
Optimizing Pinus tabuliformis Forest Spatial Structure and Function in Beijing, China

Abstract
1. Introduction
2. Materials and Methods
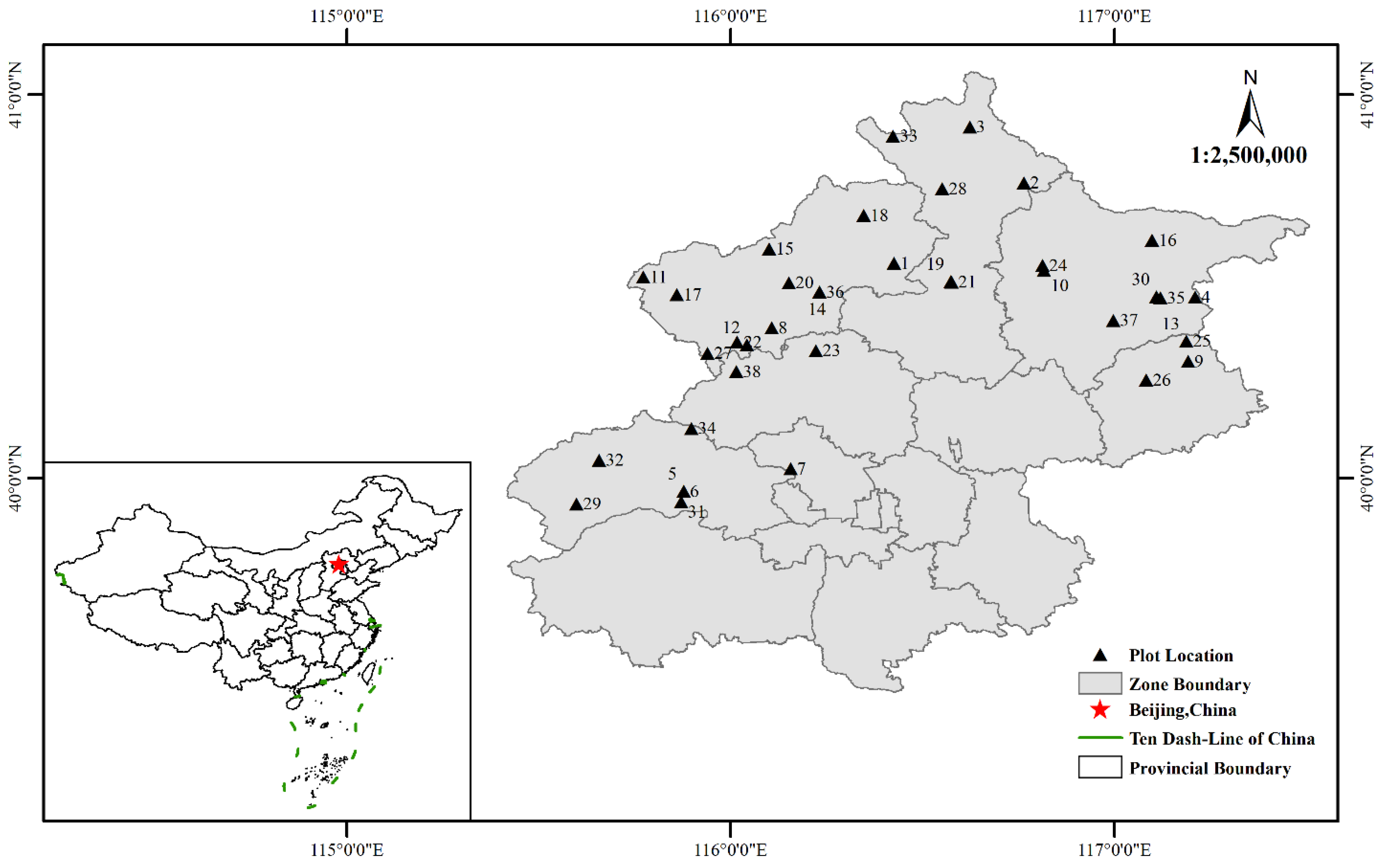
2.1. Study Area Overview
2.2. Sample Plot Settings
2.3. Research Methods
2.3.1. Stand Structure Parameters
- 1.
- The uniform angle index reflects the horizontal spatial distribution of the trees [19], as follows:where Wi is the uniform angle of the target tree i and n is the number of adjacent trees. When the angle of the adjacent trees is less than the standard angle (360°/[n + 1]), Zij = 1; otherwise, Zij = 0.
- 2.
- The neighborhood comparison index reflects the degree of differentiation of a target tree (in terms of factors such as DBH, tree height, and crown width) [20], as follows:where Ui is the neighborhood comparison index of target tree i and n is the number of adjacent trees. When the DBH of target tree i is smaller than that of the adjacent tree j, Kij = 0; otherwise, Kij = 1.
- 3.
- The mingling degree reflects the degree of isolation among the trees [21], as follows:where Mi is the mingling degree of target tree i and n is the number of adjacent trees. When the target tree i is not of the same species as the adjacent tree j, Vij = 1; otherwise, Vij = 0.
- 4.
- The openness degree reflects the light-transmission conditions in the forest [22], as follows:where Ki is the openness degree of the target tree i, and n is the number of adjacent trees. Dij is the horizontal distance between object tree i and adjacent tree j, and Hij is the height of adjacent tree j.
- 5.
- The competition index, which reflects the relationship between individual tree growth and spatial occupation, utilizes Hegyi’s diameter–distance competition index [23], as follows:where is the competition index of the target tree, di represents the DBH of the target tree, dj is the DBH of its immediate neighbors, is the distance between i and j, and ni is the number of trees adjacent to the target tree i.
- 6.
- The forest layer index represents the structural vertical complexity of a stand [24], as follows:where is the forest layer index of the target tree, is the number of forest layers of target tree i, and is the value of the forest layer. When target tree i and the adjacent tree j do not belong to the same layer, = 1; otherwise, = 0.
2.3.2. Understory Herbaceous Plant Diversity Indices
- Gleason index:
- Margalef richness index:
- Simpson index:
- Shannon–Wiener index:
- Pielou evenness index:where A is the total area surveyed, S is the total number of species, N is the number of individuals of all species, H is the species richness of various species in the habitat, Pi is the proportion of species in the total community, and Ni is the number of individuals of the species.
2.3.3. Quantification of Soil Nutrients
2.3.4. Data Processing
3. Results
3.1. Spatial Structure and Function Feature Analysis
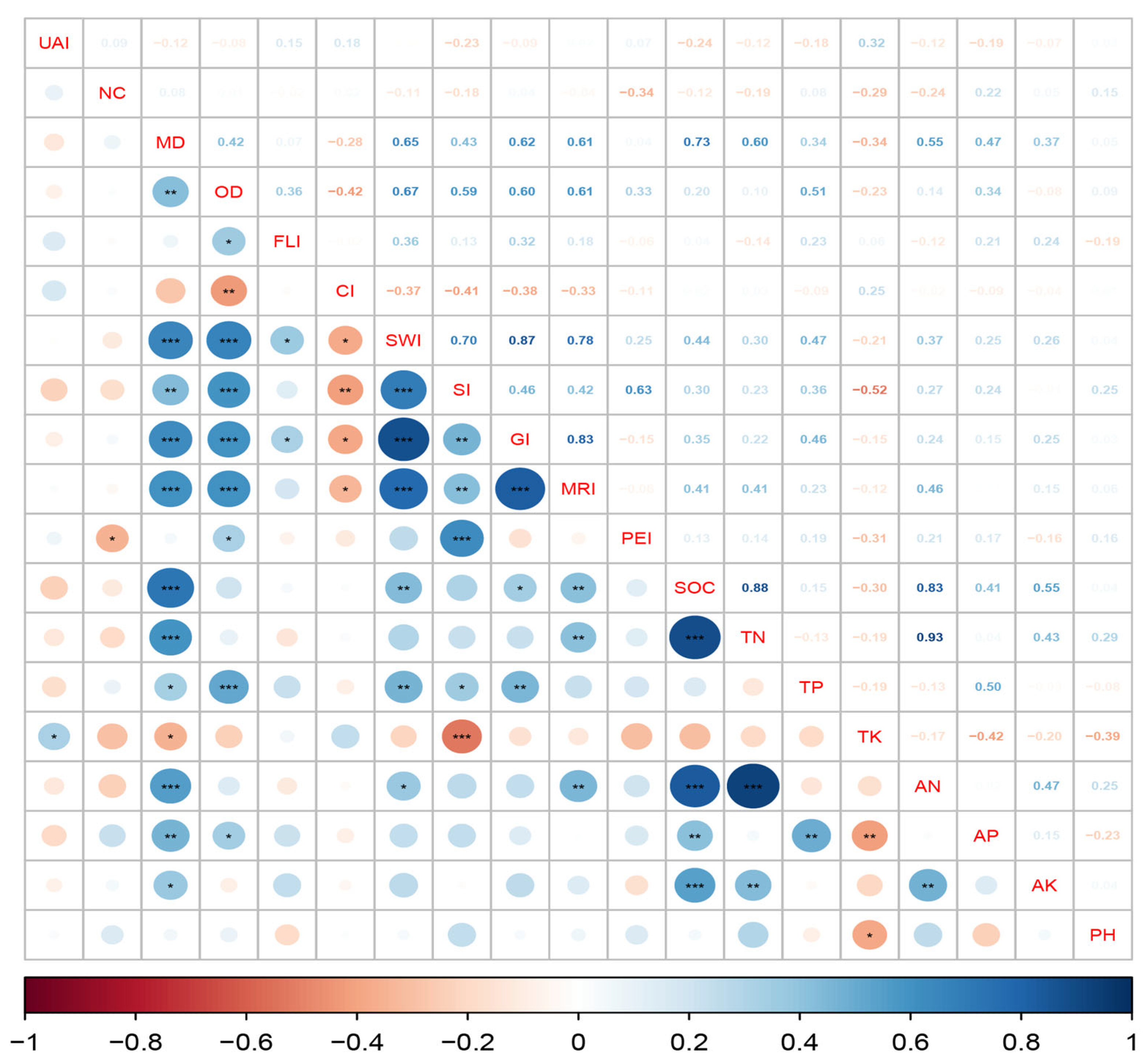
3.2. Correlations Between Spatial Structure and Understory Diversity and Soil Nutrient Levels
3.3. SEM of Effects of Stand Spatial Structure on Understory Plant Diversity and Soil Nutrient Levels
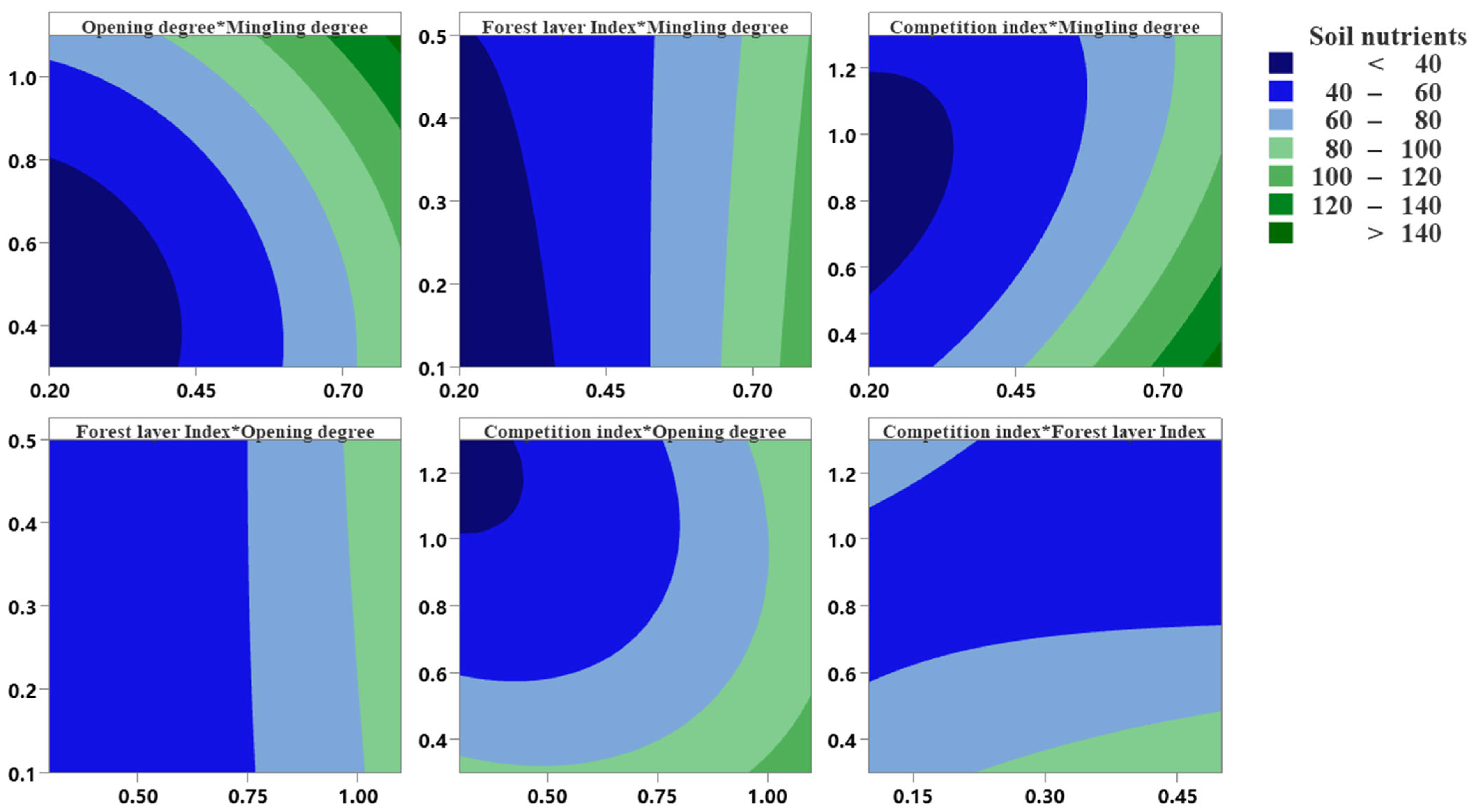
3.4. Stand Structural Optimization and Adjustment
4. Discussion
4.1. Correlations Between Stand Spatial Structure and Herbaceous Understory Diversity
4.2. Correlations Between Stand Spatial Structure and Soil Nutrient Levels
4.3. Coupling Between Stand Spatial Structure and Ecological Function
4.4. Stand Spatial Structure Optimization Strategy
4.5. Research Limitations and Future Trends
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spies, T.A. Forest Structure: A Key to the Ecosystem. Northwest Sci. 1998, 7, 34–39. [Google Scholar]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Palik, B.J.; Dumroese, R.K. Contemporary forest restoration: A review emphasizing function. For. Ecol. Manag. 2014, 331, 292–323. [Google Scholar] [CrossRef]
- Ali, A. Forest stand structure and functioning: Current knowledge and future challenges. Ecol. Indic. 2019, 98, 665–677. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Cao, X.; Liu, Z.; Lv, Y. The Multivariate Distribution of Stand Spatial Structure and Tree Size Indices Using Neighborhood-Based Variables in Coniferous and Broad Mixed Forest. Forests 2023, 14, 2228. [Google Scholar] [CrossRef]
- Zhang, H.; Lei, X.; Zhang, C.; Zhao, X.; Hu, X. Research on theory and technology of forest quality evaluation and precision improvement. J. Beijing For. Univ. 2019, 41, 1–18. [Google Scholar] [CrossRef]
- Zhu, G.; Xu, Q.; Lu, Y. Effects of stand spatial structure on species diversity of shrubs in Quercus spp. natural secondary forests in Hunan Province. Acta Ecol. Sin. 2018, 38, 5404–5412. [Google Scholar]
- Fan, B. A Study on the Stand Spatial Structure and Soil Physicochemical Properties of Natural Secondary Broad-Leaved Forest. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2012. [Google Scholar]
- Cao, X.; Li, J.; Wei, X. Effects of Spatial Structure on Soil Nutrient Content in Typical Forests in the Contral-Subtropics of China. Sci. Silvae Sin. 2020, 56, 20–28. [Google Scholar] [CrossRef]
- Zhu, J.; Zhan, B.; Chen, X.; Guan, Q. Analysis of spatial structure and species diversity of coniferous and broadleaved mixed forest in Zijin mountain. J. Cent. South Univ. For. Technol. 2018, 38, 75–80+93. [Google Scholar] [CrossRef]
- Pang, Y. Soil Fertility Analysis and Comprehensive Evaluation of Four Typical Stands in Fushou Forest Farm. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2022. [Google Scholar]
- Huang, X. Research on the spatial structure and multifunctionality of Pinus massoniana plantations in Jindong Forest Farm. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2017. [Google Scholar]
- Sun, Y.; Li, J.; Cao, X.; Yan, J. Research on the relationship between spatial structure and soil nutrients of Cunninghamia lanceolata forest based on path analysis method. J. Cent. South Univ. For. Technol. 2020, 40, 41–47. [Google Scholar] [CrossRef]
- Fang, X.; Tan, W.; Gao, X.; Chai, Z. Close-to-nature management positively improves the spatial structure of Masson pine forest stands. Web Ecol. 2021, .21, 45–54. [Google Scholar] [CrossRef]
- Tang, M.; Tang, S.; Lei, X.; Li, X. Study on Spatial Structure Optimizing Model of Stand Selection Cutting. Sci. Silvae Sin. 2004, 40, 25–31. [Google Scholar]
- Hu, Y.; Hui, G. A Discussion on Forest Management Method Optimizing Forest Spatial Structure. For. Res. 2006, 19, 1–8. [Google Scholar]
- Liang, W.; Wei, X. Relationships between ecosystems above and below ground including forest structure, herb diversity and soil properties in the mountainous area of Northern China. Glob. Ecol. Conserv. 2020, 24, e01228. [Google Scholar] [CrossRef]
- Hui, G.; Zhang, G.; Zhao, Z.; Yang, A. Methods of Forest Structure Research: A Review. Curr. For. Rep. 2019, 5, 142–154. [Google Scholar] [CrossRef]
- Hui, G.; Gadow, K.V.; Albert, M. The Neighbourhood Pattern—A New Structure Parameter for Describing Distribution of Forest Tree Position. Sci. Silvae Sin. 1999, 35, 39–44. [Google Scholar]
- Hui, G.; Gadow, K.V.; Albert, M.T. A New Parameter for Stand Spatial Structure Neighbourhood Comparison. For. Res. 1999, 12, 1–6. [Google Scholar]
- Tang, M.; Tang, S.; Lei, X.; Zhou, G.; Xie, Z. Comparison Analysis on Two Minglings. For. Grassl. Resour. Res. 2004, 25–27. [Google Scholar] [CrossRef]
- Wang, P.; Jia, L.; Wei, S.; Wang, Q. Analysis of stand spatial structure of Platycladus orientalis recreational forest based on Voronoi diagram method. J. Beijing For. Univ. 2013, 35, 39–44. [Google Scholar] [CrossRef]
- Hui, G.; Hu, Y.; Zhao, Z.; Yuan, S.; Liu, W. A Forest Competition Index Based on Intersection Angle. Sci. Silvae Sin. 2013, 49, 68–73. [Google Scholar] [CrossRef]
- Lu, Y.; Zang, H.; Wan, X.; Deng, Z.; Li, J. Storey Structure Study of Cyclobalanopsis myrsinaefolia Mixed Stand Based on Storey Index. For. Grassl. Resour. Res. 2012, 81–84. [Google Scholar] [CrossRef]
- Peng, Y.; Fan, M.; Song, J.; Cui, T.; Li, R. Assessment of plant species diversity based on hyperspectral indices at a fine scale. Sci. Rep. 2018, 8, 4776. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Bai, Y.; Yang, L. Efficient Soil Nutrient Testing Technologies and Equipment; China Agricultural Press: Beijing, China, 2006. [Google Scholar]
- Henseler, J.; Ringle, C.M.; Sarstedt, M. A new criterion for assessing discriminant validity in variance-based structural equation modeling. J. Acad. Mark. Sci. 2014, 43, 115–135. [Google Scholar] [CrossRef]
- Hou, G. Optimizition of Stand Structures for Low-efficiency Robinia pseudoacacia Forest in the Loess Plateau in Western Shanxi, China. Doctoral Thesis, Beijing Forestry University, Beijing, China, 2020. [Google Scholar]
- Yang, Y.; Shi, Y.; Li, Z.; Kang, H.; Fan, Y.; Zheng, C.; Wen, Z. Community characteristics of understory herb layer and its relationships with stand structure and soil nutrient availability in the Three-North shelterbelt of Shaanxi, China. Acta Ecol. Sin. 2020, 40, 6542–6551. [Google Scholar]
- Zhang, Y.-H.; Dian, Y.-Y.; Huang, G.; Liu, X.; Han, Z.; Jian, Y.; Li, Y.; Wang, X. Effects of spatial structure on species diversity in Pinus massoniana plantation of different succession stages. Chin. J. Ecol. 2021, 40, 2357–2365. [Google Scholar]
- Cao, X.; Li, J.; Wei, X. Analysis of the difference and correlation between the spatial structure and understory herbaceous species diversity of typical subtropical forests. Pratacultural Sci. 2019, 36, 2466–2475. [Google Scholar] [CrossRef]
- Li, F.; Pan, P.; Ning, J.; Lai, G.; Ouyang, X.; Xu, H.; Guo, L.; Wu, Z.; Yi, Z. Effects of Stand Spatial Structure on Understory Vegetation Diversity of Aerial Seeding Pinus massoniana Plantations. J. Northeast For. Univ. 2016, 44, 31–35+40. [Google Scholar] [CrossRef]
- Xie, Z. Study on the Influence of the Spatial Structure of Chinese Fir Public Welfare Forest on the Species Diversity of Shrub and Herbaceous. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2023. [Google Scholar]
- Mao, Y.; Lin, H.; Sun, L.; Xie, J.; Jiang, T.; Wang, Y. Effects of Stand Spatial Structure on Understory Species Diversity of Cupressus funebris Plantation. Guangxi For. Sci. 2021, 50, 639–645. [Google Scholar] [CrossRef]
- Grime, J.P. Competitive Exclusion in Herbaceous Vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Hui, G.; Hu, Y.; Zhao, Z. Research Progress of Structure-based Forest Management. For. Res. 2018, 31, 85–93. [Google Scholar]
- Wang, N.; Bi, H.; Peng, R.; Zhao, D.; Yun, H.; Liu, Z.; Lan, D.; Jin, C. The Strategy for Optimizing the Stand Structure of Pinus tabuliformis Carr. Forests to Enhance the Ecological Function on the Loess Plateau, China. Forests 2022, 13, 1217. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, X.; Yue, Y.; Gan, J.; Wang, X.; Li, J. Spatial structure of Robinia pseudoacacia plantation in Miyun Reservoir Watershed of Beijing. J. Beijing For. Univ. 2009, 31, 25–28. [Google Scholar]
- Xiang, Q.; Guuo, Q.; Ai, X.; Yao, L.; Zhu, J.; Xue, W.; Zuou, Y.; Zhao, H.; Wu, J. Variations on Stand Spatial Structure and Species Diversity in Different Spatial Scales. For. Res. 2022, 35, 151–160. [Google Scholar] [CrossRef]
- Huo, Y.; Guan, J.; Liu, X.; Zhao, W.; Li, S.; Deng, L. Interaction between fine roots of Picea crassifolia and soil microorganisms under different mingling degree stands. J. For. Environ. 2023, 43, 588–595. [Google Scholar]
- Jing, Y.; Wang, Q. Mixed Cunninghamia lanceolata–Michelia macclurei plantation decreased soil bacterial residue accumulation in topsoils. Chin. J. Ecol. 2024, 43, 2650–2657. [Google Scholar] [CrossRef]
- Zhao, W. Study on the Effect of Spatial Structure of Chinese Fir Public Welfare Forest on Soil Nutrients-Take Fushou Forest Farm as an Example. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2023. [Google Scholar]
- Zhang, M.; Yin, Y.; Wang, K.; Wang, S. Effects of spatial structure characteristics of Fraxinus mandshurica plantation on soil nutrient content. J. Beijing For. Univ. 2023, 45, 73–82. [Google Scholar] [CrossRef]
- Lv, K.; Zhang, E.; Li, S.; Jin, S.; Zhou, M.; Yan, D. Effects of stand spatial structure on understory plant diversity in Pinus taiwanensis plantation. J. Zhejiang AF Univ. 2022, 39, 1257–1266. [Google Scholar] [CrossRef]
- Prescott, C.E. The influence of the forest canopy on nutrient cycling. Tree Physiol. 2002, 22, 1193–1200. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Z.; Shen, L.; Zhao, Y.; Tang, Y.; Tian, J. Effects of Stand Structure of Artificial Shelter Forest on Understory Herb Diversity in Desert-Oasis Ecotone. Diversity 2023, 15, 1083. [Google Scholar] [CrossRef]
- Fiifibartels, S. Influence of Overstory Composition on Understory Vegetation and Resource Environment in the Boreal Forests of Canada. Master’s Thesis, Lakehead University, Ottawa, Canada, 2010. [Google Scholar]
- Peng, T. Differences in Spatial Structure, Understory Vegetation and Soil Characteristics of fir Large-Diameter Timber Plantations Under Different Methods. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2023. [Google Scholar]
- Liu, Y.; Wang, X.; He, L.; Liu, Z.; Zeng, X.; Sha, H.; He, B.; JIin, Y.; Li, J.; Chen, J.; et al. Effects of spatial structure on understory vegetation and soil properties in Pinus tabuliformis plantation of different succession types in Beijing. Acta Ecol. Sin. 2023, 43, 1959–1970. [Google Scholar]
- Lei, X.; Tang, S.; Li, D.; Chen, B.; Zelu, Z. Stand variables affecting understorey plant species diversity in natural forests. Chin. J. Ecol. 2003, 22, 18–22. [Google Scholar]
- Xingyun, L.; Youjun, H.; Pu, Z.; Jiantao, R.; Lin, Q.; Zhiyong, L. Effects of Different Forest Management Regimes on Plant Community Structure and Biodiversity of Natural Secondary Forests in Danqinghe Forestry Farm. Sci. Silvae Sin. 2013, 49, 93–102. [Google Scholar]
- Hu, W.; Wang, X.; Cui, H.; Pang, H.; Fu, T.; Pan, L.; Tang, W.; Meng, Y. Effects of stand structure and tree species composition on density of regeneration seedling of Pinus massoniana forest in Jiulingtou of Yichang City. J. Plant Resour. Environ. 2020, 29, 52–60. [Google Scholar]
- Fahey, R.T.; Fotis, A.T.; Woods, K.D. Quantifying canopy complexity and effects on productivity and resilience in late-successional hemlock-hardwood forests. Ecol. Appl. 2015, 25, 834–847. [Google Scholar] [CrossRef]
- Ediriweera, S.; Singhakumara, B.M.P.; Ashton, M.S. Variation in canopy structure, light and soil nutrition across elevation of a Sri Lankan tropical rain forest. For. Ecol. Manag. 2008, 256, 1339–1349. [Google Scholar] [CrossRef]
- Alem, S.; Pavlis, J.; Urban, J.; Kucera, J. Pure and Mixed Plantations of Eucalyptus camaldulensis and Cupressus lusitanica: Their Growth Interactions and Effect on Diversity and Density of Undergrowth Woody Plants in Relation to Light. Open J. For. 2015, 5, 375–386. [Google Scholar] [CrossRef]
- Huang, R.; Jia, X.; Liu, T.; Wu, Z.; Xu, M.; Su, Z.-y. Canopy Structure and Understory Radiation Dynamics of Subtropical Ecological Public Welfare Forest. J. Northwest For. Univ. 2020, 35, 28–36. [Google Scholar]
- Kathke, S.; Bruelheide, H. Gap dynamics in a near-natural spruce forest at Mt. Brocken, Germany. For. Ecol. Manag. 2010, 259, 624–632. [Google Scholar] [CrossRef]
- Hu, Z. Study On Spatial Structure Optimization Of Chinese Fir-Phoebe bournei Mixed Forest On Multifunctional Management. Master’s Thesis, Central South University of Forestry and Technology, Changsha, China, 2019. [Google Scholar]
- Zhang, L.; Zhou, P.; Qi, S.; Zhang, D.; Wu, B.; Cui, R. Difference influence of spatial structure of Platycladus orientalis plantations on diversity of understory herbaceous and its correlation degree. Ecol. Environ. Sci. 2022, 31, 1794–1801. [Google Scholar] [CrossRef]
- Li, X.; Zhang, G.; Li, J. Spatial structure evaluation of natural secondary forest around Dongting Lake based on entropy weight-cloud model. J. Coast. Res. 2020, 103, 484–489. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot No. | ASL (m) | Slope (°) | Aspect | Soil Depth (cm) | Tree Density (Stem·ha−1) | Tree Height (m) | Age Stage | DBH (cm) | Plot No. | ASL (m) | Slope (°) | Aspect | Soil Depth (cm) | Tree Density (Stem·ha−1) | Tree Height (m) | Age Stage | DBH (cm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 764 | 35 | semi–SS | 49 | 725 | 6.51 | N | 13.33 | 20 | 714 | 20 | SS | 45 | 925 | 5.3 | Mi | 10.05 |
| 2 | 703 | 28 | semi–SS | 47 | 725 | 5.54 | Mi | 10.89 | 21 | 769 | 28 | SS | 40 | 850 | 14.72 | N | 18.76 |
| 3 | 815 | 25 | semi–SS | 39 | 850 | 9.07 | N | 15.73 | 22 | 930 | 28 | SS | 38 | 875 | 11.81 | Ma | 20.04 |
| 4 | 950 | 27 | semi–SS | 49 | 925 | 3.75 | Y | 6.57 | 23 | 905 | 30 | SS | 43 | 725 | 12.41 | Ma | 21.85 |
| 5 | 833.8 | 12 | semi–SS | 41 | 750 | 7.72 | Mi | 11.44 | 24 | 940 | 33 | SS | 44 | 825 | 4.1 | Mi | 11.9 |
| 6 | 902 | 22 | semi–SS | 42 | 900 | 6.57 | Mi | 15.27 | 25 | 870 | 12 | SS | 43 | 1300 | 7.51 | N | 12.58 |
| 7 | 1023 | 24 | semi–SS | 40 | 1275 | 5.4 | N | 12.33 | 26 | 1271 | 29 | SS | 42 | 1275 | 8.13 | Mi | 14.26 |
| 8 | 992 | 32 | semi–SS | 40 | 725 | 6.94 | N | 15.288 | 27 | 1293 | 26 | SS | 44 | 850 | 6.1 | Mi | 14.05 |
| 9 | 1024 | 32 | semi–SS | 45 | 1300 | 7.41 | Mi | 10.97 | 28 | 1308 | 29 | SS | 42 | 1050 | 7.28 | N | 17.06 |
| 10 | 892 | 20 | semi–SS | 41 | 725 | 7.91 | N | 15.81 | 29 | 1274 | 26 | SS | 42 | 750 | 8.03 | Ma | 24.4 |
| 11 | 847 | 25 | semi–SS | 40 | 775 | 7.72 | Mi | 13.49 | 30 | 1165 | 20 | SS | 42 | 1200 | 6.4 | N | 12.26 |
| 12 | 1123 | 26 | semi–SS | 41 | 1325 | 2.01 | Y | 4.02 | 31 | 1358 | 33 | SS | 42 | 775 | 5.81 | N | 16.74 |
| 13 | 840 | 24 | semi–SS | 44 | 725 | 6.78 | Mi | 11.1 | 32 | 1359 | 28 | SS | 42 | 1250 | 7.54 | N | 14.58 |
| 14 | 914.4 | 24 | semi–SS | 41 | 775 | 7 | N | 12.4 | 33 | 869.6 | 17 | SS | 42 | 950 | 4.72 | N | 12.02 |
| 15 | 877.7 | 35 | semi–SS | 43 | 725 | 10.46 | Ma | 21.1 | 34 | 897 | 28 | SS | 42 | 875 | 11.81 | Ma | 21.04 |
| 16 | 973.2 | 15 | semi–SS | 41 | 1175 | 7.94 | Mi | 11.27 | 35 | 798 | 34 | SS | 42 | 1100 | 11.8 | Mi | 13.68 |
| 17 | 841.5 | 25 | semi–SS | 42 | 725 | 2.29 | Y | 5.69 | 36 | 1053 | 24 | SS | 36 | 925 | 5.3 | Mi | 10.05 |
| 18 | 970 | 19 | semi–SS | 40 | 1250 | 7.14 | N | 12.38 | 37 | 855 | 39 | SS | 49 | 1050 | 5.47 | Mi | 9.14 |
| 19 | 790 | 25 | semi–SS | 35 | 725 | 7.14 | Mi | 9.66 | 38 | 1083 | 15 | SS | 41 | 925 | 12.28 | N | 16.69 |
| Stand Spatial Structure | Mean ± Standard Deviation | Maximum | Minimum | Coefficient of Variation (%) |
|---|---|---|---|---|
| Uniform angle | 0.52 ± 0.08 | 0.75 | 0.33 | 15.09 |
| Neighborhood comparison | 0.52 ± 0.1 | 0.75 | 0.25 | 19.03 |
| Mingling degree | 0.51 ± 0.29 | 1 | 0 | 57.36 |
| Opening degree | 0.6 ± 0.33 | 1.35 | 0.21 | 55.15 |
| Forest layer index | 0.29 ± 0.17 | 0.62 | 0 | 59.88 |
| Competition index | 0.68 ± 0.29 | 1.45 | 0.3 | 42.95 |
| Ecological Functions | Index | Mean ± Standard Deviation | Maximum | Minimum | Coefficient of Variation (%) |
|---|---|---|---|---|---|
| Herb species diversity | Gleason | 0.66 ± 0.17 | 1.11 | 0.33 | 25.74 |
| Shannon–Wiener | 1.16 ± 0.25 | 1.7 | 0.66 | 21.81 | |
| Simpson | 0.64 ± 0.1 | 0.86 | 0.41 | 15.16 | |
| Margalef | 0.66 ± 0.17 | 1.92 | 0.29 | 30.16 | |
| Pielou | 0.95 ± 0.29 | 0.99 | 0.58 | 10.76 | |
| Soil nutrient | Soil organic carbon (g/kg) | 41.56 ± 20.36 | 78.96 | 12.84 | 48.99 |
| Total nitrogen (g/kg) | 2.36 ± 1.3 | 5.29 | 0.73 | 55.11 | |
| Total phosphorus (g/kg) | 1.24 ± 0.67 | 9.1 | 0.22 | 50.27 | |
| Total potassium (g/kg) | 20.09 ± 7.15 | 39.51 | 1.73 | 35.59 | |
| Available nitrogen (mg/kg) | 185.71 ± 97.31 | 404.68 | 31.7 | 52.4 | |
| Available phosphorus (mg/kg) | 2.62 ± 2.25 | 11.13 | 0.3 | 85.9 | |
| Available potassium (g/kg) | 146.57 ± 79.52 | 356.56 | 8.5 | 54.26 | |
| PH | 7.19 ± 0.75 | 8.76 | 5.73 | 10.47 |
| Impact Factor | Function | Direct Effect | Indirect Effect | Total Effect |
|---|---|---|---|---|
| Mingling degree | Species diversity | 0.480 | 0.203 | 0.683 |
| Soil nutrition | 0.697 | −0.076 | 0.621 | |
| Opening degree | Species diversity | 0.370 | −0.001 | 0.369 |
| Soil nutrition | −0.049 | / | −0.049 | |
| Competition index | Species diversity | −0.114 | −0.122 | −0.236 |
| Soil nutrition | 0.201 | 0.019 | 0.220 | |
| Forest layer Index | Species diversity | 0.178 | −0.004 | 0.174 |
| Soil nutrition | −0.128 | / | −0.128 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Qi, S.; Zhang, L.; Guo, Y.; Zhang, D.; Liu, S.; Ma, L.; Hu, J.; Lu, J.; Wang, X. Optimizing Pinus tabuliformis Forest Spatial Structure and Function in Beijing, China. Forests 2024, 15, 1963. https://doi.org/10.3390/f15111963
Zhang Y, Qi S, Zhang L, Guo Y, Zhang D, Liu S, Ma L, Hu J, Lu J, Wang X. Optimizing Pinus tabuliformis Forest Spatial Structure and Function in Beijing, China. Forests. 2024; 15(11):1963. https://doi.org/10.3390/f15111963
Chicago/Turabian StyleZhang, Yan, Shi Qi, Lin Zhang, Yanrui Guo, Dai Zhang, Shaodong Liu, Luxiao Ma, Jun Hu, Jinsheng Lu, and Xiangyu Wang. 2024. "Optimizing Pinus tabuliformis Forest Spatial Structure and Function in Beijing, China" Forests 15, no. 11: 1963. https://doi.org/10.3390/f15111963
APA StyleZhang, Y., Qi, S., Zhang, L., Guo, Y., Zhang, D., Liu, S., Ma, L., Hu, J., Lu, J., & Wang, X. (2024). Optimizing Pinus tabuliformis Forest Spatial Structure and Function in Beijing, China. Forests, 15(11), 1963. https://doi.org/10.3390/f15111963