
Naturalization of the Ornamental Plant Crocus tommasinianus Herb. (Iridaceae) in Forest Ecosystems: A Case Study from Poland


Abstract
1. Introduction
1.1. Problem of Alien and Invasive Plant Species in Europe and in Poland
1.2. Horticulture and Bulbous Plants in the Context of the Spread of Alien Species
1.3. Crocus tommasinianus—A Cultivated, Alien, Naturalized Species
2. Materials and Methods
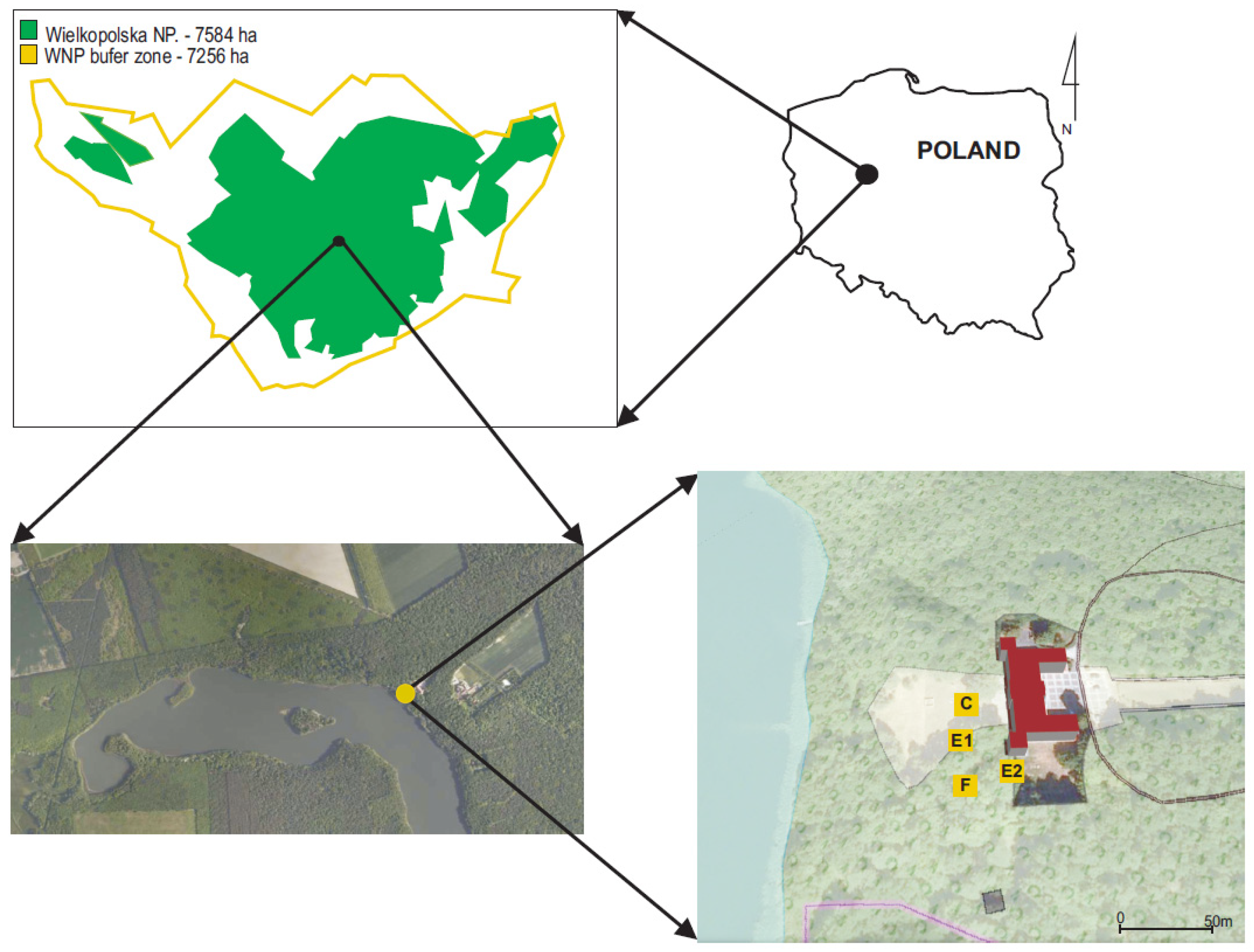
2.1. Description of the New Location
2.2. Morphometric Measurements of Individuals
2.3. Carpological Examination
2.4. Statistical Analyses
3. Results
3.1. Description of the New Location
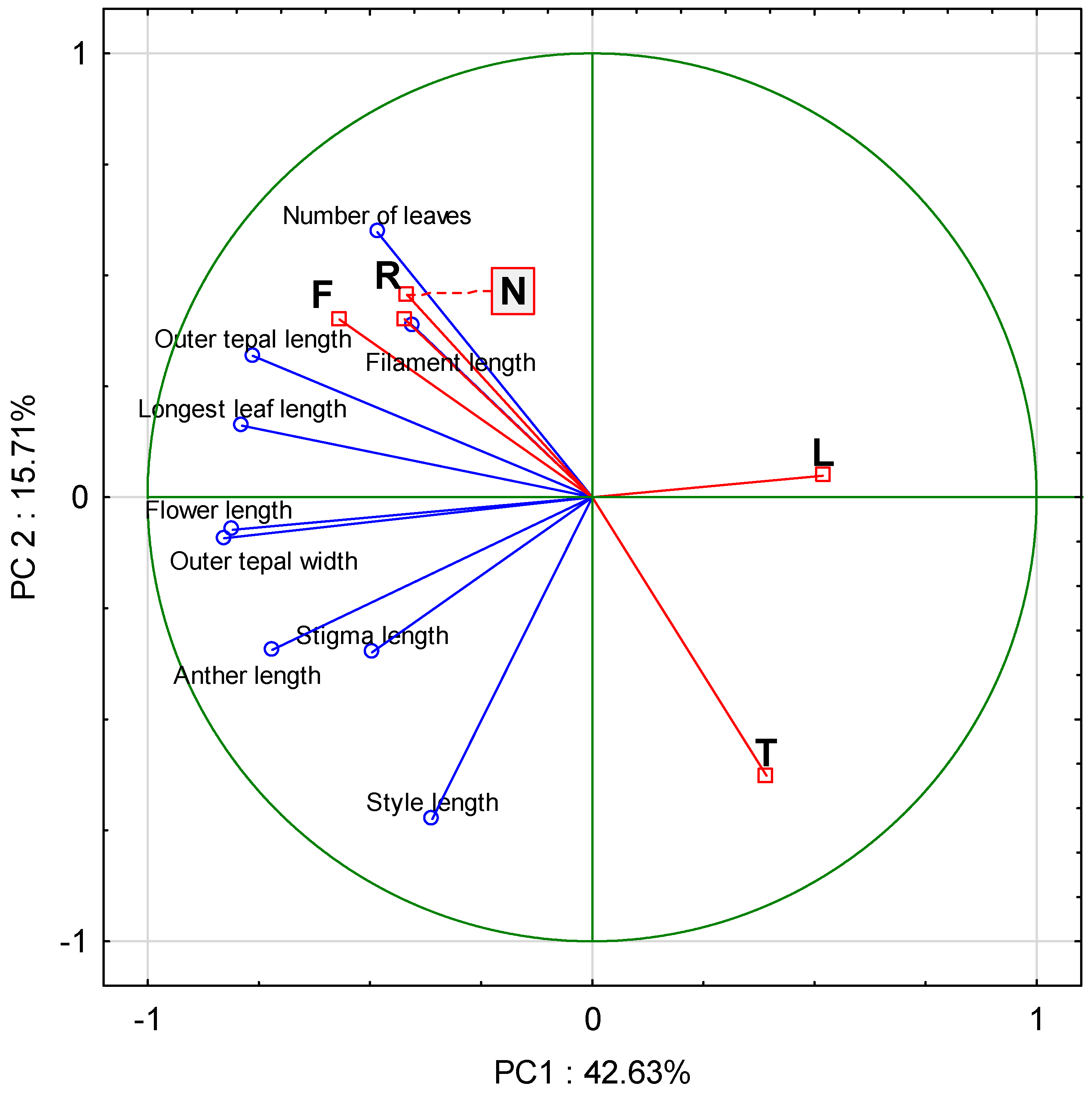
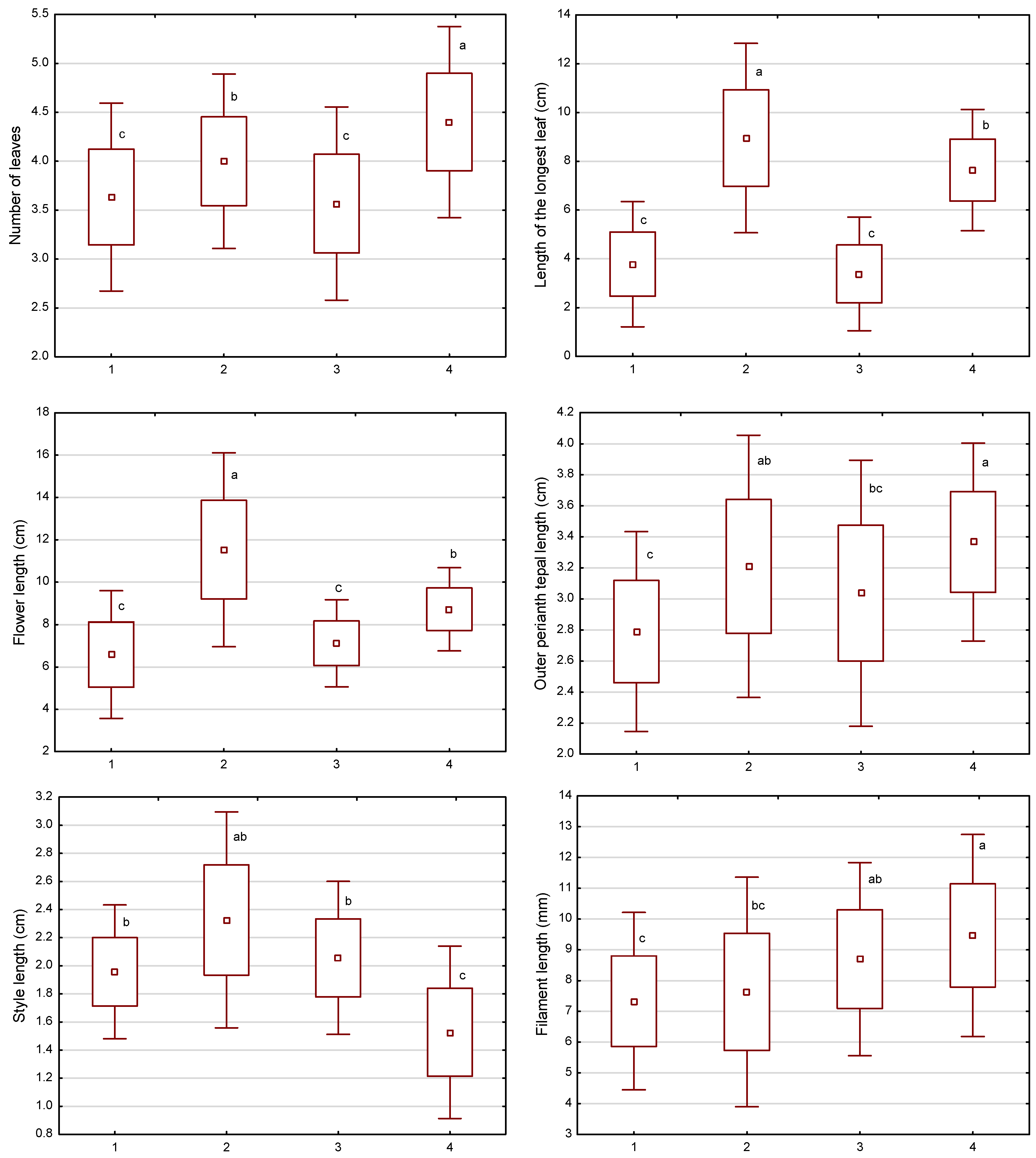
3.2. Morphometric Measurements of Individuals
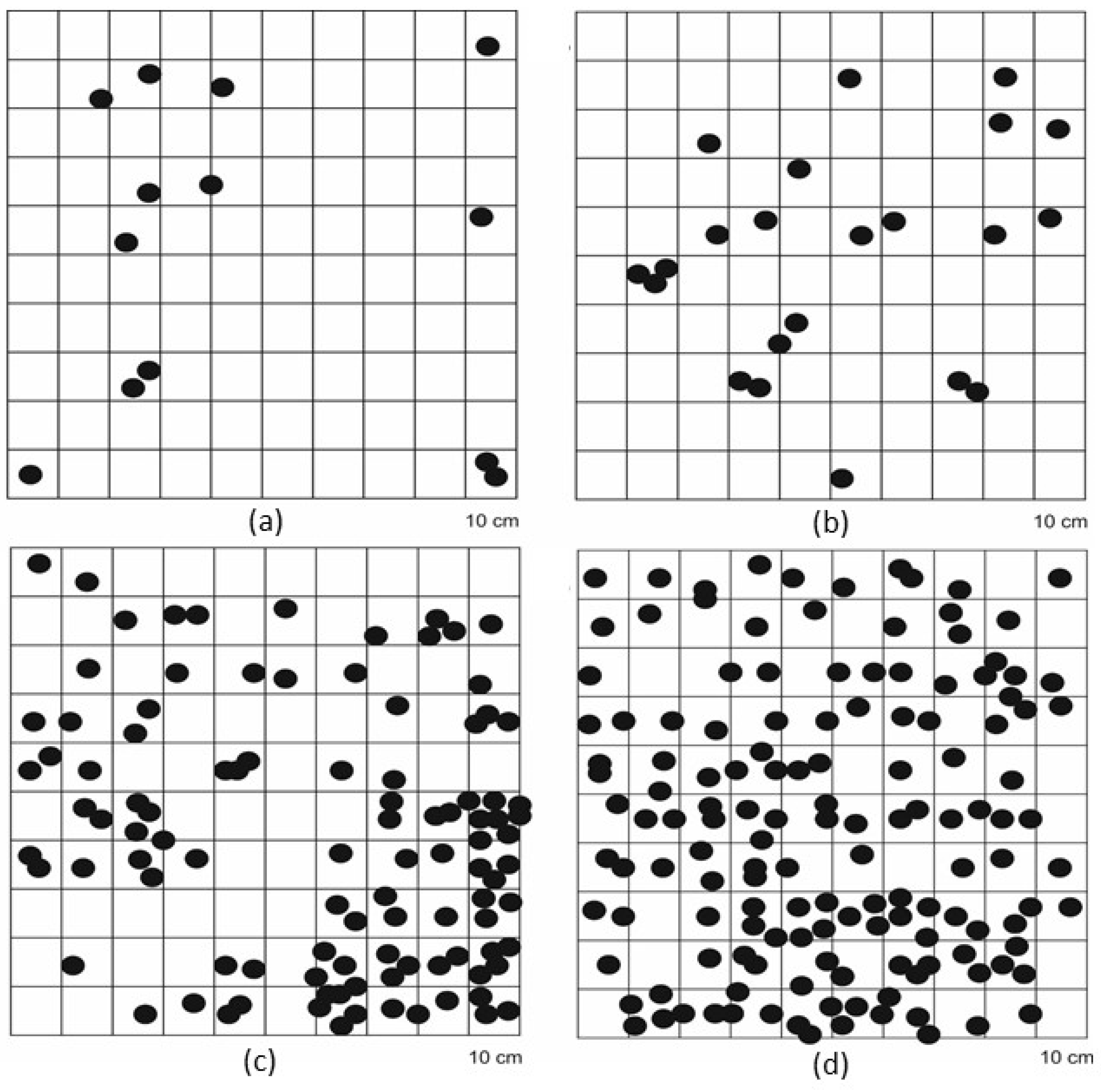
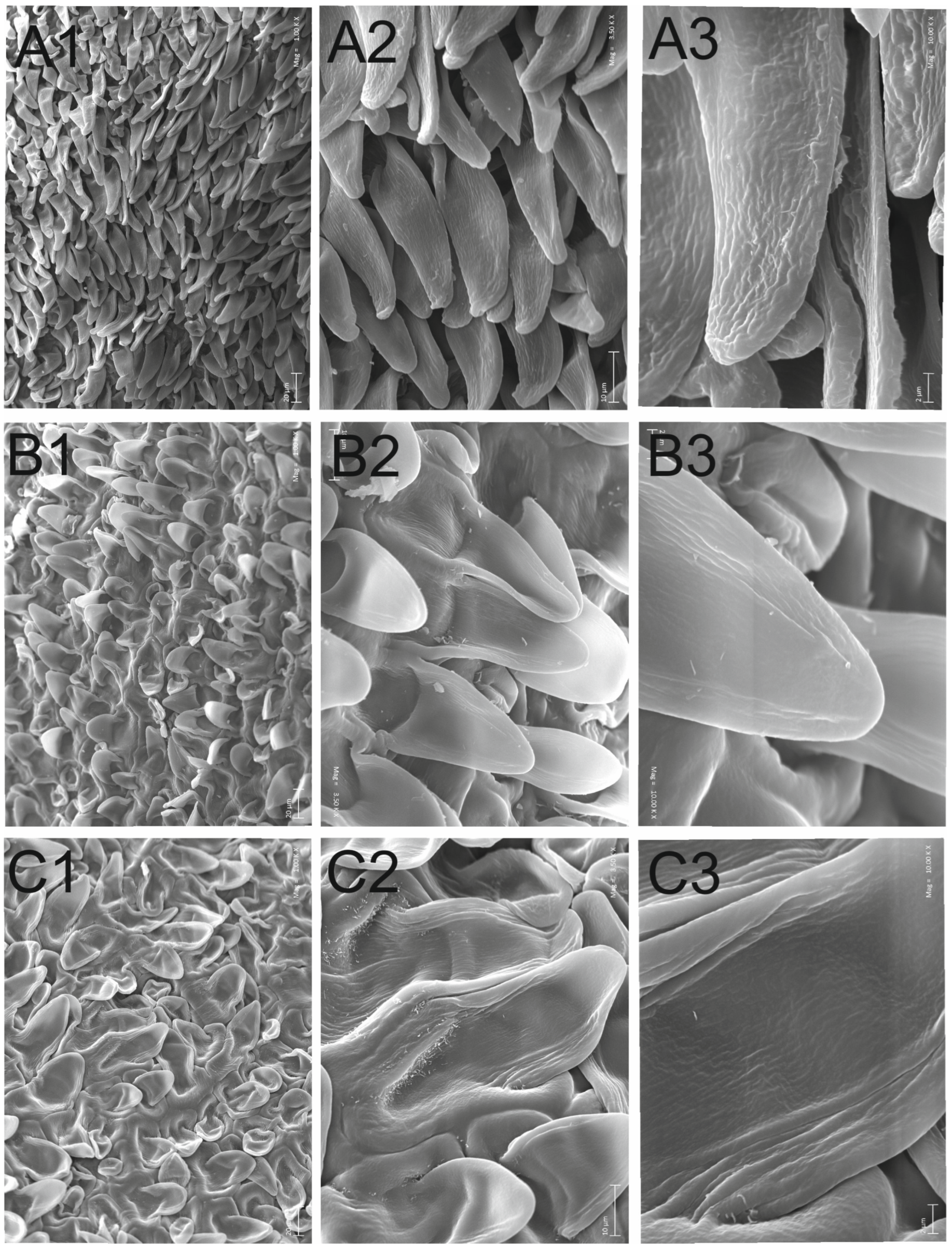
3.3. Carpological Examinaton
4. Discussion
4.1. Description of the New Location
4.2. Morphometric Measurements of Individuals
4.3. Carpological Examination
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position of Locality | 1 | 2 | 3 | 4 | Ellenberg Indicators | ||||
|---|---|---|---|---|---|---|---|---|---|
| L | T | F | R | N | |||||
| Date | 26.03.2021 | 26.03.2021 | 26.03.2021 | 26.03.2021 | |||||
| Coordinates | 52°16′11.0″ N 16°47′81.0″ E | 52°16′08.4″ N 16°47′81.2″ E | 52°16′11.6″ N 16°47′82.2″ E | 52°16′09.5″ N 16°47′84.5″ E | |||||
| Sample collection area [m2] | 10 | 10 | 10 | 8 | |||||
| Cover of tree layer a [%] | 0 | 35 | 40 | 15 | |||||
| Cover of shrub layer b [%] | 0 | 5 | 2 | 0 | |||||
| Cover of herbaceous layer c [%] | 65 | 60 | 50 | 65 | |||||
| Number of species | 22 | 21 | 38 | 11 | |||||
| Crocus tommasinianus Herb. | 1.2 | 1.1 | 2.2 | 3.4 | |||||
| ChO. Arrhenatheretalia Pawł. 1928 | |||||||||
| Achillea millefolium L. | r | . | . | . | 8 | x | 4 | x | 5 |
| Dactylis glomerata L. | . | . | . | r | 7 | x | 5 | x | 6 |
| Taraxacum officinale Web. | + | . | r | 8 | x | 5 | x | 7 | |
| ChCl. Molinio-Arrhenatheretea T.Tx. 1937 | |||||||||
| Festuca pratensis Huds. | 2.2 | . | . | . | 8 | x | 6 | x | 6 |
| Plantago lanceolata L. | + | . | . | . | 6 | x | x | x | x |
| Rumex acetosa L. | r | . | . | . | 8 | x | x | x | 5 |
| ChCl. Querco-Fagetea Br.-Bl. et Vlieg. 1937 | |||||||||
| Acer platanoides L. (a) | . | 1.1 | 1.1 | 1.1 | (5) | 6 | x | x | x |
| Acer platanoides L. (b) | r | + | + | . | |||||
| Aegopodium podagraria L. | . | 1.1 | . | + | 5 | x | 6 | 7 | 8 |
| Anemone nemorosa L. | . | . | + | . | x | x | x | 5 | x |
| Fraxinus excelsior L. (a) | . | . | 1.1 | . | (4) | 5 | x | 7 | 7 |
| Fraxinus excelsior L. (c) | r | . | . | ||||||
| Lonicera xylosteum L. | . | + | . | . | 5 | 5 | 5 | 7 | x |
| Poa nemoralis L. | . | 2.2 | 1.1 | . | 5 | x | 5 | 5 | 3 |
| ChCl. Nardo-Callunetea Prsg 1949 | |||||||||
| Luzula campestris (L.) DC. | . | r | . | . | 7 | x | 4 | 3 | 2 |
| Pilosella officinarum Vaill. | 1.1 | . | . | . | 7 | x | 4 | x | 2 |
| ChCl. Koelerio-Corynephoretea Klika in Klika et Novak, 1941 | |||||||||
| Helichrysum arenarium (L.) Moench | + | . | . | . | 7 | 6 | 3 | 5 | 1 |
| Sedum sexangulare L. | + | . | . | . | 7 | 5 | 7 | 8 | 1 |
| ChAll. Vicio lathyroides-Potentillion Brzeg in Brzeg et M. Woj. 1996 | |||||||||
| Hypochoeris radicata L. | r | . | . | . | 8 | 5 | 5 | 4 | 3 |
| Potentilla argentea L. | r | . | . | . | 9 | x | 2 | 3 | 1 |
| Vicia lathyroides L. | + | . | . | . | 8 | 7 | 2 | 3 | 2 |
| ChAll. Carpinion Issl. 1931 em. Oeberd. 1953 | |||||||||
| Carpinus betulus L. (a) | . | 1.1 | . | 1.1 | (4) | 6 | x | x | x |
| ChAll. Fagion R.Tx. et Diem. 1936 | |||||||||
| Fagus sylvatica L. (c) | . | . | r | . | (3) | 5 | 5 | x | x |
| ChO. Fagetalia Pawł. in Pawł., Sokoł. et Wall. 1928 | . | ||||||||
| Corydalis intermedia (L.) Mérat | . | r | + | . | 3 | 5 | 5 | 7 | 7 |
| Ficaria verna Huds. | 1.1 | + | 1.1 | 4 | 5 | 6 | 7 | 7 | |
| ChAss. Lolio-Cynosuretum R.Tx. 1947 | |||||||||
| Trifiolium repens L. | 1.1 | . | . | . | 8 | x | x | x | 7 |
| ChAll. Alliarion Oberd. (1957) 1962 | . | ||||||||
| Geranium robertianum L. | . | + | + | . | 4 | x | x | x | 7 |
| ChAss. Alliario-Chaerophylletum temuli (Kreh 1935) Lohm. 1949 | |||||||||
| Chaerophyllum temulum L. | . | . | + | . | 5 | 6 | 5 | x | 8 |
| Hedera helix L. | . | + | + | . | (4) | 5 | 5 | x | x |
| ChO. Glechometalia R.Tx. in R.Tx. et Brun.-Hool 1975 | . | ||||||||
| Geum urbanum L. | . | + | . | . | 4 | 5 | 5 | x | 7 |
| ChCl. Festuco-Brometea Br.Bl. et R.Tx. 1943 | . | ||||||||
| Ajuga genevensis L. | . | r | + | . | 8 | x | 4 | 7 | 2 |
| Euphorbia cyparissias L. | . | . | r | . | 8 | x | 3 | x | 3 |
| ChAll. Alliarion Oberd. (1957) 1962 | |||||||||
| Impatiens parviflora DC. | . | + | . | 4 | 6 | 5 | x | 6 | |
| ChO. Glechometalia R.Tx. in R.Tx. et Braun-Hool 1975 | |||||||||
| Alliaria petiolata (M. Bieb.) Cavara and Grande | . | . | + | . | 5 | 6 | 5 | 7 | 9 |
| Glechoma hedaracea L. | + | + | . | 6 | 5 | 6 | x | 7 | |
| ChAss. Hordeo-Brometeum (Allorge 1922) Lohm. 1950 | . | ||||||||
| Bromus sterilis L. | . | + | . | 7 | 7 | 4 | x | 5 | |
| ChAll. Ulmenion minoris Oberd. 1953 | |||||||||
| Ulmus laevis Pall. (a) | . | . | 1.1 | (4) | 6 | 8 | 7 | 7 | |
| ChCl. Stellarietea mediae R.Tx., Lohm. et Prsg. 1950 | . | ||||||||
| Stellaria media (L.) Vill. | . | r | + | 6 | x | 4 | 7 | 8 | |
| Others | |||||||||
| Aesculus hippocastanum L. (a) | . | . | 1.1 | . | . | . | . | . | . |
| Allium vineale L. | r | . | + | . | 5 | 7 | 4 | x | 7 |
| Arenaria serpyllifolia L. | . | . | + | . | 9 | 5 | 4 | x | x |
| Carex spicata Huds. | . | . | r | . | 7 | 5 | 5 | x | 6 |
| Chionodoxa sardensis Barr. et Sudg. | r | . | r | . | |||||
| Conyza canadensis (L.) Cronquist | . | . | + | . | 8 | x | 4 | x | 4 |
| Euonymus alatus (Thunb.) Siebold (b) | . | + | . | . | . | . | . | . | . |
| Festuca trachyphylla Sibth. | 2.2 | . | . | . | . | . | . | . | . |
| Gagea pratensis (Pers.) Dumort. | . | . | + | . | . | . | . | . | . |
| Galanthus nivalis L. | r | . | + | . | 5 | 7 | x | 7 | 7 |
| Geranium molle L. ‘Alba’ | + | . | . | . | 7 | 6 | 3 | 5 | 4 |
| Medicago lupulina L. | r | . | . | . | 7 | 5 | 4 | 8 | x |
| Myosotis stricta Link ex Roem. and Schult. | . | . | + | . | 8 | x | 3 | 4 | 2 |
| Ornithogalum nutans L. | . | . | + | . | . | . | . | . | . |
| Poa annua L. | . | . | . | r | 7 | x | 6 | x | 8 |
| Prunus domestica L. subsp. syriaca (Borkh.) Janch. (b) | . | . | + | . | . | . | . | . | . |
| Pyrus pyraster (L.) Burgsd. (c) | . | r | . | . | . | . | . | . | . |
| Quercus petraea (Matt.) Liebl. (a) | r | 2.2 | 2.2 | . | (6) | 6 | 5 | x | x |
| Scilla sibirica Haw. | . | . | r | 1.1 | . | . | . | . | . |
| Scilla sibirica Haw. ‘Alba’ | . | . | + | + | . | . | . | . | . |
| Veronica chamaedrys L. | + | . | r | . | 6 | x | 4 | x | x |
| Veronica sublobata M. A. Fisch. | . | 1.1 | 1.1 | 6 | 6 | 5 | 7 | 7 | |
| Vinca minor L. | . | 1.1 | + | . | 4 | 6 | 5 | x | 6 |
| Viola suavis M. Bieb. | . | . | r | . | . | . | . | . | . |
References
- Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. Available online: https://eur-lex.europa.eu/eli/reg/2014/1143/oj (accessed on 25 August 2024).
- Pyšek, P.; Richardson, D.M. The biogeography of naturalization in alien plants. J. Biogeogr. 2006, 33, 2040–2050. [Google Scholar] [CrossRef]
- Aguin-Pombo, D. Biological Invasions and Global Trade: An Urgent Issue. In Natural Resources, Sustainability and Humanity: A Comprehensive View; Aguin-Pombo, D., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 83–99. [Google Scholar]
- Van Kleunen, M.; Dawson, W.; Essl, F.; Pergl, J.; Winter, M.; Weber, E.; Pyšek, P. Global exchange and accumulation of non-native plants. Nature 2015, 525, 100–103. [Google Scholar] [CrossRef] [PubMed]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibáñez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [PubMed]
- Zenni, R.D.; Essl, F.; García-Berthou, E.; McDermott, S.M. The economic costs of biological invasions around the world. NeoBiota 2021, 67, 1–9. [Google Scholar] [CrossRef]
- Seebens, H.; Essl, F.; Hulme, P.E.; van Kleunen, M. Development of pathways of global plant invasions in space and time. In Global Plant Invasions; Clements, D.R., Upadhyaya, M.K., Joshi, S., Shrestha, A., Eds.; Springer: Cham, Switzerland, 2022; pp. 35–50. [Google Scholar] [CrossRef]
- Lenzner, B.; Latombe, G.; Schertler, A.; Seebens, H.; Yang, Q.; Winter, M.; Weigelt, P.; van Kleunen, M.; Pyšek, P.; Pergl, J.; et al. Naturalized alien floras still carry the legacy of European colonialism. Nat. Ecol. Evol. 2022, 6, 1723–1732. [Google Scholar] [CrossRef]
- Sudnik-Wójcikowska, B. Rośliny Synantropijne; Multico, Oficyna Wydawnicza: Warsaw, Poland, 2011. [Google Scholar]
- Tokarska-Guzik, B. The Establishment and Spread of Alien Plant Species (Kenophytes) in the Flora of Poland; Wydawnictwo Uniwersytetu Śląskiego: Katowice, Poland, 2005. [Google Scholar]
- Jackowiak, B. Man-made changes in the flora and vegetation of Poland: Current review. Diversity 2023, 15, 618. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce; Generalna Dyrekcja Ochrony Środowiska: Warsaw, Poland, 2012. [Google Scholar]
- Obidziński, A.; Kołaczkowska, E.; Otręba, A. Metody Zwalczania Obcych Gatunków Roślin Występujących na Terenie Puszczy Kampinoskiej; Kampinoski Park Narodowy: Izabelin, Poland; Wydawnictwo BioDar: Kraków, Poland, 2016. [Google Scholar]
- Słowiński, K.; Grygierzec, B.; Synowiec, A.; Tabor, S.; Araniti, F. Preliminary study of control and biochemical characteristics of giant hogweed (Heracleum sosnowskyi Manden.) treated with microwaves. Agronomy 2022, 12, 1335. [Google Scholar] [CrossRef]
- Švec, P.; Perglová, I.; Fröhlich, V.; Laštovička, J.; Seidl, J.; Růžičková, K.; Horáková, I.; Lukavský, J.; Ferko, M.; Štych, P.; et al. Perseverance of management is needed–Efficient long-term strategy of Reynoutria management. NeoBiota 2024, 94, 261–288. [Google Scholar] [CrossRef]
- Szymura, M.; Szymura, T.H.; Wolski, K. Invasive Solidago species: How large area do they occupy and what would be the cost of their removal? Pol. J. Ecol. 2016, 64, 25–34. [Google Scholar] [CrossRef]
- Winter, M.; Schweiger, O.; Klotz, S.; Nentwig, W.; Andriopoulos, P.; Arianoutsou, M.; Basnou, C.; Delipetrou, P.; Didžiulis, V.; Hejda, M.; et al. Plant extinctions and introductions lead to phylogenetic and taxonomic homogenization of the European flora. Proc. Natl. Acad. Sci. USA 2009, 106, 21721–21725. [Google Scholar] [CrossRef]
- Yang, Q.; Weigelt, P.; Fristoe, T.S.; Zhang, Z.; Kreft, H.; Stein, A.; Seebens, H.; Dawson, W.; Essl, F.; König, C.; et al. The global loss of floristic uniqueness. Nat. Commun. 2021, 12, 7290. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Guo, Q. Linking biotic homogenization to habitat type, invasiveness, and growth form of naturalized alien plants in North America. Divers. Distrib. 2010, 16, 119–125. [Google Scholar] [CrossRef]
- Daru, B.H.; Davies, T.J.; Willis, C.G.; Meineke, E.K.; Ronk, A.; Zobel, M.; Pärtel, M.; Antonelli, A.; Davis, C.C. Widespread homogenization of plant communities in the Anthropocene. Nat. Commun. 2021, 12, 6983. [Google Scholar] [CrossRef] [PubMed]
- Šibíková, M.; Jarolímek, I.; Hegedüšová, K.; Májeková, J.; Mikulová, K.; Slabejová, D.; Škodová, I.; Zaliberová, M.; Medvecká, J. Effect of planting alien Robinia pseudoacacia trees on homogenization of Central European forest vegetation. Sci. Total Environ. 2019, 687, 1164–1175. [Google Scholar] [CrossRef]
- Heywood, V.H.; Brunel, S. Code of Conduct on Horticulture and Invasive Alien Plants; Council of Europe Publ: Strasbourg, France, 2008. [Google Scholar]
- Fry, C. The Plant Hunters: The Adventures of the World’s Greatest Botanical Explorers; Univ. Chicago Press: Chicago, IL, USA, 2013. [Google Scholar]
- Richardson, M.D.; McCalla, J.; Buxton, T.; Lulli, F. Incorporating early spring bulbs into dormant warm-season turfgrasses. HortTechnology 2015, 25, 228–232. [Google Scholar] [CrossRef]
- van Kleunen, M.; Essl, F.; Pergl, J.; Brundu, G.; Carboni, M.; Dullinger, S.; Early, R.; González-Moreno, P.; Groom, Q.J.; Hulme, P.E.; et al. The changing role of ornamental horticulture in alien plant invasions. Biol. Rev. 2018, 93, 1421–1437. [Google Scholar] [CrossRef]
- Alam, A.; Iqbal, M.; Vats, S. Cultivation of Some Overlooked Bulbous Ornamentals—A Review on its Commercial Viability. Rep. Opin. 2013, 5, 9–34. [Google Scholar]
- Invasive Species Specialist Group ISSG. The Global Invasive Species Database. Version 2015.1. Available online: https://www.iucngisd.org/gisd/ (accessed on 24 September 2024).
- Plants of the World Online (POWO). Available online: https://powo.science.kew.org (accessed on 25 February 2024).
- Hoste, I. Ornamentals and Invasive Plants: If You Choose One You Get the Other as Well. In Mini Symposium of Aliens and Invasive Species; National Botanic Garden of Belgium, Bouchout Castle: Meise, Belgium, 2011. [Google Scholar]
- Czarna, A.; Klimko, M.; Janyszek, S. Vascular Flora and Vegetation of the Former Manor Park in Radojewo (Wielkopolska Region, Poland). Rocz. AR Pozn. 2009, 388, 37–47. [Google Scholar]
- Czarna, A. Vascular Flora in the Park at Jeziory in the National Park of Wielkopolska. Rocz. AR Pozn. 2010, 389, 39–44. [Google Scholar]
- Czarna, A. Vascular Plant Flora in the Cytadela Cemeteries in Poznań (Poland). Acta Agrobot. 2016, 69, 1–17. [Google Scholar] [CrossRef]
- Czarna, A. Vascular Plants in the Cemetery of the Meritorious (Cmentarz Zasłużonych) in Poznań (Poland). Ann. UMCS 2016, 71, 59–73. [Google Scholar] [CrossRef]
- Nowińska, R.; Czarna, A.; Czekalski, M.; Morozowska, M. Vascular Flora of Selected Palace Parks in the Wielkopolska Region. Steciana 2016, 20, 137–157. [Google Scholar] [CrossRef]
- Żukowski, W.; Latowski, K.; Jackowiak, B.; Chmiel, J. Rośliny Naczyniowe Wielkopolskiego Parku Narodowego. In Prace Zakładu Taksonomii Roślin Uniwersytetu im. Adama Mickiewicza w Poznaniu; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 1995. [Google Scholar]
- Drobnik, J. Allium paradoxum from Asia to Europe: Ornamental, Invasive, Edible, and Medicinal. Hum. Ecol. 2023, 51, 559–567. [Google Scholar] [CrossRef]
- Rukšāns, J. The World of Crocuses, the First Supplement; The Latvian Academy of Sciences: Riga, Latvia, 2023. [Google Scholar]
- Mathew, B.F. Crocus L. In Flora Europaea, Vol. 5: Alismataceae to Orchidaceae (Monocotyledones); Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 92–97. [Google Scholar]
- Petersen, G.; Seberg, O.; Thorsøe, S.; Jørgensen, T.; Mathew, B. A phylogeny of the genus Crocus (Iridaceae) based on sequence data from five plastid regions. Taxon 2008, 57, 487–499. [Google Scholar] [CrossRef]
- Surányi, G.; Máthé, C.; Mosolygó, Á.; Borbély, G.; Vasas, G. Analysis of genetic diversity in crocuses with Carpathian Basin origin using AFLP-markers. Acta Biol. Hung. 2010, 61, 149–155. [Google Scholar] [CrossRef]
- Alsayied, N.F.; Fernández, J.A.; Schwarzacher, T.; Heslop-Harrison, J.S. Diversity and relationships of Crocus sativus and its relatives analysed by inter-retroelement amplified polymorphism (IRAP). Ann. Bot. 2015, 116, 359–368. [Google Scholar] [CrossRef]
- Rukšans, J. The World of Crocuses; Latvian Academy of Sciences: Riga, Latvia, 2017. [Google Scholar]
- Rand, E.S. Popular Flowers, and How to Cultivate Them; Hurd and Houghton: New York, NY, USA, 1876. [Google Scholar]
- Snowarski, M. Flora Polski, Atlas Roślin.pl. Available online: https://atlas-roslin.pl/index.html (accessed on 25 February 2024).
- Harris, S. Crocus species (Iridaceae). Oxford University Plants 400. Department of Plant Sciences; Oxford University. Available online: https://herbaria.plants.ox.ac.uk/bol/plants400/Profiles/cd/crocus (accessed on 18 October 2024).
- Szilagyi, C.; Băla, M.; Toța, C. The behaviour of some crocus species upon different fertilizers and substrates. J. Name 2018, 22, 98–101. [Google Scholar]
- Wurzell, B. Spring flowering crocuses. BSBI News 1992, 60, 36–38. [Google Scholar]
- The Complete List of Taxon Names from the BSBI’s Database. Available online: https://bsbi.org/taxon-lists (accessed on 14 October 2024).
- McAwoy, M. Non-Native Plants of Delaware; Species Conservation and Research Program, Delaware Department of Natural Resources and Environmental Control, Division of Fish and Wildlife: Dover, DE, USA, 2024. [Google Scholar]
- Tuttle, J. Plant spring blooming bulbs now. In Ask a Master Gardener; University of Minnesota Extension: St Paul, MN, USA, 2023; pp. 1–6. [Google Scholar]
- Brighton, C.A. Cytological problems in the genus Crocus (Iridaceae): I. Crocus vernus aggregate. Kew Bull. 1976, 31, 33–46. [Google Scholar] [CrossRef]
- Fedorov, A.A. Flora Partis Europaeae URSS. 1979; Volume 4. [Google Scholar]
- Dostál, J. Nová Květena ČSSR; Academia: Prague, Czech Republic, 1989; 1548p. [Google Scholar]
- Prisžter, S.Z. Uj säfränyfaj (Crocus tommasinianus HERB.) Magyarorszägon. Bot. Közlemények 1964, 51, 183–186. [Google Scholar]
- Mihaly, A.; Kricsfalusy, V. Population biology and ecology of Crocus heuffelianus Herb. (Iridaceae) in Ukraine. Linz. Biol. Beiträge 1997, 29, 641–681. [Google Scholar]
- Ževrnja, N.; Vladović, D. The genus Crocus L. in the flora of Svilaja Mountain. Nat. Croat. Period. Musei Hist. Nat. Croat. 2005, 14, 363–368. [Google Scholar]
- Hitchmough, J.; Fieldhouse, K. Plant User Handbook: A Guide to Effective Specifying; Blackwell Science: Oxford, UK, 2004. [Google Scholar] [CrossRef]
- Ogle, C.C.; La Cock, G.D. Additional records and observations of monocotyledons naturalised or casual in Manawatu Ecological Region, New Zealand. Perspect. Biosecurity 2019, 4, 7–31. [Google Scholar]
- Hill, M.O.; Preston, C.D.; Shanklin, J.D. Geographical patterns in the flora of Cambridgeshire (vc 29). Br. Ir. Bot. 2020, 2, 285–308. [Google Scholar] [CrossRef]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Vascular Plants of Poland: An Annotated Checklist; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2020. [Google Scholar]
- Harpke, D.; Carta, A.; Tomović, G.; Ranđelović, V.; Ranđelović, N.; Blattner, F.R.; Peruzzi, L. Phylogeny, karyotype evolution and taxonomy of Crocus series Verni (Iridaceae). Plant Syst. Evol. 2015, 301, 309–325. [Google Scholar] [CrossRef]
- Mosolygó, Á.; Sramkó, G.; Barabás, S.; Czeglédi, L.; Jávor, A.; Molnár, A.V.; Surányi, G. Molecular genetic evidence for allotetraploid hybrid speciation in the genus Crocus L. (Iridaceae). Phytotaxa 2016, 258, 121–136. [Google Scholar] [CrossRef]
- Raca, I.; Blattner, F.R.; Waminal, N.E.; Kerndorff, H.; Ranđelović, V.; Harpke, D. Disentangling Crocus series Verni and its polyploids. Biology 2023, 12, 303. [Google Scholar] [CrossRef]
- Kandemir, N. Comparative leaf anatomy of some endemic Crocus L. taxa from Turkey. Bangladesh J. Bot. 2011, 40, 155–162. [Google Scholar] [CrossRef]
- Raca, I.; Ljubisavljević, I.; Jušković, M.; Ranđelović, N.; Ranđelović, V. Comparative anatomical study of the taxa from series Verni Mathew (Crocus L.) in Serbia. Biol. Nyssana 2017, 8, 15–22. [Google Scholar] [CrossRef]
- Raca, I.; Jovanovic, M.; Ljubisavljevic, I.; Juskovic, M.; Randelovic, V. Morphological and leaf anatomical variability of Crocus cf. heuffelianus Herb. (Iridaceae) populations from the different habitats of the Balkan Peninsula. Turk. J. Bot. 2019, 43, 645–658. [Google Scholar] [CrossRef]
- Kerndorff, H.; Pasche, E.; Harpke, D. The Genus Crocus (Liliiflorae, Iridaceae): Life Cycle, Morphology, Phenotypic Characteristics, and Taxonomically Relevant Parameters. Stapfia 2015, 103, 27–65. [Google Scholar]
- Karaismailoğlu, M.C.; Şik, L.; Gemicioğlu, A.; Erol, O. Seed Structure of Some Taxa of the Genus Crocus L. (Iridaceae) Series Crocus. Turk. J. Bot. 2018, 42, 722–731. [Google Scholar] [CrossRef]
- Song, Y.X.; Peng, S.; Mutie, F.M.; Jiang, H.; Ren, J.; Cong, Y.Y.; Hu, G.W. Evolution and taxonomic significance of seed micromorphology in Impatiens (Balsaminaceae). Front. Plant Sci. 2022, 13, 835943. [Google Scholar] [CrossRef] [PubMed]
- Dyderski, M.K.; Jagodziński, A.M. Impact of invasive tree species on natural regeneration species composition, diversity, and density. Forests 2020, 11, 456. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde, 3rd ed.; Springer: Vienna, Austria, 1964. [Google Scholar]
- Di Biase, L.; Tsafack, N.; Pace, L.; Fattorini, S. Ellenberg Indicator Values Disclose Complex Environmental Filtering Processes in Plant Communities along an Elevational Gradient. Biology 2023, 12, 161. [Google Scholar] [CrossRef]
- Ellenberg, H. Zeigerwerte der Gefäßpflanzen Mitteleuropas; Verlag Erich Goltze KG: Göttingen, Germany, 1974. [Google Scholar]
- Cattell, R.B. The Scree Test for the Number of Factors. Multivar. Behav. Res. 1966, 1, 245–276. [Google Scholar] [CrossRef]
- Chudzicki, M. Operat Ekosystemów Leśnych Wielkopolskiego Parku Narodowego—Opis Taksacyjny cz. 2. Projekt Planu Ochrony Wielkopolskiego Parku Narodowego; Biuro Urządzania Lasu i Geodezji Leśnej Oddział w Poznaniu: Poznań, Poland, 2013. [Google Scholar]
- Szafer, W.; Kulczyński, S.; Pawłowski, B. Rośliny Polskie; Volume I-II; PWN: Warsaw, Poland, 1988. [Google Scholar]
- Kornaś, J. Geograficzno-historyczna klasyfikacja roślin synantropijnych. Mater. Zakładu Fitosocjologii Stosow. UW 1968, 25, 33–41. [Google Scholar]
- iNaturalist. Crocus tommasinianus, C. vernus . iNaturalist. Available online: https://www.inaturalist.org (accessed on 23 September 2024).
- Krause, J. Niskie, Ale Pięknie Kwitnące Rośliny Cebulowe. Krokus, Szafirek; PWRiL: Warszawa, Poland, 1987. [Google Scholar]
- Knebel, C.H. Rośliny Cebulowe; Klub dla Ciebie: Warsaw, Poland, 2006. [Google Scholar]
- Wendebourg, T. Zwiebelpflanzen für den Garten; Verlag Eugen Ulmer: Stuttgart, Germany, 2004. [Google Scholar]
- Adams, K. Krokusy, Tulipany, Lilie; Świat Książki: Warsaw, Poland, 2014. [Google Scholar]
- Johansson, B.G. Crocus på Gotland. Gotlands Bot. Förening 1991, 11, 28–32. [Google Scholar]
- Wall, S.V.; Longland, W.S. Diplochory and the evolution of seed dispersal. In Seed Fate: Predation, Dispersal and Seedling Establishment; Forget, P.M., Lambert, J.E., Hulme, P.E., Vander Wall, S.B., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 297–314. [Google Scholar] [CrossRef]
- Gorb, E.; Gorb, S. Seed Dispersal by Ants in a Deciduous Forest Ecosystem: Mechanisms, Strategies, Adaptations; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Peruzzi, L.; Carta, A. Crocus ilvensis sp. nov. (Sect. Crocus, Iridaceae), Endemic to Elba Island (Tuscan Archipelago, Italy). Nord. J. Bot. 2011, 29, 6–13. [Google Scholar] [CrossRef]
- Biały, K. The Effect of Boar (Sus scrofa) Rooting on the Distribution of Organic Matter in Soil Profiles and the Development of Wood Anemone (Anemone nemorosa L.) in the Oak-Hornbeam Stand (Tilio-Carpinetum) in the Białowieza Primeval Forest. Forestry 1996, 38, 77–88. [Google Scholar] [CrossRef]
- Faliński, J.B. Interaction: Wild Boar Rooting—Participation of Geophytes in the Herb Layer of Oak-Linden-Hornbeam Forest. Phytocoennosis 2002, 5, 70. [Google Scholar]
- Podgórski, T.; Lusseau, D.; Scandura, M.; Sönnichsen, L.; Jędrzejewska, B. Long-Lasting, Kin-Directed Female Interactions in a Spatially Structured Wild Boar Social Network. PLoS ONE 2014, 9, e99875. [Google Scholar] [CrossRef] [PubMed]
- Faliński, J.B. Stadia Neofityzmu i Stosunek Neofitów do Innych Komponentów Zbiorowiska. Mater. Zakładu Fitosocjologii Stosow. Uniw. Warsz. 1968, 25, 15–31. [Google Scholar]
- Faliński, J.B. Inwazje w Świecie Roślin: Mechanizmy, Zagrożenia, Projekt Badań. Phytocoen. N.S. 2004, 16, 1–32. [Google Scholar]
- Piskorz, R.; Klimko, M. Współwystępowanie Niecierpka Drobnokwiatowego Impatiens parviflora DC. i wybranych roślin lasu dębowo-grabowego w Wielkopolskim Parku Narodowym. Sylwan 2007, 151, 43–58. [Google Scholar]
- Rice, E.L. Allelopathy; Acad. Press: New York, NY, USA, 1984. [Google Scholar]
- Wink, M. Evolution of Secondary Metabolites from an Ecological and Molecular Phylogenetic Perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Barkhordari, K.; Sorooshzadeh, A.; Mokhtassi, B.A. Allelopathic Effect of Extraction Solution of Leaves and Corms of Saffron (Crocus sativus) in Phenological Stages on Seed Germination of Jimson Weed (Datura stramonium). Modares J. Biotechnol. 2018, 9, 233–239. [Google Scholar]
- Mikolajchuk, V.; Panfilova, A.; Korkhova, M.; Drobitko, A. Allelopathic Activity of Water-Soluble and Volatile Secretions of Crocus sativus L. (Iridaceae) Flowers in the Northern Black Sea Region. J. Biotechnol. 2022, 9, 233–239. [Google Scholar] [CrossRef]
- Hosseini, M.; Rizvi, S.J.H. A Preliminary Investigation on Possible Role of Allelopathy in Saffron (Crocus sativus L.). In II International Symposium on Saffron Biology and Technology; Kafi, M., Koocheki, A., Eds.; ISHS: Leuven, Belgium, 2006; pp. 75–79. [Google Scholar]
- Feizi, H.; Salari, A.; Gharar, F. Study of the Allelopathic Effect of Saffron (Crocus sativus L.) Organs’ Aqueous Extract on the Seed Germination and Seedling Growth of Sugar Beet and Safflower at Different Concentrations. J. Med. Spice Plants 2018, 22, 156–161. [Google Scholar]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of Allelopathy on Plant Performance: A Meta-Analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Dolatyari, A.; Abolhasani, M.T.; Ardalani, F.; Rukšāns, J. A taxonomic revision of the genus Crocus (Iridaceae) in Iran. Nord. J. Bot. 2024, e04270. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005; Available online: https://www.bsbi.org/ (accessed on 17 October 2014).



| Character | Mean ± SD | Min. | Max. |
|---|---|---|---|
| Number of leaves | 3.90 ± 0.59 | 3.0 | 5.0 |
| Length of longest leaf (cm) | 5.94 ± 2.82 | 2.0 | 14.5 |
| Flower length (cm) | 8.49 ± 2.48 | 4.5 | 16.5 |
| Outer tepal length (cm) | 3.10 ± 0.44 | 2.2 | 4.5 |
| Outer tepal width (cm) | 1.03 ± 0.19 | 0.6 | 1.5 |
| Stigma length (mm) | 3.90 ± 0.80 | 2.0 | 6.0 |
| Style length (cm) | 1.97 ± 0.42 | 0.8 | 3.0 |
| Anther length (cm) | 1.27 ± 0.17 | 0.8 | 1.7 |
| Filament length (mm) | 8.35 ± 1.74 | 4.0 | 14.0 |
| Character | Wilk’s Lambda | Partial Lambda | p | Standardized Coefficients | ANOVA F | p | |
|---|---|---|---|---|---|---|---|
| CAN1 | CAN2 | ||||||
| Number of leaves | 0.05 | 0.92 | 0.03 | −0.09 | −0.32 | 17.48 | <0.001 |
| Length of longest leaf (cm) | 0.09 | 0.59 | <0.001 | −0.84 | −0.29 | 107.62 | <0.001 |
| Flower length (cm) | 0.06 | 0.82 | <0.001 | −0.46 | 0.40 | 62.47 | <0.001 |
| Outer tepal length (cm) | 0.06 | 0.91 | 0.02 | 0.33 | −0.30 | 12.95 | <0.001 |
| Outer tepal width (cm) | 0.05 | 0.97 | 0.32 | −0.15 | 0.01 | 12.10 | <0.001 |
| Stigma length (mm) | 0.06 | 0.84 | <0.001 | 0.34 | −0.35 | 3.10 | 0.03 |
| Style length (cm) | 0.09 | 0.54 | <0.001 | 0.03 | 0.94 | 36.99 | <0.001 |
| Anther length (cm) | 0.05 | 0.96 | 0.26 | −0.04 | 0.16 | 10.00 | <0.001 |
| Filament length (mm) | 0.07 | 0.77 | <0.001 | 0.22 | −0.59 | 7.63 | <0.001 |
| Eigenvalues | 3.59 | 2.51 | |||||
| Cumul. total variance expl. (%) | 0.57 | 0.96 | |||||
| Crocus Species | Seed Length (mm) | Seed Width (mm) | Elaiosome Length (mm) | |
|---|---|---|---|---|
| C. vernus | Mean ± SD | 2.87 ± 0.24 ab | 2.13 ± 0.22 | 0.42 ± 0.06 |
| Min–Max | 2.13–3.15 | 1.71–2.43 | 0.30–0.53 | |
| C. scepusiensis | Mean ± SD | 3.06 ± 0.26 a | 2.10 ± 0.10 | 0.47 ± 0.11 |
| Min–Max | 2.77–3.69 | 1.90–2.28 | 0.30–0.65 | |
| C. tommasinianus | Mean ± SD | 2.78 ± 020 b | 2.08 ± 0.16 | 0.48 ± 0.09 |
| Min–Max | 2.43–3.00 | 1.63–2.24 | 0.34–0.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowińska, R.; Czarna, A. Naturalization of the Ornamental Plant Crocus tommasinianus Herb. (Iridaceae) in Forest Ecosystems: A Case Study from Poland. Forests 2024, 15, 1851. https://doi.org/10.3390/f15111851
Nowińska R, Czarna A. Naturalization of the Ornamental Plant Crocus tommasinianus Herb. (Iridaceae) in Forest Ecosystems: A Case Study from Poland. Forests. 2024; 15(11):1851. https://doi.org/10.3390/f15111851
Chicago/Turabian StyleNowińska, Renata, and Aneta Czarna. 2024. "Naturalization of the Ornamental Plant Crocus tommasinianus Herb. (Iridaceae) in Forest Ecosystems: A Case Study from Poland" Forests 15, no. 11: 1851. https://doi.org/10.3390/f15111851
APA StyleNowińska, R., & Czarna, A. (2024). Naturalization of the Ornamental Plant Crocus tommasinianus Herb. (Iridaceae) in Forest Ecosystems: A Case Study from Poland. Forests, 15(11), 1851. https://doi.org/10.3390/f15111851