 1.62 |
| Chebewor | 182 | 337 | 12 | 9 | 2.89 | 2.58 |
| Londiani | 174 | 198 | 8 | 7 | 1.64 | 1.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chepkoech, E.K.; Agevi, H.; Lung’ayia, H.; Tsingalia, H.M. Woody Species Composition, Stand Structure and Regeneration Status of Londiani Forest in Kenya. Forests 2024, 15, 653. https://doi.org/10.3390/f15040653
Chepkoech EK, Agevi H, Lung’ayia H, Tsingalia HM. Woody Species Composition, Stand Structure and Regeneration Status of Londiani Forest in Kenya. Forests. 2024; 15(4):653. https://doi.org/10.3390/f15040653
Chicago/Turabian StyleChepkoech, Evalyne Kosgey, Humphrey Agevi, Henry Lung’ayia, and Harrison Mugatsia Tsingalia. 2024. "Woody Species Composition, Stand Structure and Regeneration Status of Londiani Forest in Kenya" Forests 15, no. 4: 653. https://doi.org/10.3390/f15040653
APA StyleChepkoech, E. K., Agevi, H., Lung’ayia, H., & Tsingalia, H. M. (2024). Woody Species Composition, Stand Structure and Regeneration Status of Londiani Forest in Kenya. Forests, 15(4), 653. https://doi.org/10.3390/f15040653