
Suitable Habitat Prediction and Analysis of Dendrolimus houi and Its Host Cupressus funebris in the Chinese Region


Abstract
1. Introduction
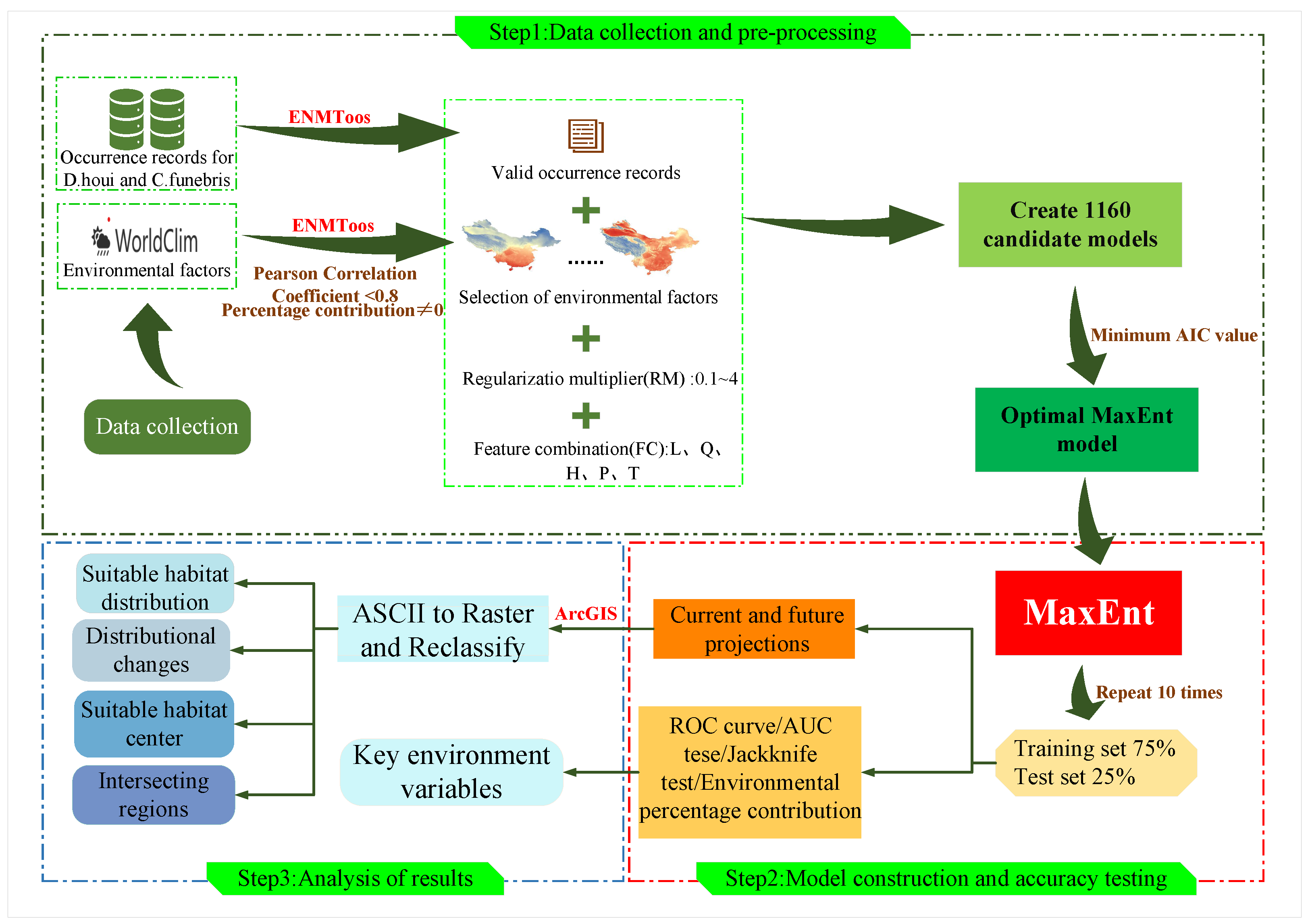
2. Materials and Methods
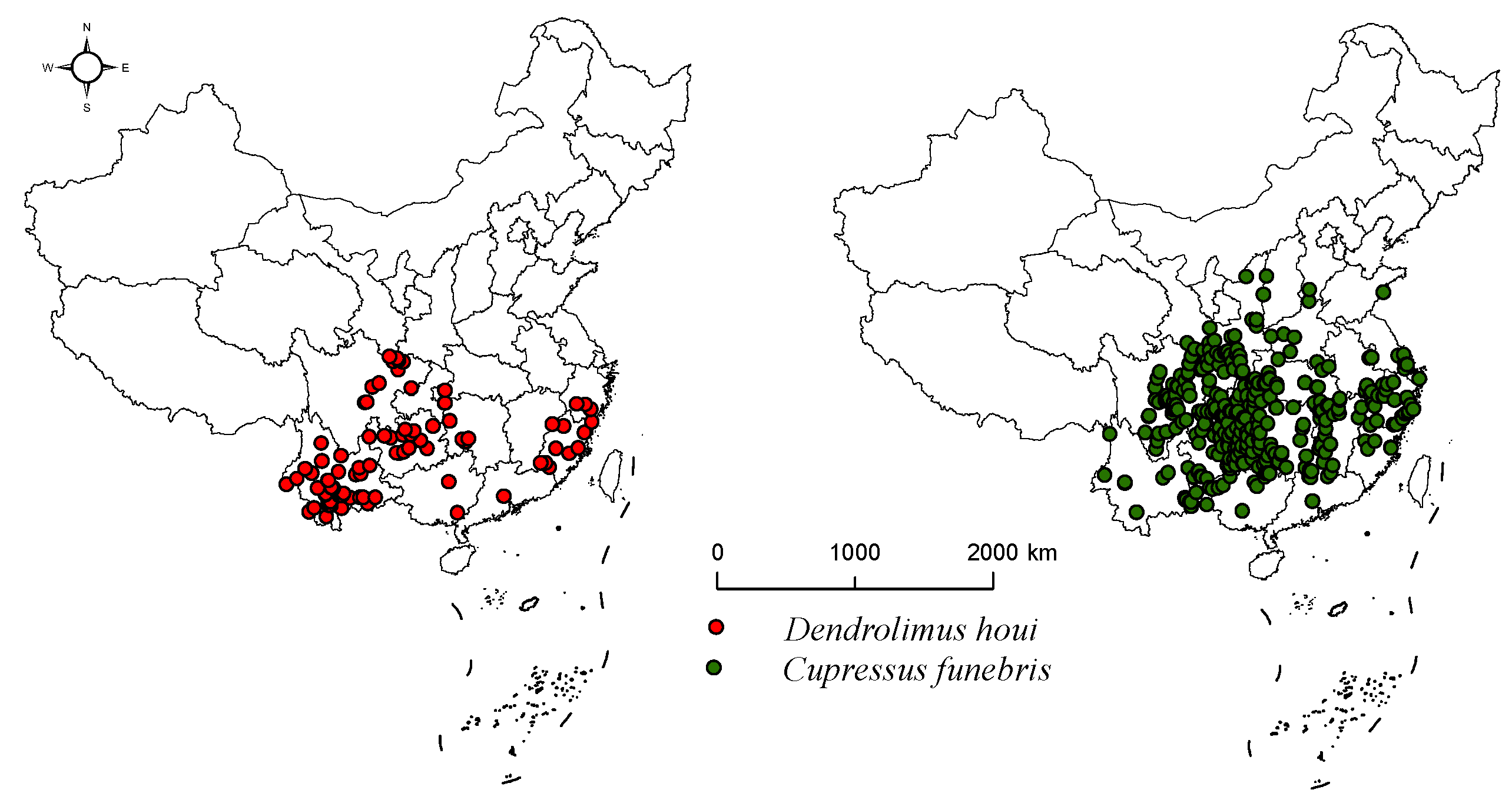
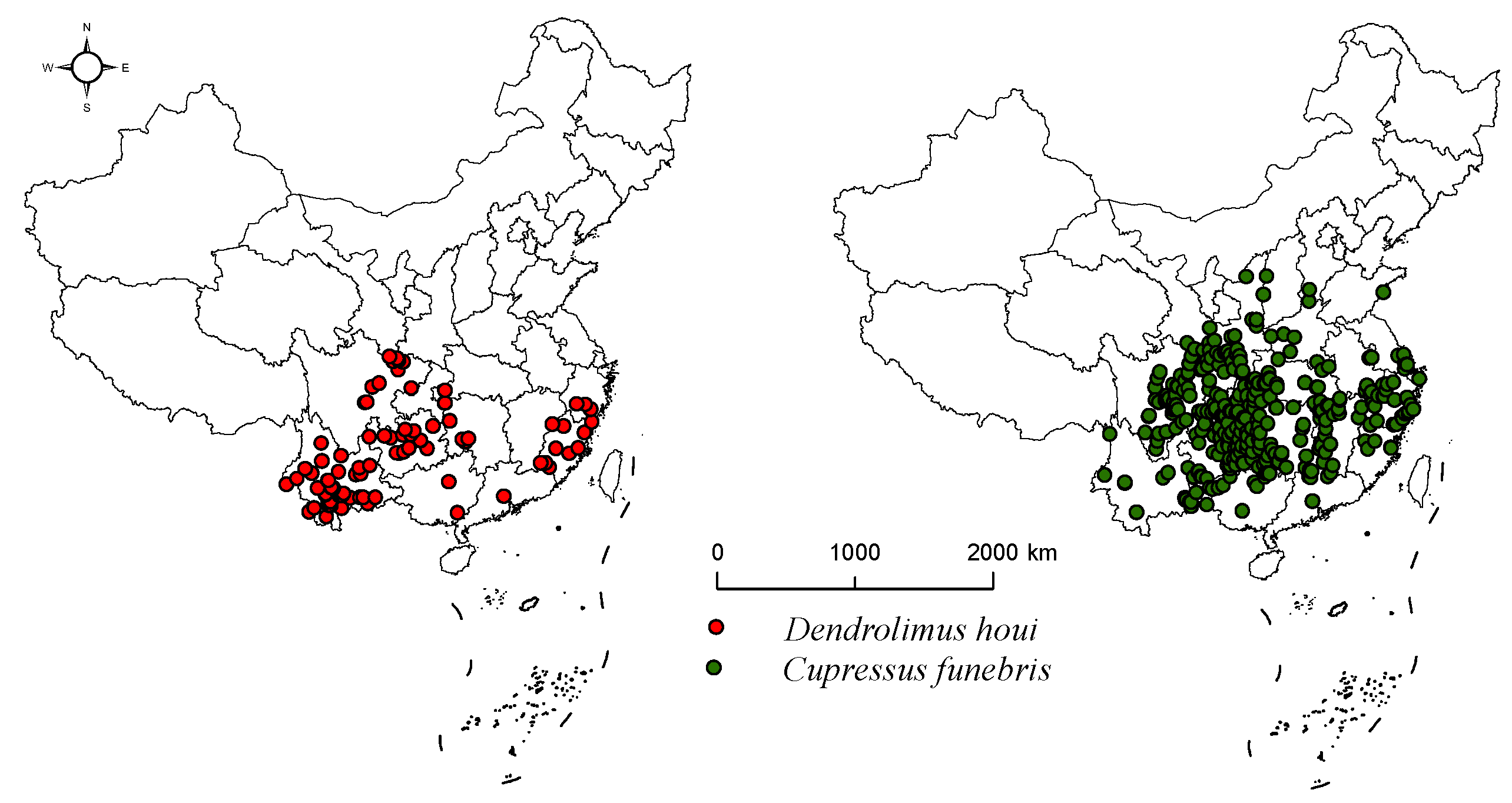
2.1. Data on the Geographical Distribution of Species
2.2. Environmental Data
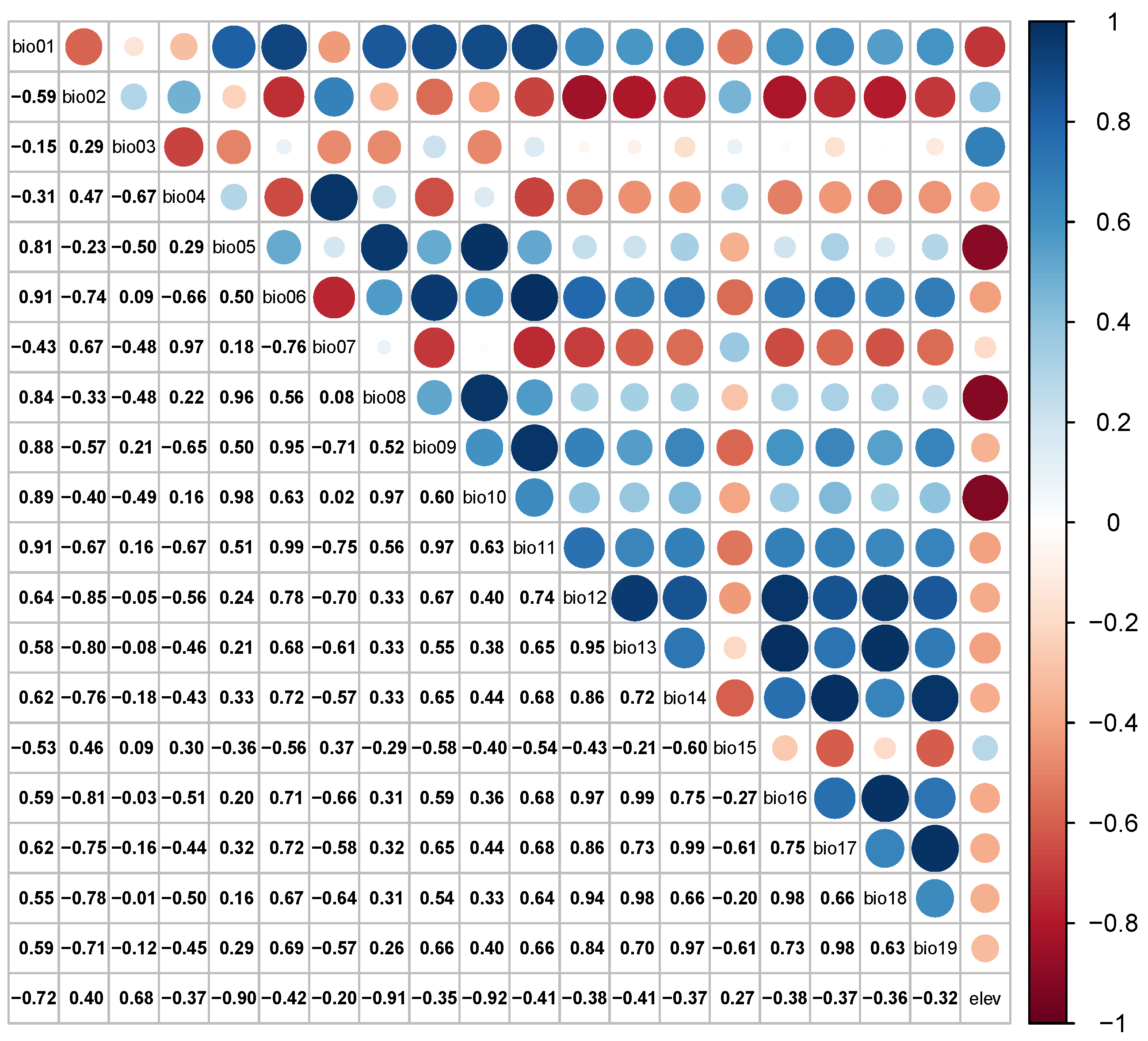
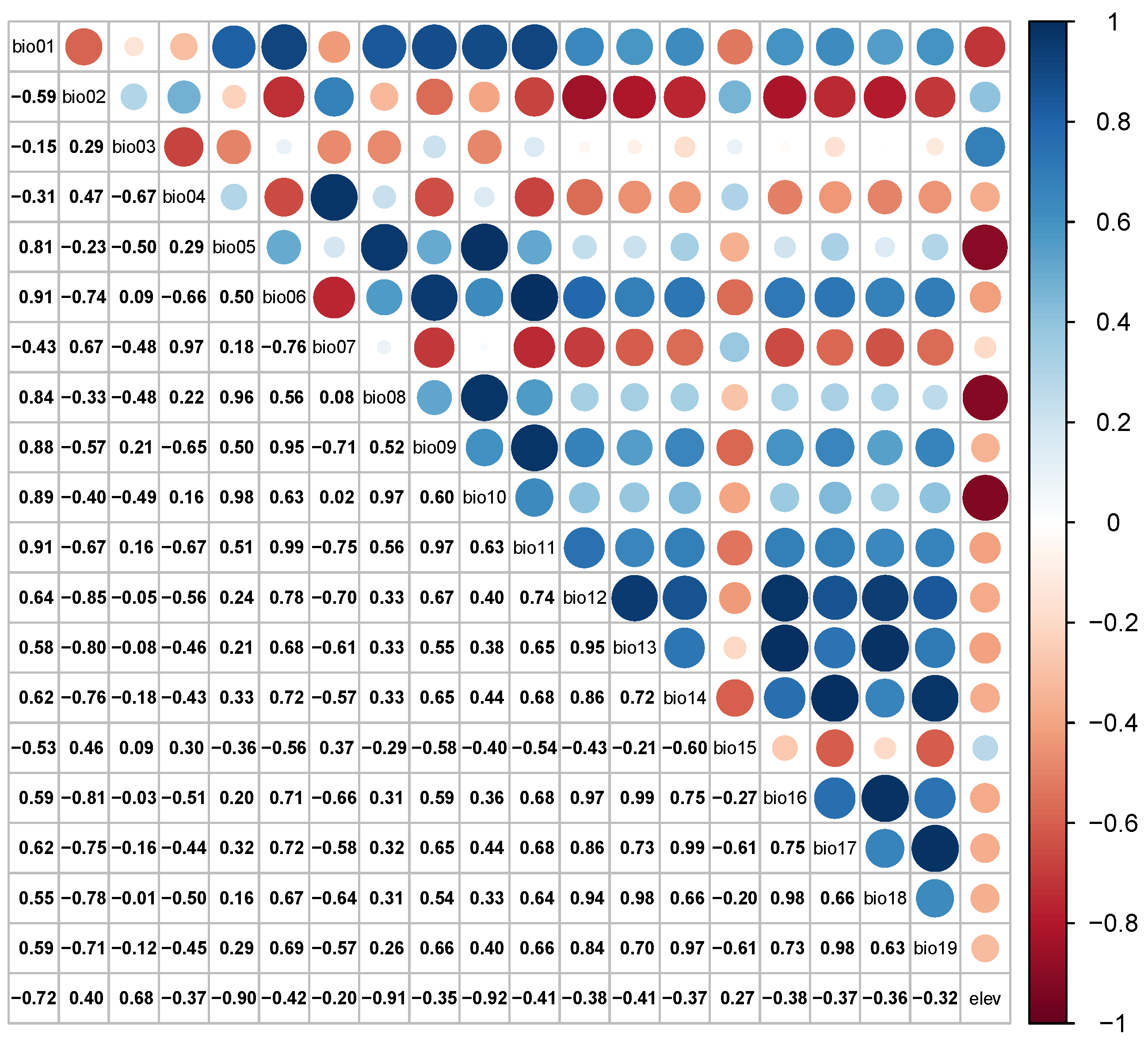
2.3. Environmental Factors Pre-Treatment
2.4. Model Construction and Evaluation
2.5. Analysis of Results
3. Result
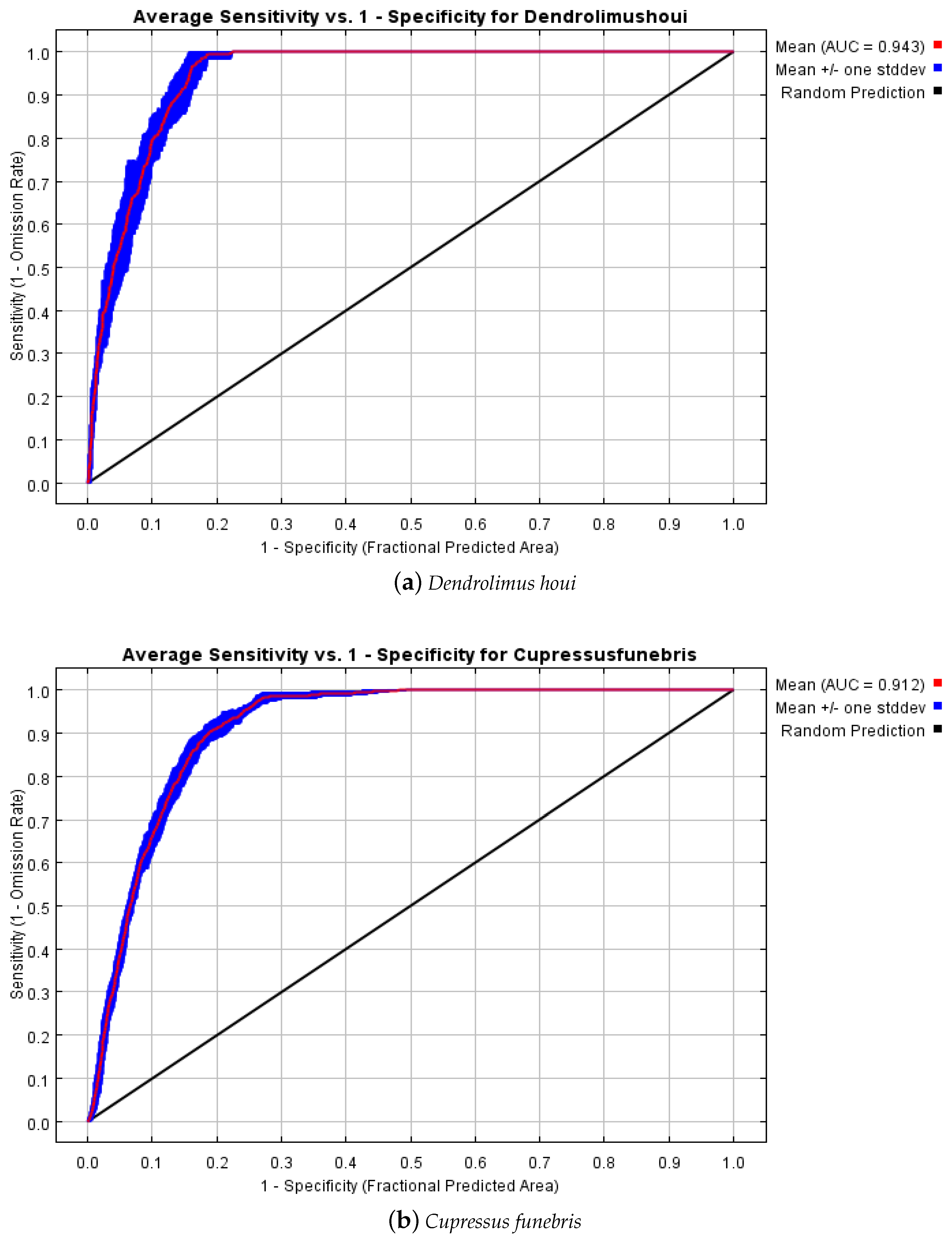
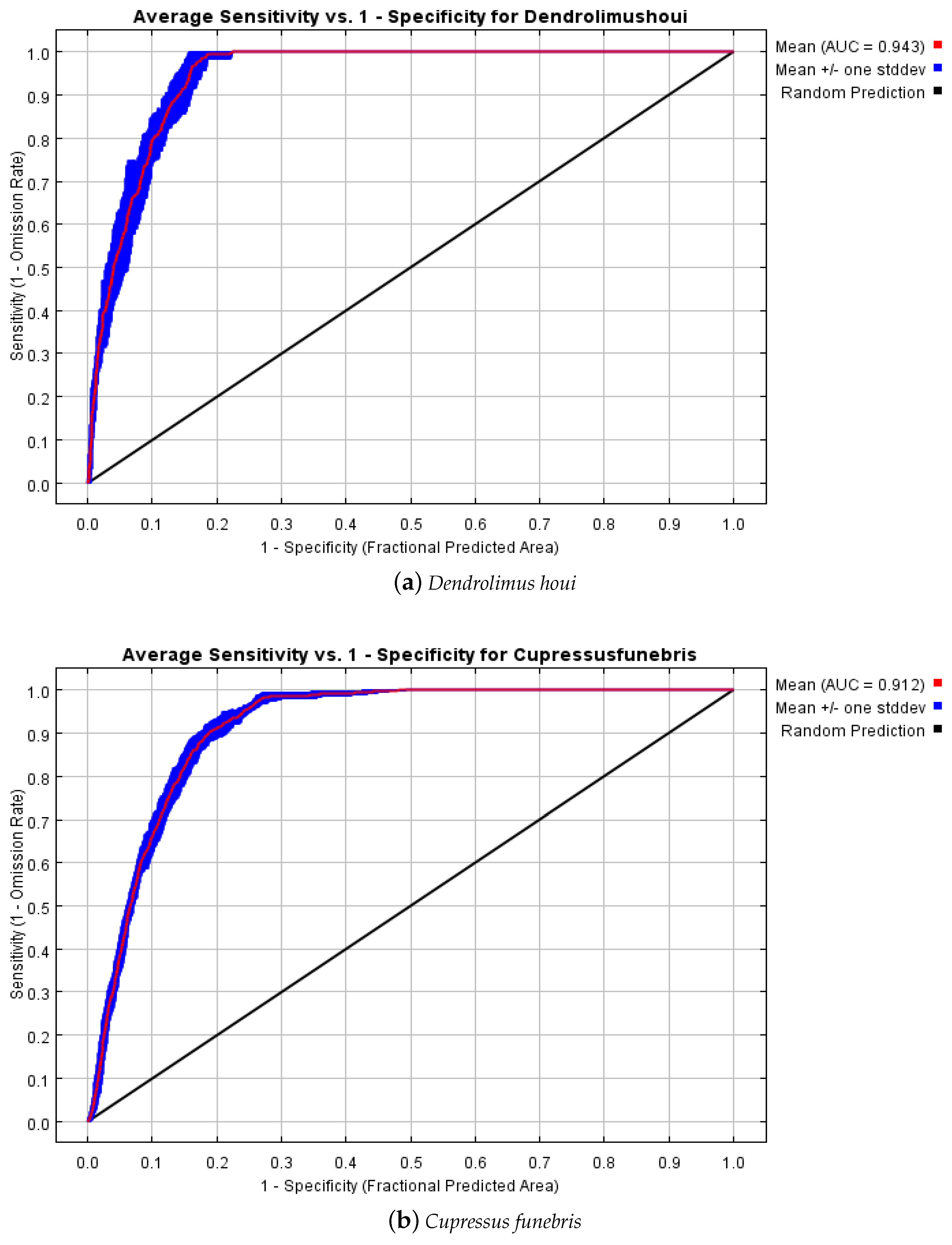
3.1. Model Optimisation and Accuracy
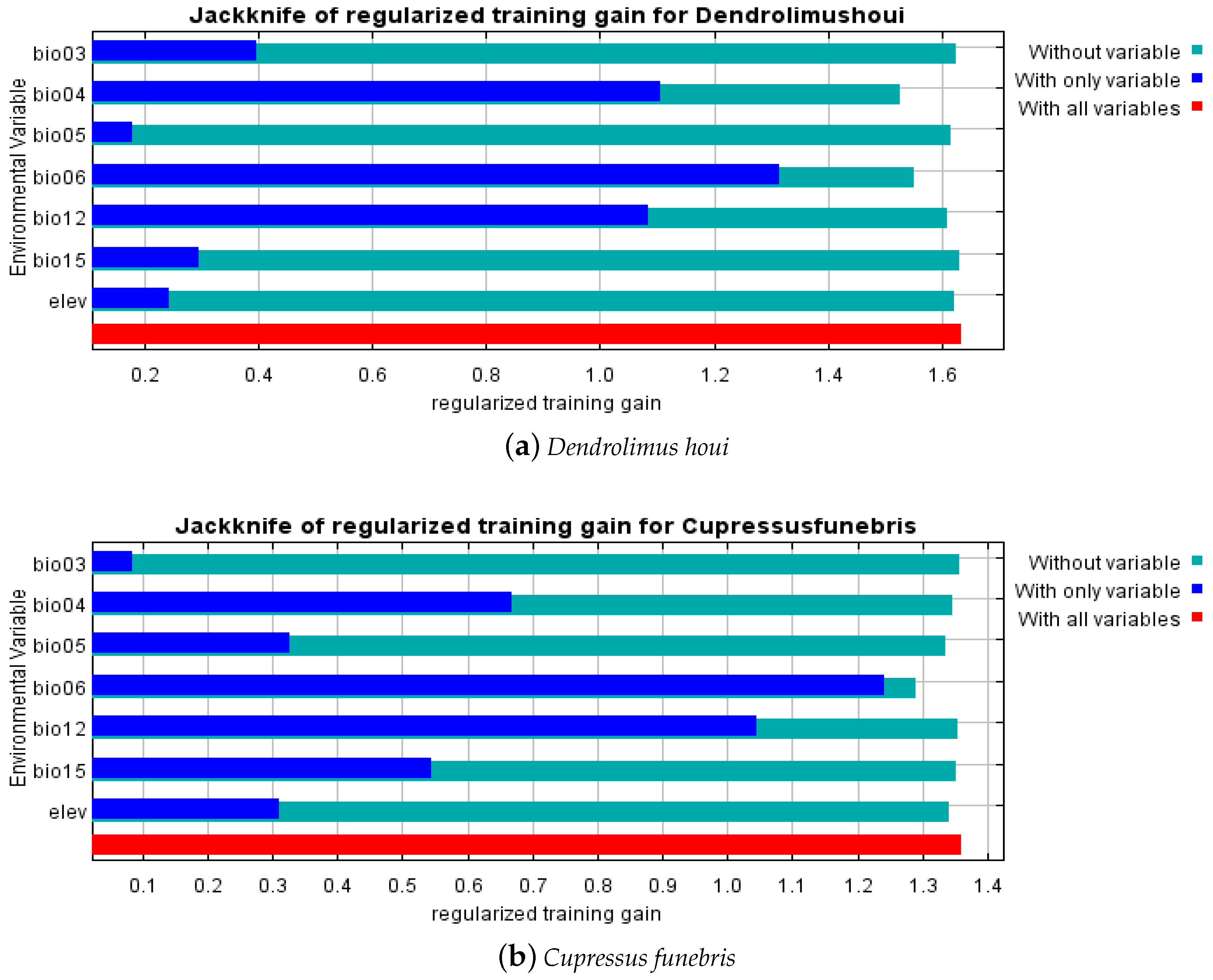
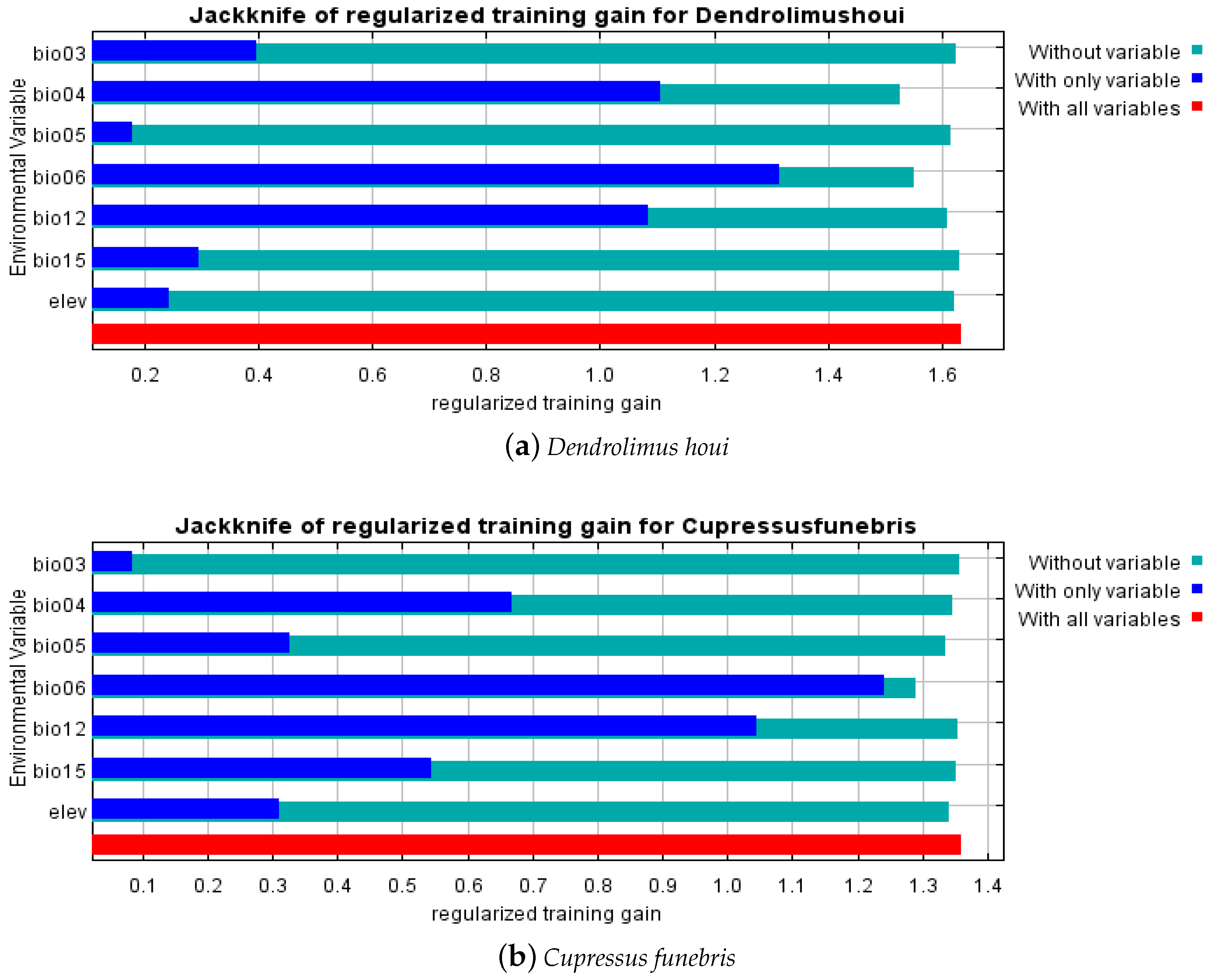
3.2. Critical Environmental Factors
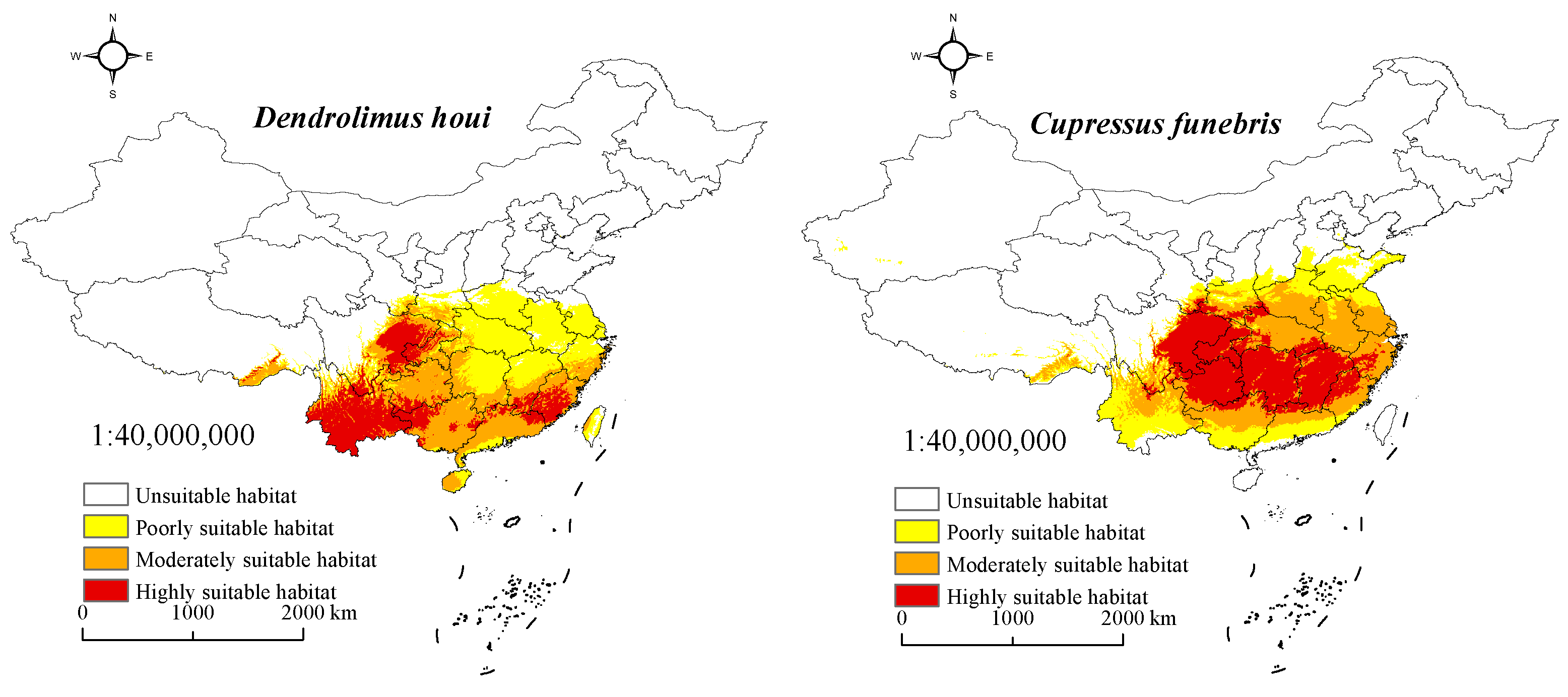
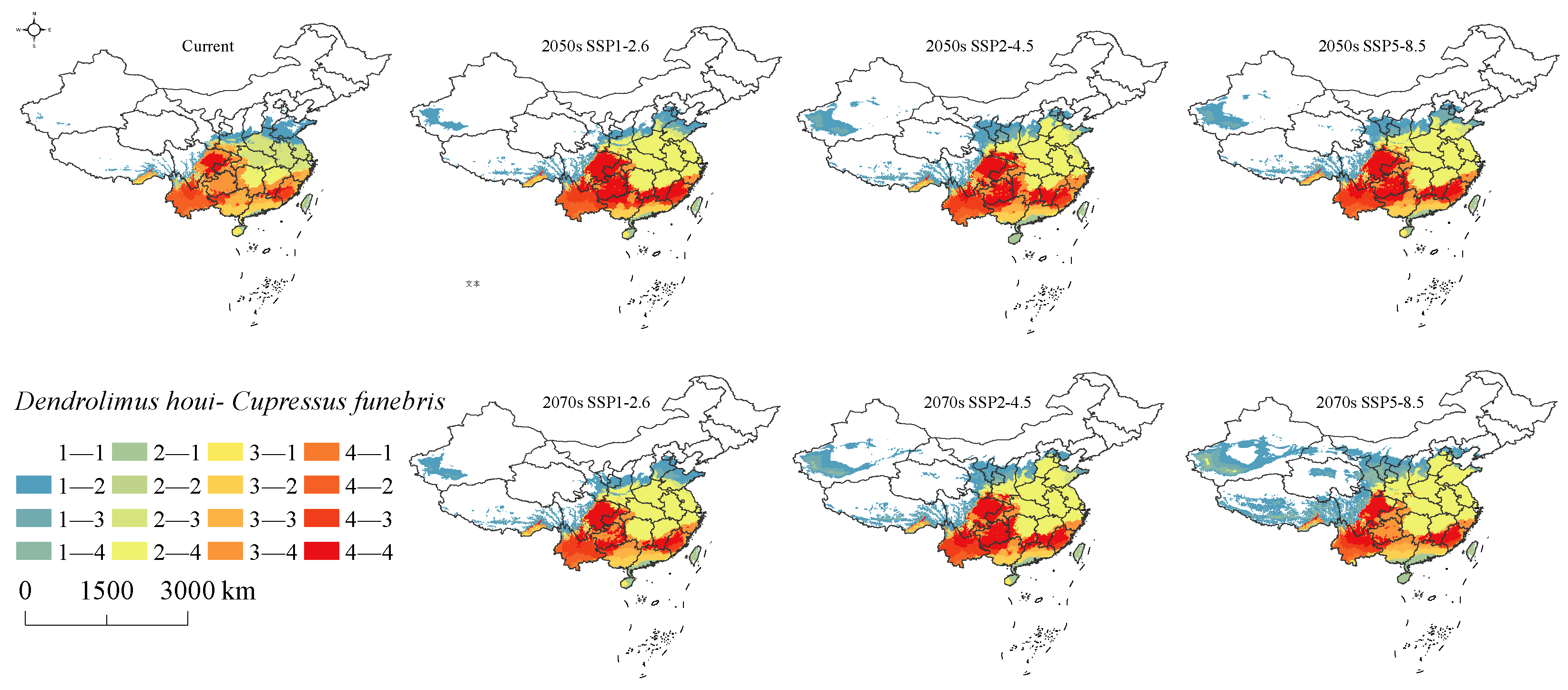
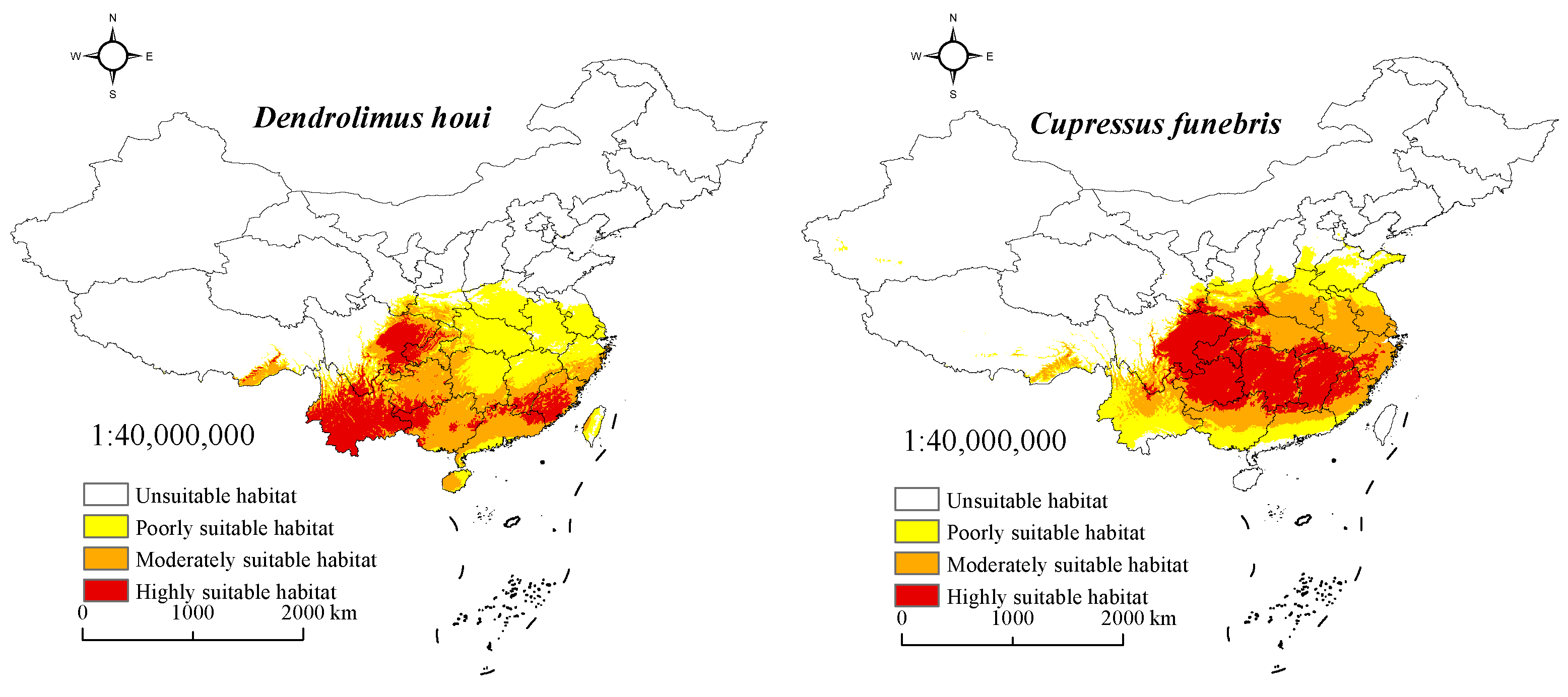
3.3. Suitable Habitat for D. houi and Its Host under Current Climate Scenarios
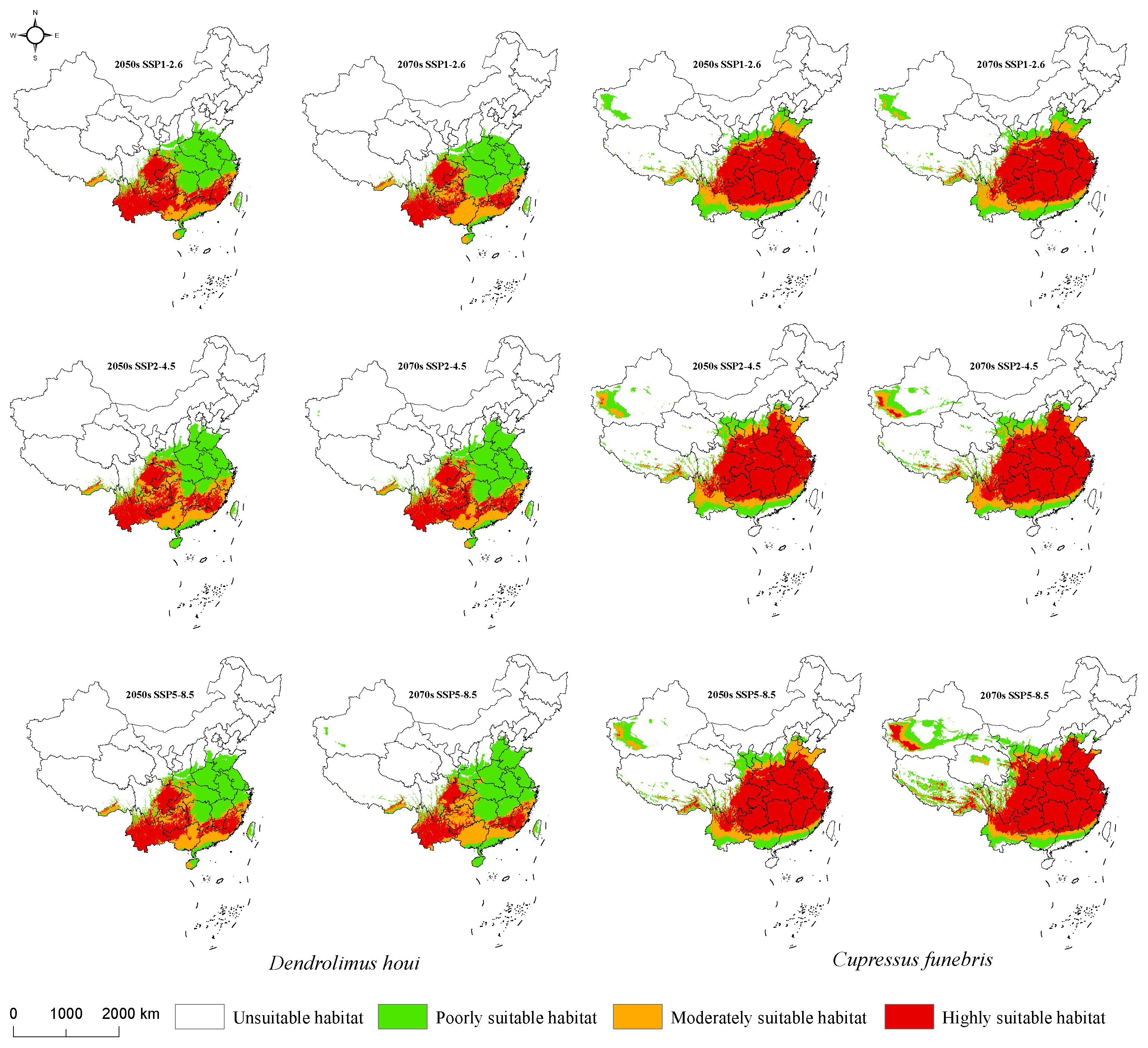
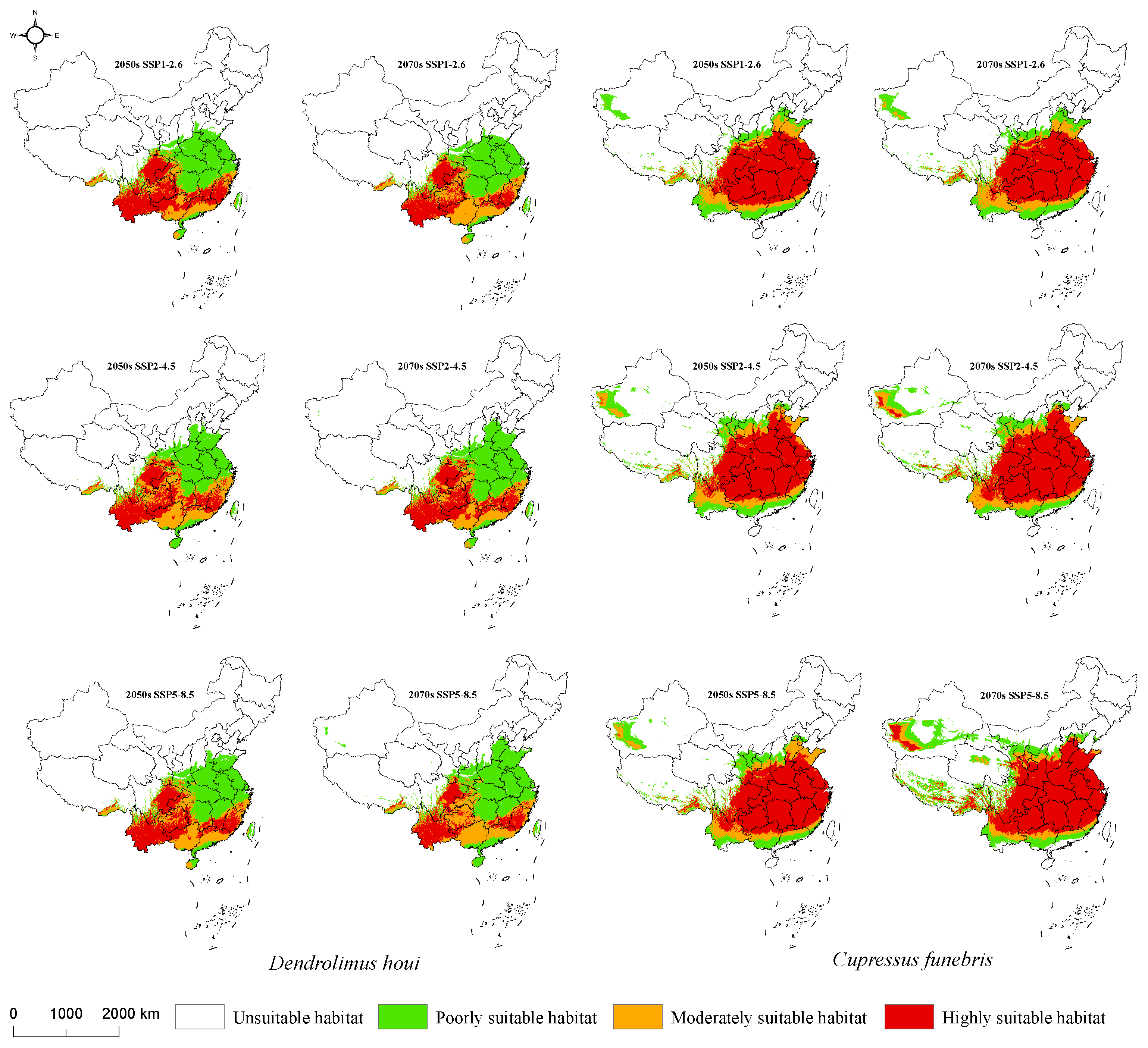
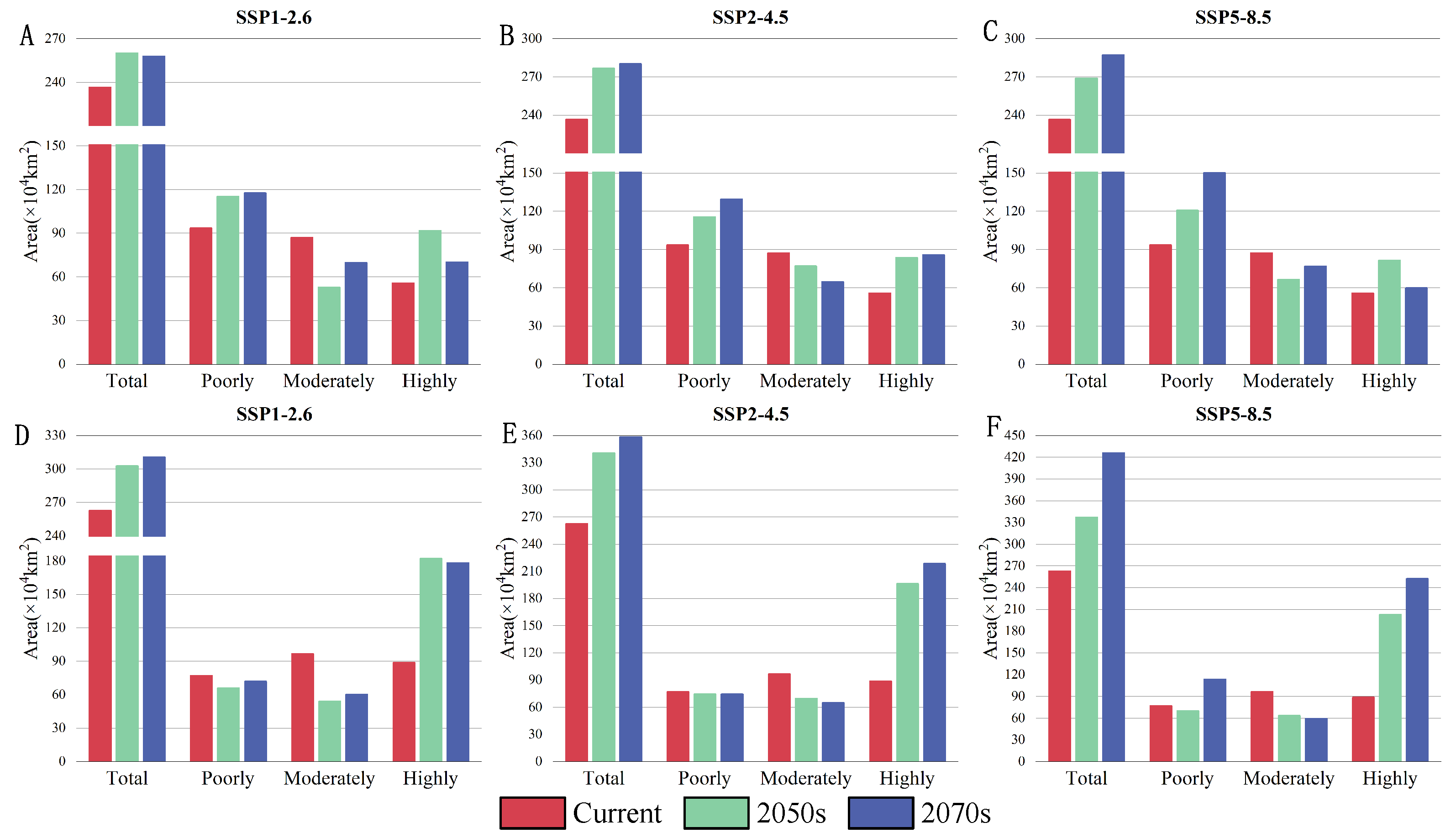
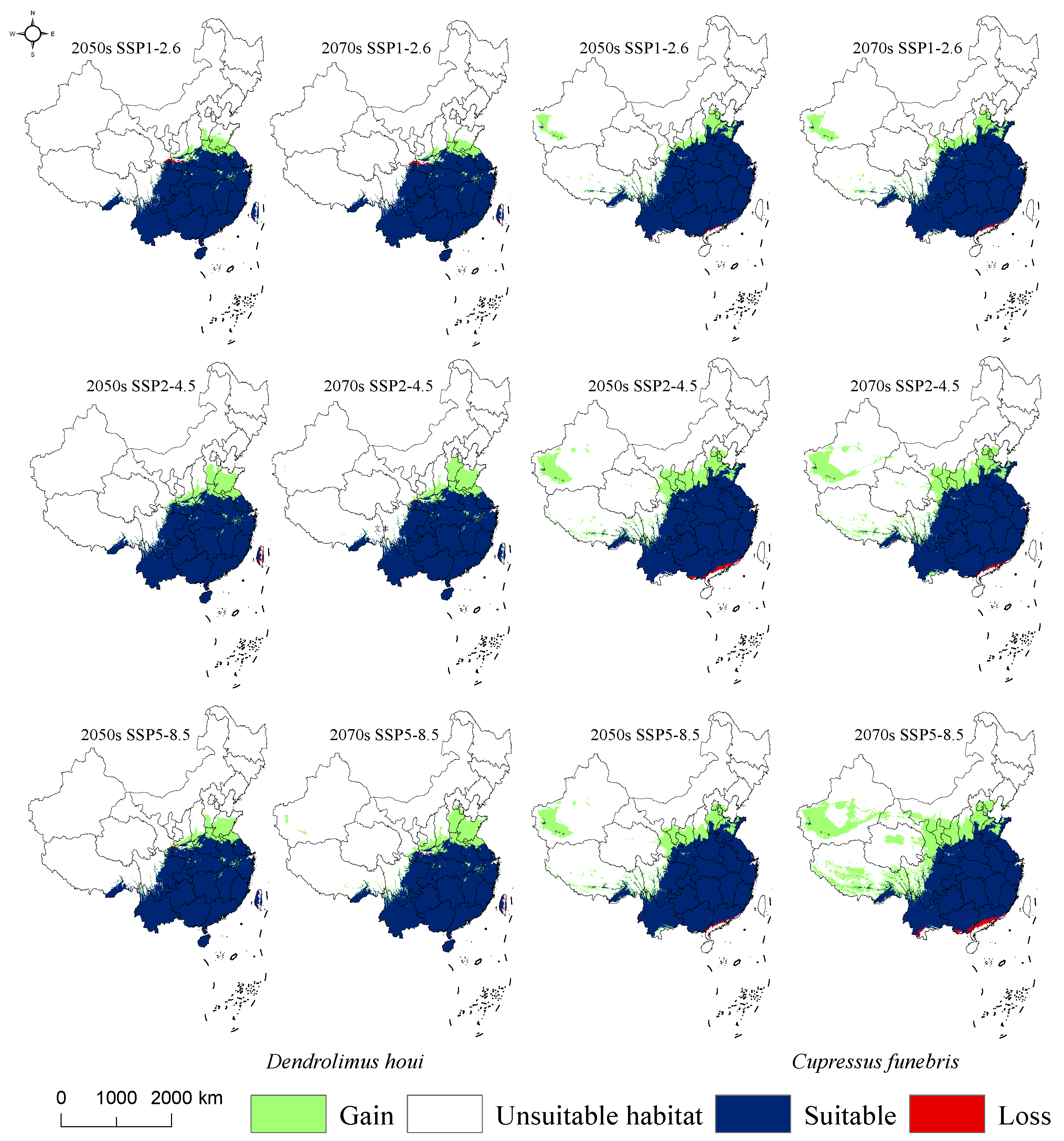
3.4. Suitable Habitat for D. houi and Its Host under Future Climate Scenarios
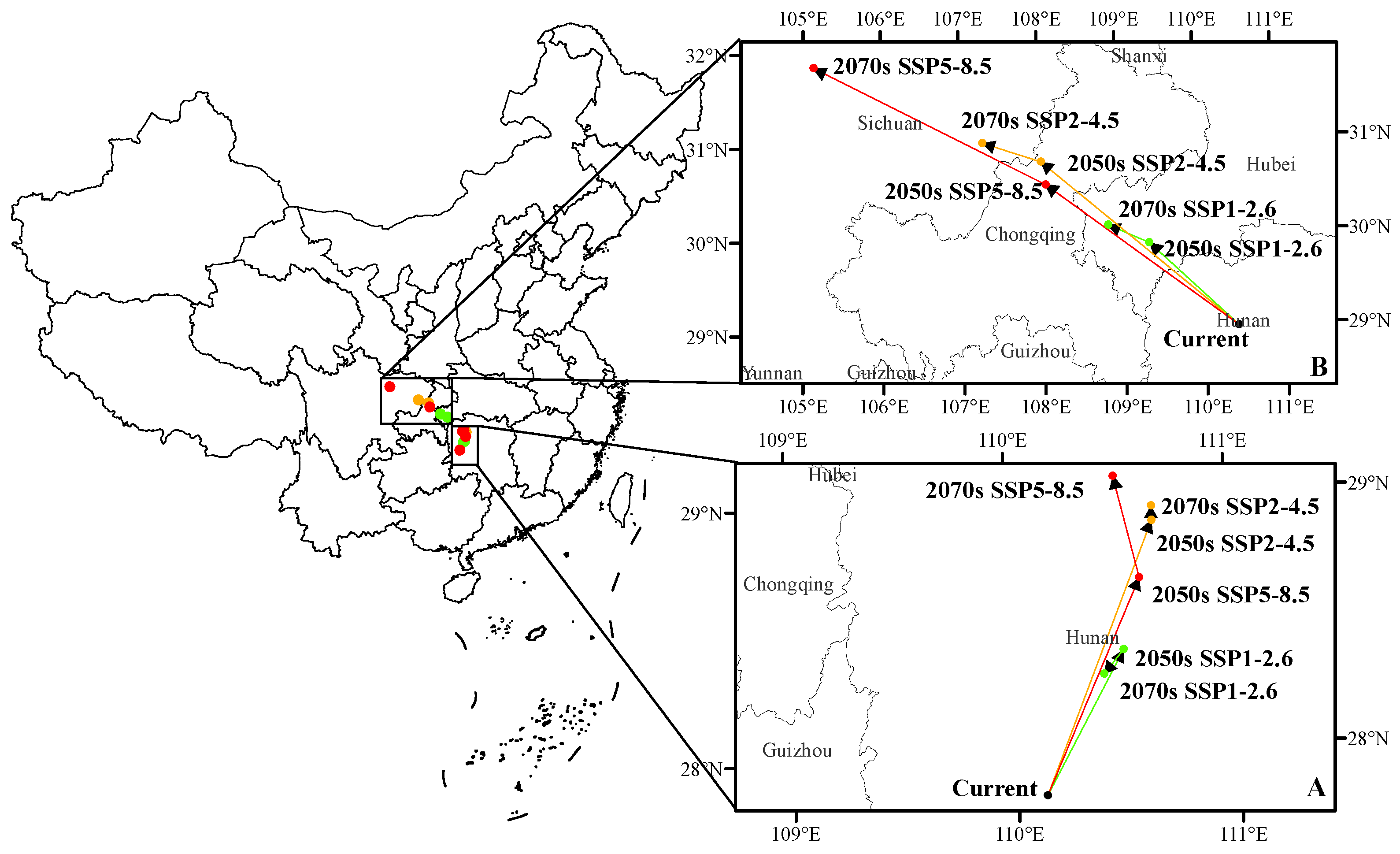
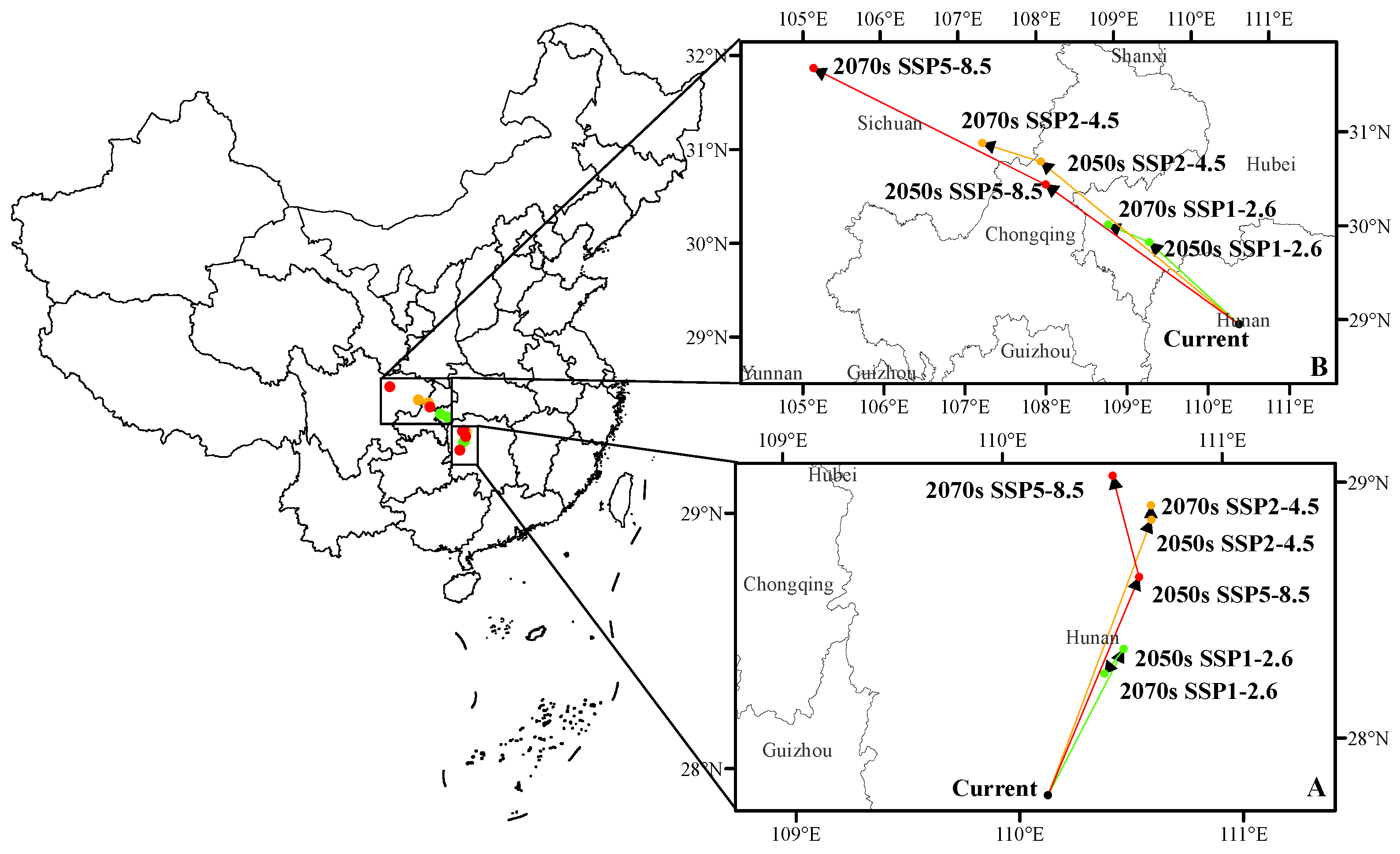
3.5. The Shift of Suitable Habitat Center
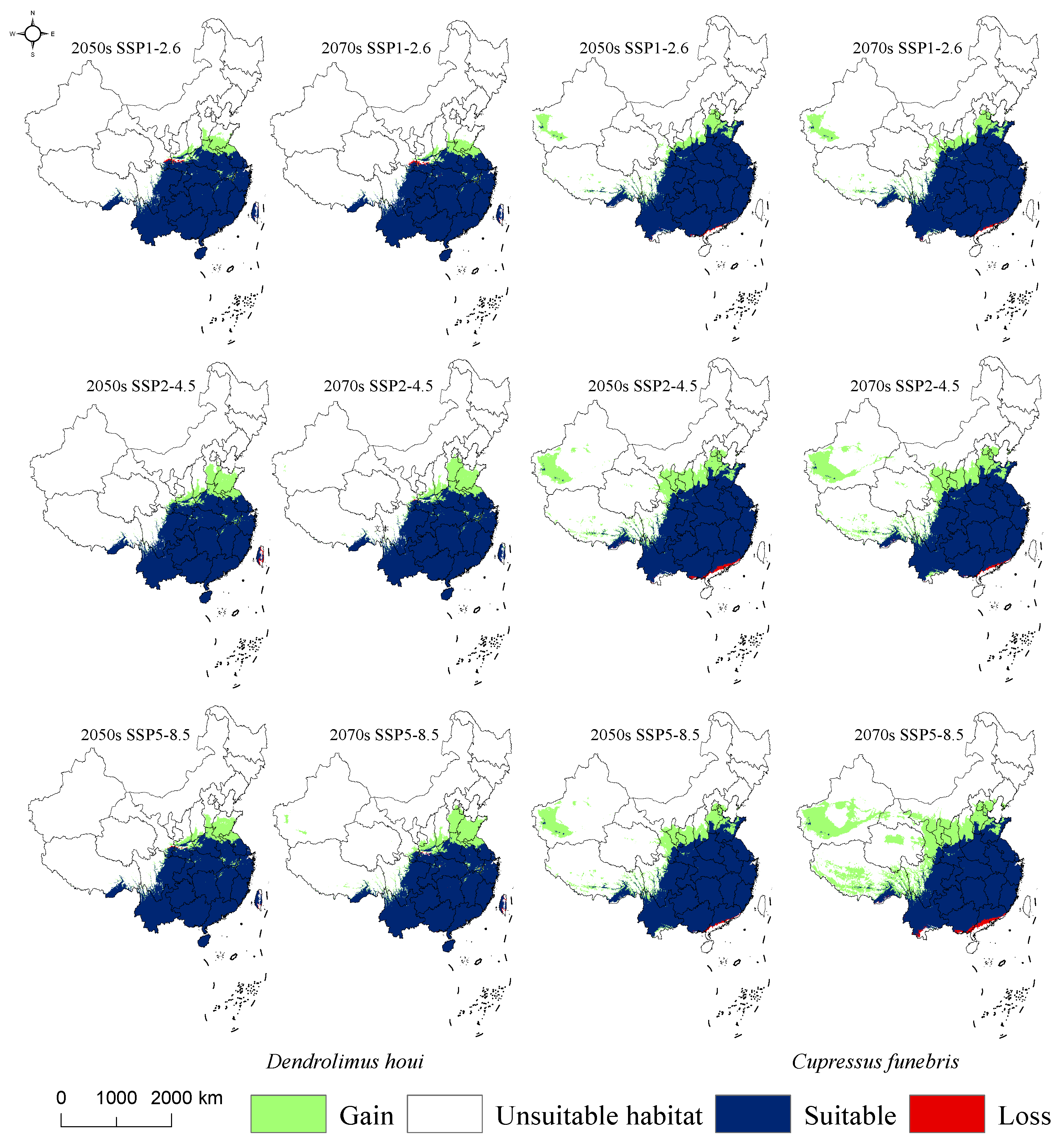
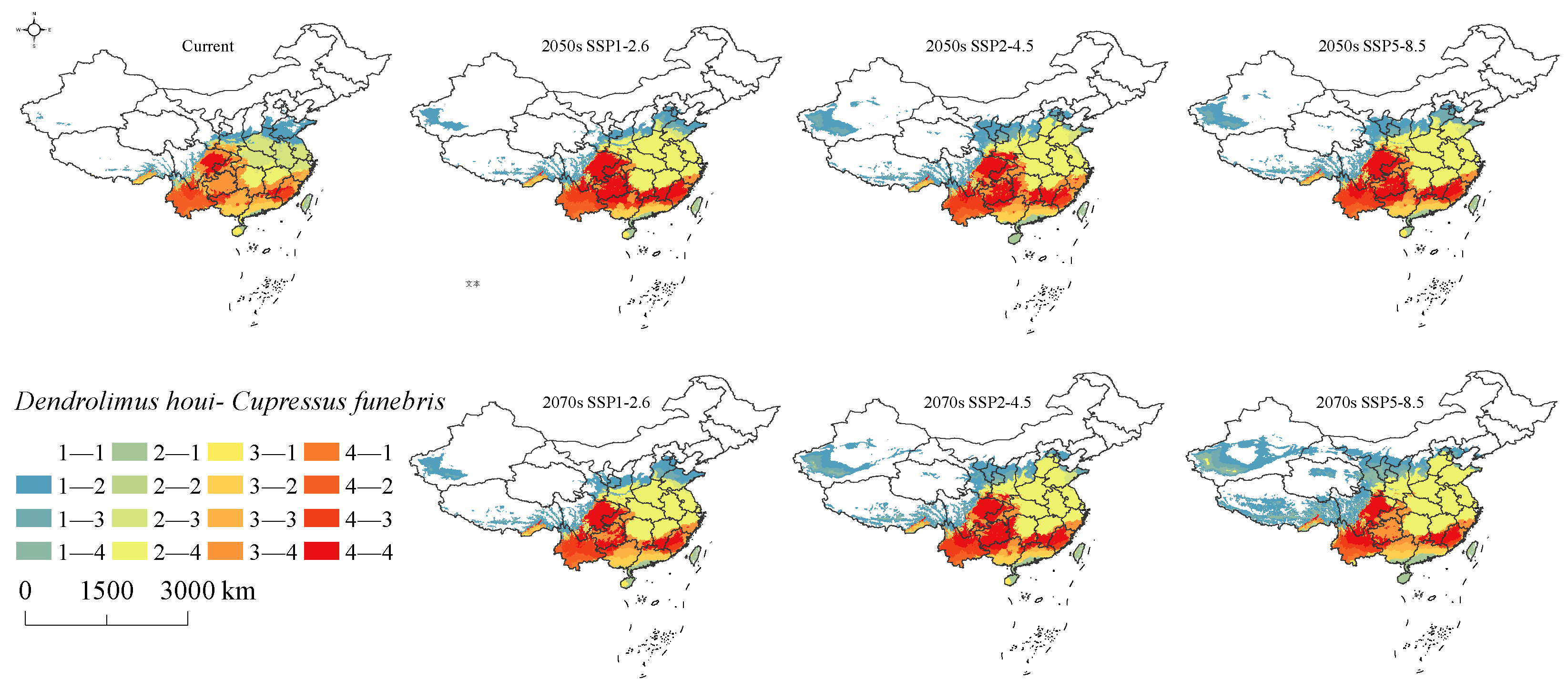
3.6. Changes in Intersecting Regions
4. Discussion
4.1. Reliability of Simulation Results
4.2. Influence of Climatic Factors
4.3. Changes of Suitable Habitat
4.4. Implications for Conservation Planning
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Song, Y.; Dong, Y.; Cui, D.; Yue, F.; Zhu, N.; Bai, H. Dynamic analysis of forestry pest species in my country V. Insects. Chin. For. Dis. Pests 2019, 38, 26–30. [Google Scholar]
- Hua, Y.; Lin, H.; Lu, C.; Shi, Y.; He, H.; Han, X.; Zhang, F.; Liang, G. Research advances in Dendrolimus houi Lajonquiere of China. World For. Res. 2019, 32, 62–68. [Google Scholar]
- Shen, H.; Liang, Y.; Chen, Z.; Hua, Y.; Li, C.; Zhang, Z.; Weng, H.; Huang, B.; Zhang, F.; Liang, G. Effects of Different Host Plants on the Growth, Reproduction and Physiological Enzyme Activity of Dendrolimus houi Lajonquiere (Lepidoptera: Lasiocampidae). For. Res. 2022, 35, 63–70. [Google Scholar]
- Zhang, S.; Kong, X.; Zhang, Z. Research Progress on the Dendrolimus spp. pheromone: From identification to molecular recognition. Front. Cell Dev. Biol. 2022, 10, 829826. [Google Scholar] [CrossRef]
- Kong, X.B.; Zhang, Z.; Zhao, C.H.; Wang, H.B. Female sex pheromone of the Yunnan pine caterpillar moth Dendrolimus houi: First (E, Z)-isomers in pheromone components of Dendrolimus spp. J. Chem. Ecol. 2007, 33, 1316–1327. [Google Scholar] [CrossRef]
- Stephens, P.A.; Mason, L.R.; Green, R.E.; Gregory, R.D.; Sauer, J.R.; Alison, J.; Aunins, A.; Brotons, L.; Butchart, S.H.; Campedelli, T.; et al. Consistent response of bird populations to climate change on two continents. Science 2016, 352, 84–87. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Palmer, G.; Platts, P.J.; Brereton, T.; Chapman, J.W.; Dytham, C.; Fox, R.; Pearce-Higgins, J.W.; Roy, D.B.; Hill, J.K.; Thomas, C.D. Climate change, climatic variation and extreme biological responses. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160144. [Google Scholar] [CrossRef]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.M.B.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of IPCC the Intergovernmental Panel on Climate Change. 2014, Volume 18, pp. 95–123. Available online: http://www.ipcc.ch/publications_and_data/publications_ipcc_fourth_assessment_report_wg1_report_the_physical_science_basis.htm (accessed on 18 April 2023).
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, R.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.; Plattner, G.K.; Rogelj, J.; et al. Climate Change 2021: The physical science basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Technical Summary. 2021. Available online: https://www.ipcc.ch/report/ar6/wg1/chapter/technical-summary/ (accessed on 18 April 2023).
- Lin, L.; He, J.; Xie, L.; Cui, G. Prediction of the suitable area of the Chinese white pines (Pinus subsect. strobus) under climate changes and implications for their conservation. Forests 2020, 11, 996. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Wang, Y.C.; Feng, C.C.; Wan, P.H.M.; Chang, K.T.T. Potential distributional changes of invasive crop pest species associated with global climate change. Appl. Geogr. 2017, 82, 83–92. [Google Scholar] [CrossRef]
- Ning, H.; Ling, L.; Sun, X.; Kang, X.; Chen, H. Predicting the future redistribution of Chinese white pine Pinus armandii Franch. Under climate change scenarios in China using species distribution models. Glob. Ecol. Conserv. 2021, 25, e01420. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J. Global change sharpens the double-edged sword effect of aquatic alien plants in China and beyond. Front. Plant Sci. 2019, 10, 787. [Google Scholar] [CrossRef]
- Kumar, D.; Rawat, S. Modeling the effect of climate change on the distribution of threatened medicinal orchid Satyrium nepalense D. Don in India. Environ. Sci. Pollut. Res. 2022, 29, 72431–72444. [Google Scholar] [CrossRef]
- Ouyang, X.; Bai, S.; Strachan, G.B.; Chen, A. Simulation of the potential distribution of rare and endangered Satyrium species in China under climate change. Ecol. Evol. 2022, 12, e9054. [Google Scholar] [CrossRef]
- Hou, J.; Xiang, J.; Li, D.; Liu, X. Prediction of Potential Suitable Distribution Areas of Quasipaa spinosa in China Based on MaxEnt Optimization Model. Biology 2023, 12, 366. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef]
- Scott, L.M.; Janikas, M.V. Spatial statistics in ArcGIS. In Handbook of Applied Spatial Analysis: Software Tools, Methods and Applications; Springer: Berlin/Heidelberg, Germany, 2009; pp. 27–41. [Google Scholar]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Tebaldi, C.; Van Vuuren, D.P.; Eyring, V.; Friedlingstein, P.; Hurtt, G.; Knutti, R.; Kriegler, E.; Lamarque, J.F.; Lowe, J.; et al. The scenario model intercomparison project (ScenarioMIP) for CMIP6. Geosci. Model Dev. 2016, 9, 3461–3482. [Google Scholar] [CrossRef]
- Wu, T.; Song, L.; Li, W.; Wang, Z.; Zhang, H.; Xin, X.; Zhang, Y.; Zhang, L.; Li, J.; Wu, F.; et al. An overview of BCC climate system model development and application for climate change studies. J. Meteorol. Res. 2014, 28, 34–56. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Tarabon, S.; Bergès, L.; Dutoit, T.; Isselin-Nondedeu, F. Environmental impact assessment of development projects improved by merging species distribution and habitat connectivity modelling. J. Environ. Manag. 2019, 241, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Akaike, H. Information Theory as an Extension of the Maximum likelihood. International Symposium on Information Theory. Available online: https://inspirehep.net/literature/1795438 (accessed on 26 April 2023).
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Guo, R.; Deser, C.; Terray, L.; Lehner, F. Human influence on winter precipitation trends (1921–2015) over North America and Eurasia revealed by dynamical adjustment. Geophys. Res. Lett. 2019, 46, 3426–3434. [Google Scholar] [CrossRef]
- Simpson, I.R.; Yeager, S.G.; McKinnon, K.A.; Deser, C. Decadal predictability of late winter precipitation in western Europe through an ocean–jet stream connection. Nat. Geosci. 2019, 12, 613–619. [Google Scholar] [CrossRef]
- Dale, A.G.; Frank, S.D. Warming and drought combine to increase pest insect fitness on urban trees. PLoS ONE 2017, 12, e0173844. [Google Scholar] [CrossRef]
- Tang, X.; Yuan, Y.; Li, X.; Zhang, J. Maximum entropy modeling to predict the impact of climate change on pine wilt disease in China. Front. Plant Sci. 2021, 12, 652500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.S. Relationship Between Occurrence Period of Pine Moth and Meteorological Conditions. Chin. J. Agrometeorol. 2004, 25, 26–28. [Google Scholar]
- Dawei, L. Meteorological Condition Analysis and Trend Prediction of Dendrolimus houi Occurrence in Lishui City. J. Anhui Agric. Sci. 2008, 36, 1106. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Environmental Factors | Description of Environmental Factors |
|---|---|---|
| Description of Environmental Factors | Bio1 | Annual Mean Temperature |
| Bio2 | Mean Diurnal Range | |
| Bio3 * | Isothermality | |
| Bio4 * | Temperature Seasonality | |
| Bio5 * | Max Temperature of Warmest Month | |
| Bio6 * | Min Temperature of Coldest Month | |
| Bio7 | Temperature Annual Range | |
| Bio8 | Mean Temperature of Wettest Quarter | |
| Bio9 | Mean Temperature of Driest Quarter | |
| Bio10 | Mean Temperature of Warmest Quarter | |
| Bio11 | Mean Temperature of Coldest Quarter | |
| Precipitation | Bio12 * | Annual Precipitation |
| Bio13 | Precipitation of Wettest Month | |
| Bio14 | Precipitation of Driest Month | |
| Bio15 * | Precipitation Seasonality | |
| Bio16 | Precipitation of Wettest Quarter | |
| Bio17 | Precipitation of Driest Quarter | |
| Bio18 | Precipitation of Warmest Quarter | |
| Bio19 | Precipitation of Coldest Quarter | |
| Terrain | elev * | Elevation |
| Species | Type | RM | FC | delta.AICc |
|---|---|---|---|---|
| Dendrolimus houi | Default | 1 | LQPH | 269.46 |
| Optimisation | 3.4 | QTH | 0 | |
| Cupressus funebris | Default | 1 | LQPH | 132.67 |
| Optimization | 0.5 | LQ | 0 |
| Variable | Environmental Variable | Perecent Contribution | |
|---|---|---|---|
| Dendrolimus houi | Cupressus funebris | ||
| Bio3 | Isothermality | 4.5 | 3.1 |
| Bio4 | Temperature Seasonality | 24 | 1.8 |
| Bio5 | Max Temperature of Warmest Month | 0.7 | 1.7 |
| Bio6 | Min Temperature of Coldest Month | 65.8 | 91.2 |
| Bio12 | Annual Precipitation | 3 | 0.4 |
| Bio15 | Precipitation Seasonality | 0.1 | 0.4 |
| Elev | Elevation | 2 | 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, G.; Zhao, Y.; Wang, Y.; Yu, C.; Xiong, F.; Sun, Y.; Cao, Y. Suitable Habitat Prediction and Analysis of Dendrolimus houi and Its Host Cupressus funebris in the Chinese Region. Forests 2024, 15, 162. https://doi.org/10.3390/f15010162
Miao G, Zhao Y, Wang Y, Yu C, Xiong F, Sun Y, Cao Y. Suitable Habitat Prediction and Analysis of Dendrolimus houi and Its Host Cupressus funebris in the Chinese Region. Forests. 2024; 15(1):162. https://doi.org/10.3390/f15010162
Chicago/Turabian StyleMiao, Guangting, Youjie Zhao, Yijie Wang, Chunjiang Yu, Fei Xiong, Yongke Sun, and Yong Cao. 2024. "Suitable Habitat Prediction and Analysis of Dendrolimus houi and Its Host Cupressus funebris in the Chinese Region" Forests 15, no. 1: 162. https://doi.org/10.3390/f15010162
APA StyleMiao, G., Zhao, Y., Wang, Y., Yu, C., Xiong, F., Sun, Y., & Cao, Y. (2024). Suitable Habitat Prediction and Analysis of Dendrolimus houi and Its Host Cupressus funebris in the Chinese Region. Forests, 15(1), 162. https://doi.org/10.3390/f15010162