
Early Growth Characterization and C:N:P Stoichiometry in Firmiana simplex Seedlings in Response to Shade and Soil Types

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Material
2.3. Seedling Culture and Experimental Design
2.4. Measurement of Growth Parameters
2.5. Analysis of Plant and Soil Physicochemical Properties
2.6. Statistical Analyses
3. Results
3.1. Effects of Shading Treatments and Soil Types on Growth Parameters
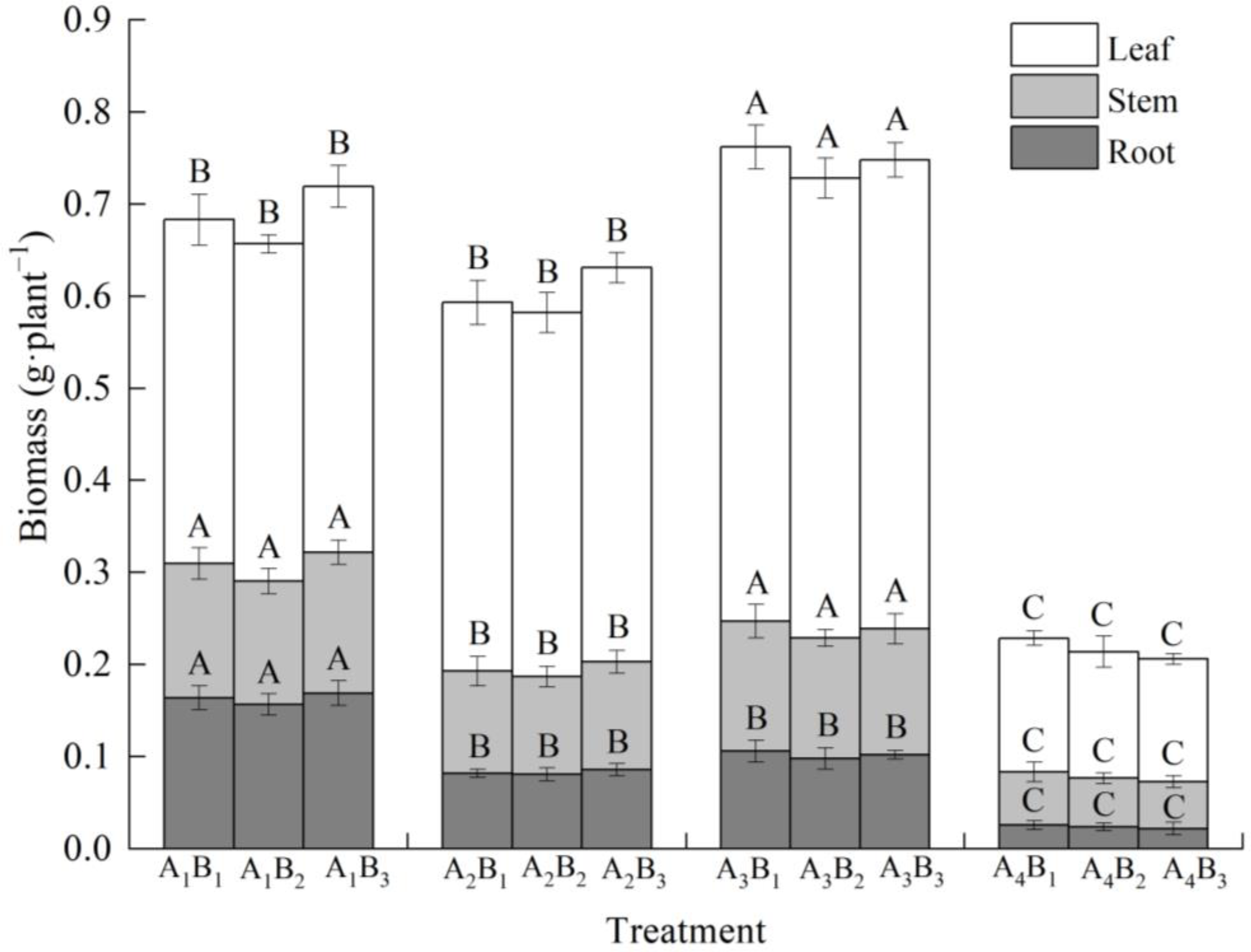
3.2. Effects of Shading Treatments and Soil Types on Biomass and Its Distribution
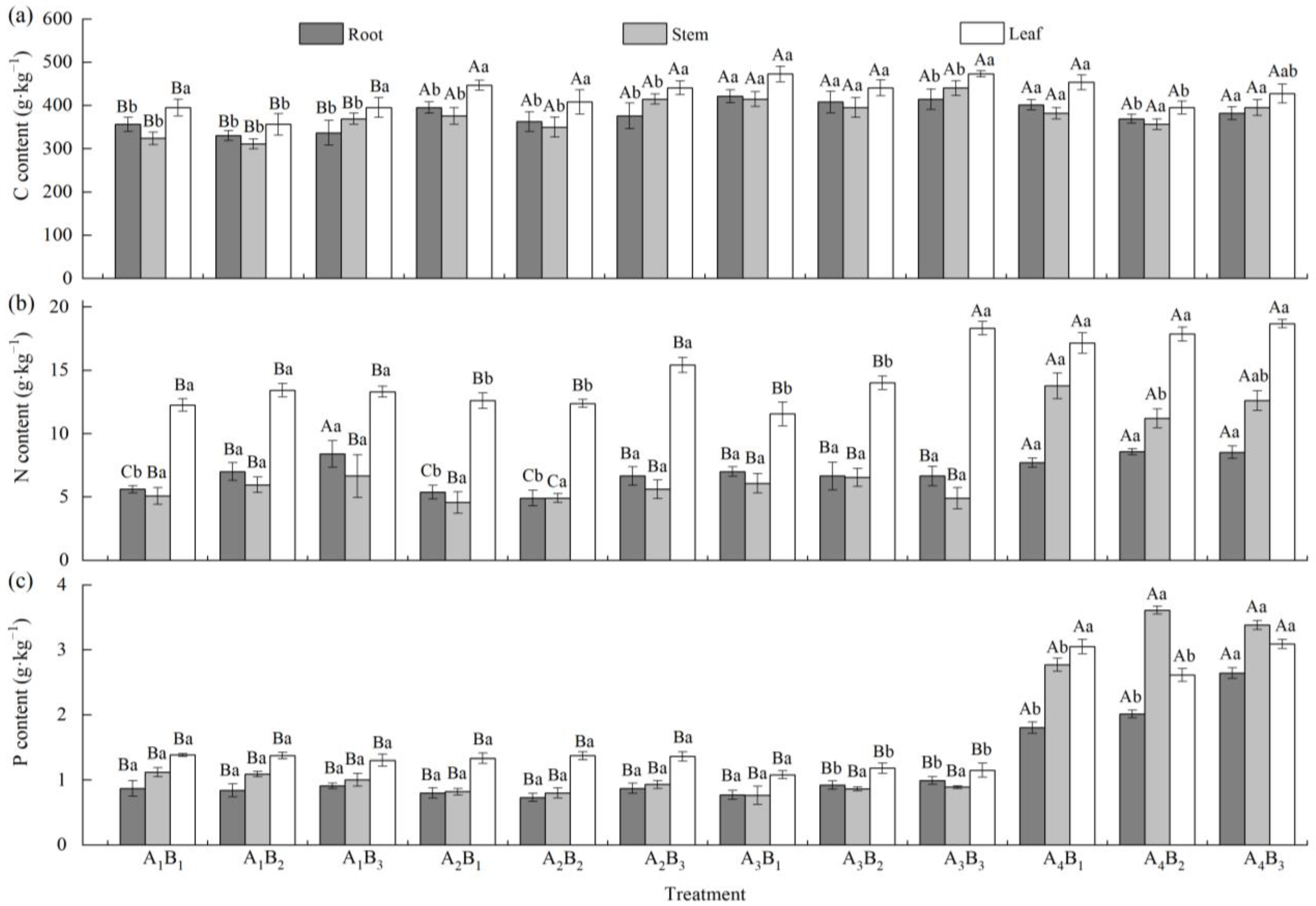
3.3. Effects of Shading Treatments and Soil Types on C, N, and P Contents
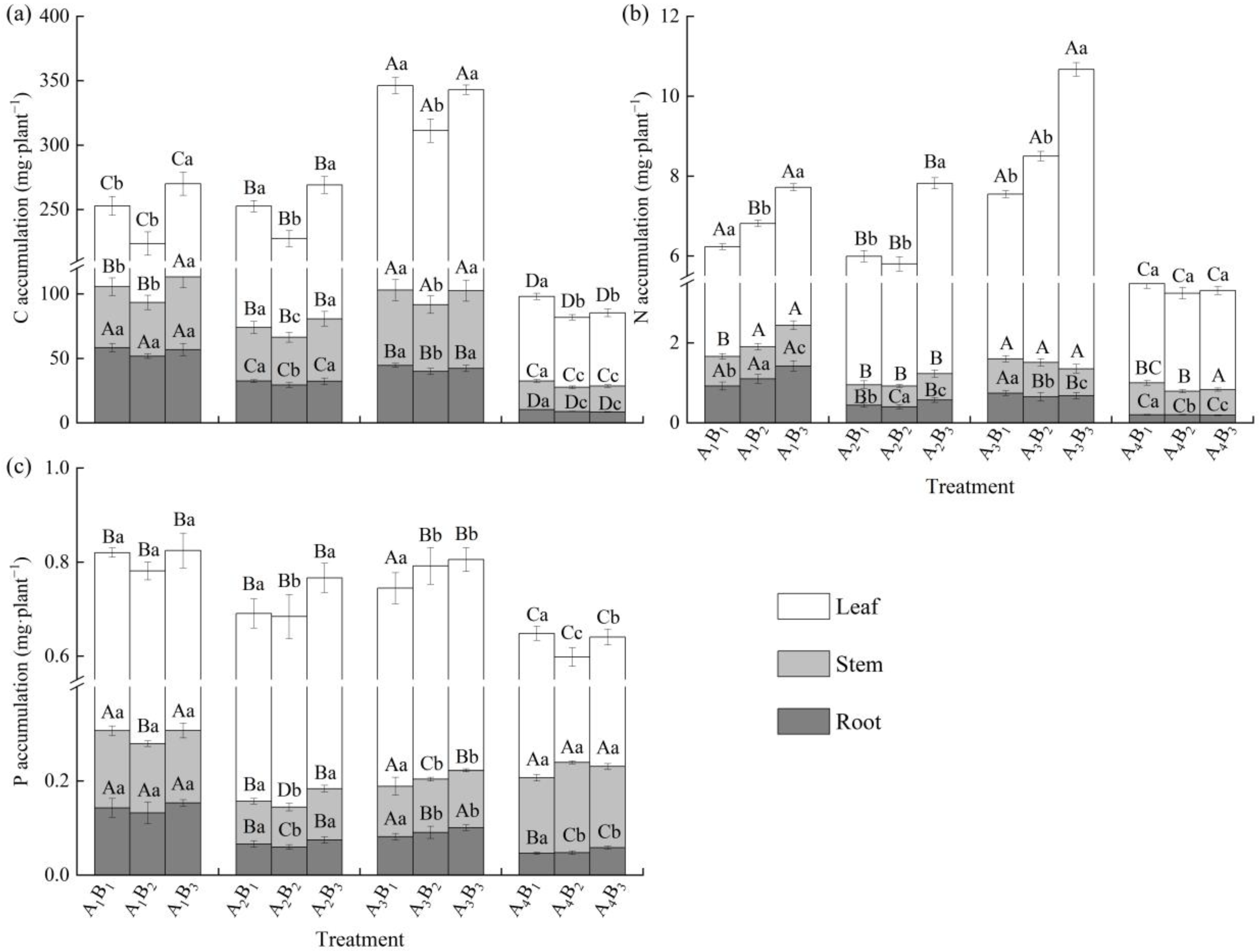
3.4. Effects of Shading Treatments and Soil Types on C, N, and P Accumulation
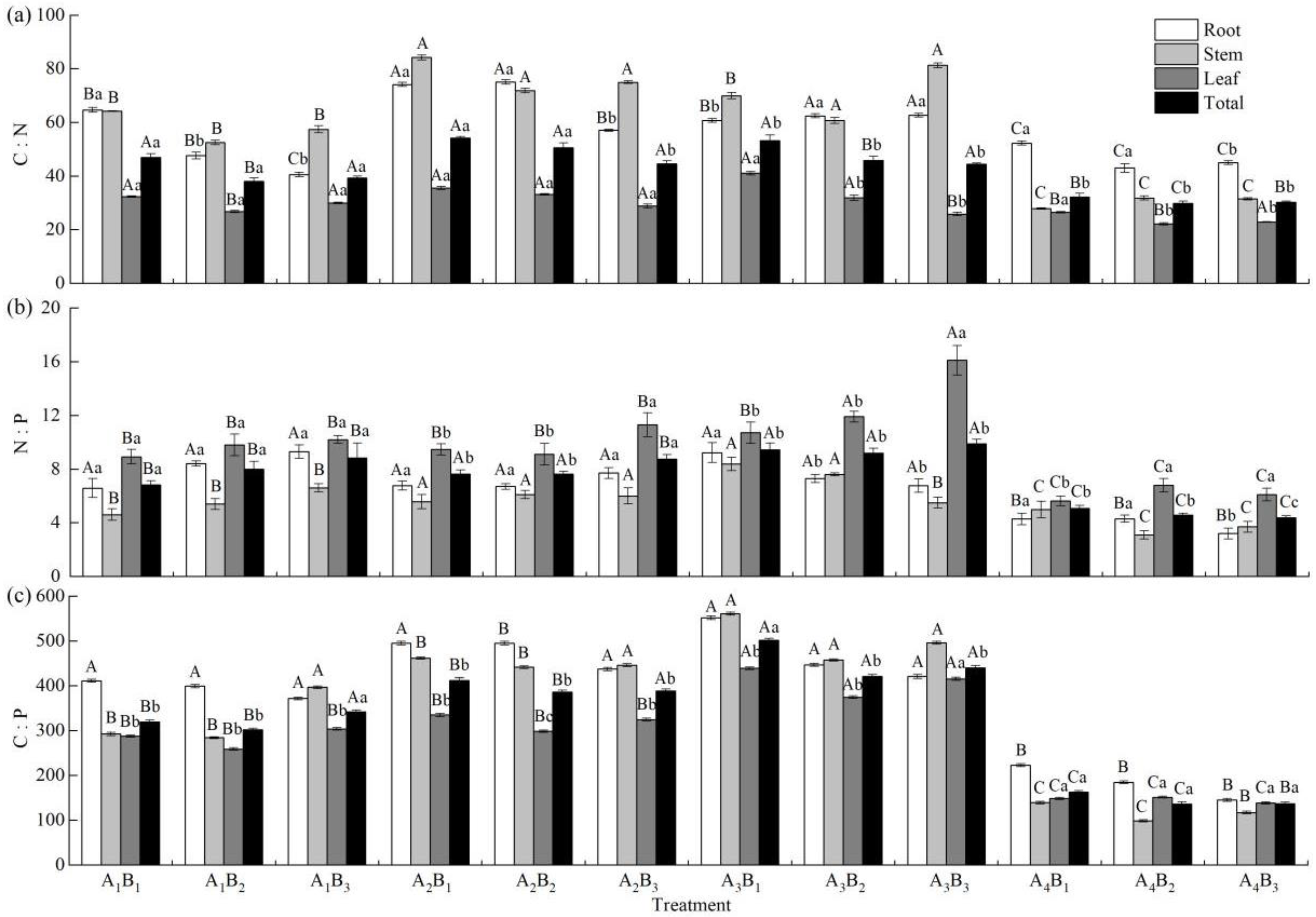
3.5. Effects of Shading Treatments and Soil Types on C, N, and P Stoichiometric Ratio
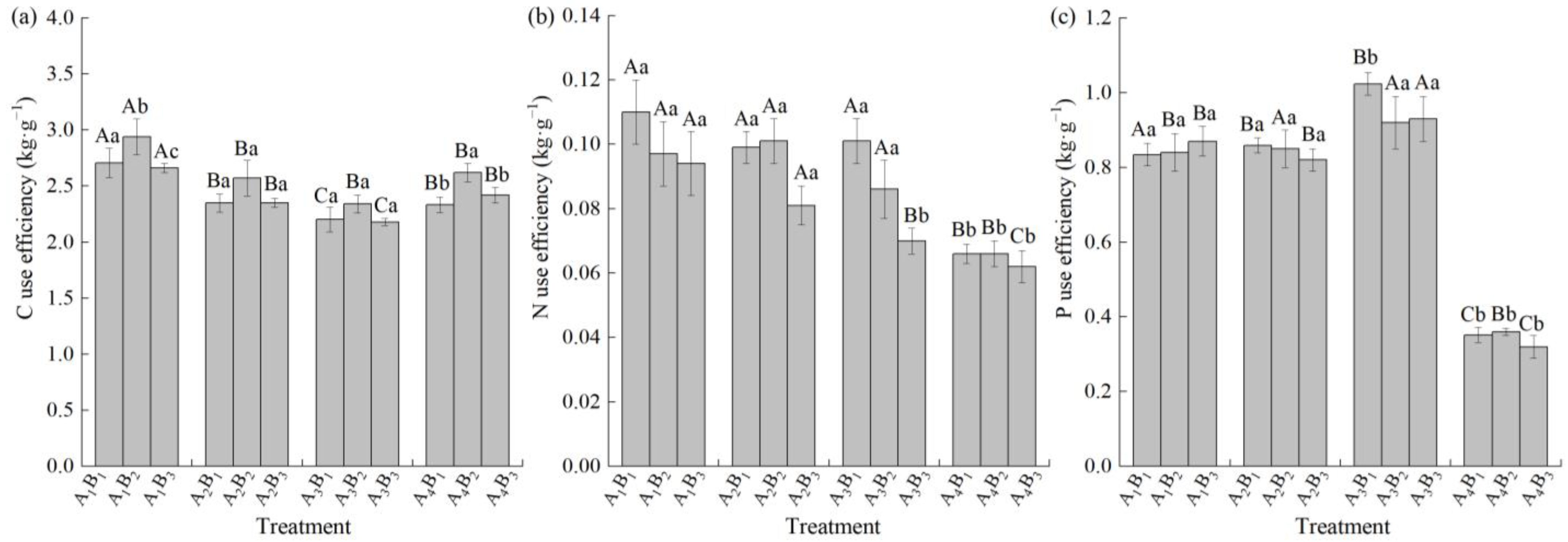
3.6. Effects of Shading Treatments and Soil Types on C, N, and P Use Efficiency
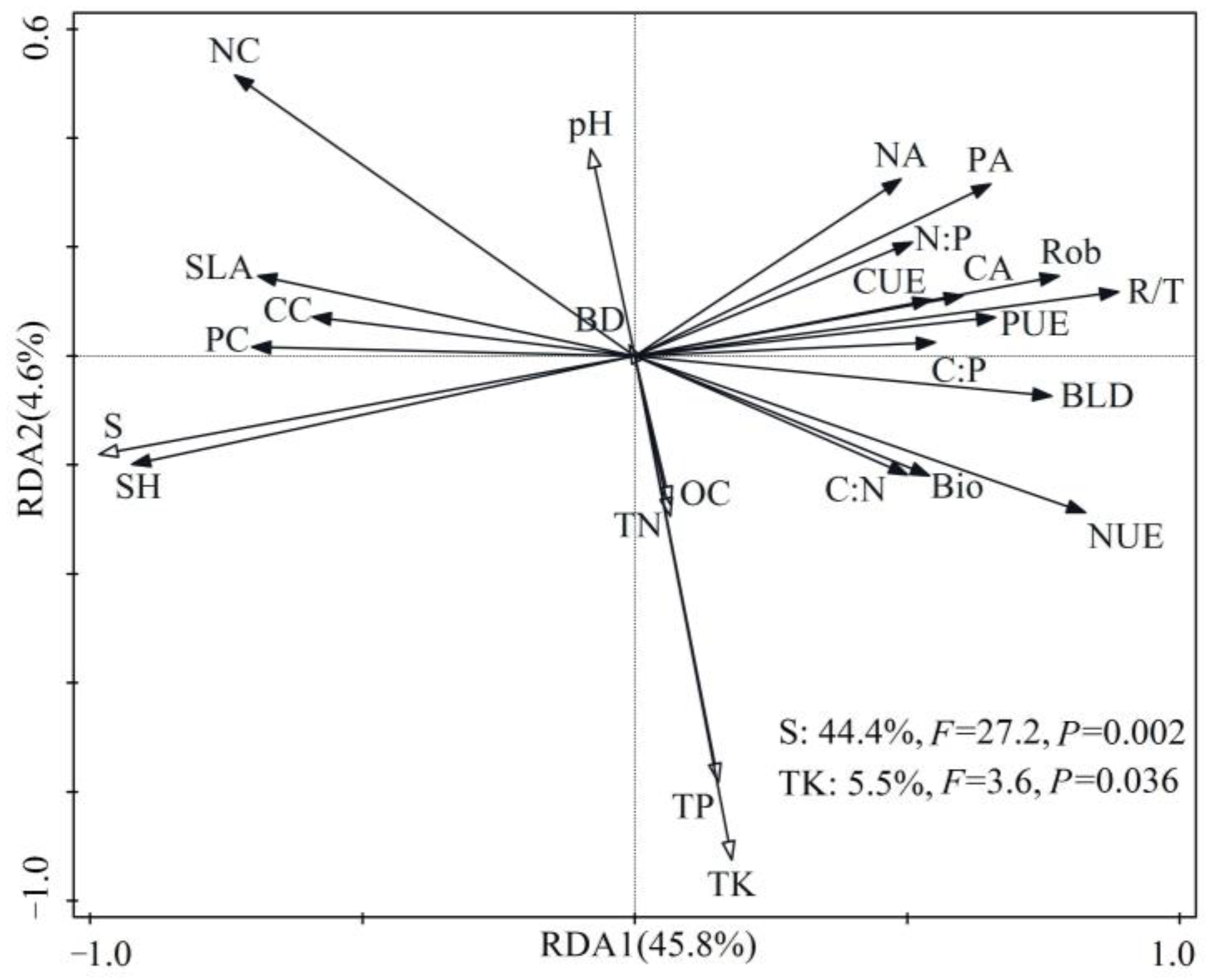
3.7. Relationships among Growth Indices, Stoichiometric Characteristics, and Environmental Factors
4. Discussion
4.1. Effects of Shading Treatments and Soil Types on Growth Performance
4.2. Effects of Shading Treatments and Soil Types on C, N, and P Contents and C:N:P Stoichiometric Characteristics
4.3. Interrelationship between Environmental Factors and Growth Parameters, Stoichiometric Characteristics
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, B.; Yang, H.; Liu, H.; Qiao, Y.; Zhang, M.; Wang, Y.; Xie, Z.; Liu, M. Effects of shading stress on grain number, yield, and photosynthesis during early reproductive growth in wheat. Crop. Sci. 2019, 59, 363–378. [Google Scholar] [CrossRef]
- Luo, G.Y.; Li, J.M.; Guo, S.L.; Li, Y.L.; Jin, Z.X. Photosynthesis, nitrogen allocation, non-structural carbohydrate allocation, and C:N:P stoichiometry of Ulmus elongata seedlings exposed to different light intensities. Life 2022, 12, 1310. [Google Scholar] [CrossRef]
- Yang, F.; Feng, L.; Liu, Q.; Wu, X.; Fan, Y.; Raza, M.A.; Cheng, Y.; Chen, J.; Wang, X.; Yong, T.; et al. Effect of interactions between light intensity and red-to- far-red ratio on the photosynthesis of soybean leaves under shade condition. Environ. Exp. Bot. 2018, 150, 79–87. [Google Scholar] [CrossRef]
- Xie, H.; Yu, M.; Cheng, X. Leaf non-structural carbohydrate allocation and C:N:P stoichiometry in response to light acclimation in seedlings of two subtropical shade-tolerant tree species. Plant Physiol. Biochem. 2018, 124, 146–154. [Google Scholar] [CrossRef]
- Liu, Y.J.; Dawson, W.; Prati, D.; Haeuser, E.; Feng, Y.H.; van Kleunen, M. Does greater specific leaf area plasticity help plants to maintain a high performance when shaded? Ann. Bot. 2016, 118, 1329–1336. [Google Scholar] [CrossRef]
- Akram, M.A.; Wang, X.; Hu, W.; Xiong, J.; Zhang, Y.; Deng, Y.; Ran, J.; Deng, J. Convergent variations in the leaf traits of desert plants. Plants 2020, 9, 990. [Google Scholar] [CrossRef]
- Akram, M.A.; Zhang, Y.; Wang, X.; Shrestha, N.; Malik, K.; Khan, I.; Ma, W.; Sun, Y.; Li, F.; Ran, J.; et al. Phylogenetic independence in the variations in leaf functional traits among different plant life forms in an arid environment. J. Plant Physiol. 2022, 272, 153671. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, Z.; Wang, Z.; Chen, Y.; Wen, Z.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 354. [Google Scholar] [CrossRef]
- Adejumo, S.A.; Owoseni, O.; Mur, L.A. Low light intensity and compost modified biochar enhanced maize growth on contaminated soil and minimized Pb induced oxidative stress. J. Environ. Chem. Eng. 2021, 9, 104764. [Google Scholar] [CrossRef]
- Wang, Y.-B.; Huang, R.-D.; Zhou, Y.-F. Effects of shading stress during the reproductive stages on photosynthetic physiology and yield characteristics of peanut (Arachis hypogaea Linn.). J. Integr. Agric. 2021, 20, 1250–1265. [Google Scholar] [CrossRef]
- Fernandez-Milmanda, G.L.; Ballare, C.L. Shade avoidance: Expanding the color and hormonepalette. Trends Plant Sci. 2021, 26, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Danyagri, G.; Dang, Q.-L. Effects of elevated carbon dioxide concentration and soil temperature on the growth and biomass responses of mountain maple (Acer spicatum) seedlings to light availability. J. Plant Ecol. 2014, 7, 535–543. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, S.; He, X.; Li, Y.; Zhang, Y.; Chen, W. Response of total phenols, flavonoids, minerals, and amino acids of four edible fern species to four shading treatments. PeerJ 2020, 8, e8354. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, T.; Liu, J.; Sun, J.; Zhang, P. Ecological stoichiometry, salt ions and homeostasis characteristics of different types of halophytes and soils. Front. Plant Sci. 2022, 13, 990246. [Google Scholar] [CrossRef] [PubMed]
- Crovo, O.; Aburto, F.; Albornoz, M.F.; Southard, R. Soil type modulates the response of C, N, P stocks and stoichiometry after native forest substitution by exotic plantations. Catena 2021, 197, 104997. [Google Scholar] [CrossRef]
- Caporale, A.G.; Vitaglione, P.; Troise, A.D.; Pigna, M.; Ruocco, M. Influence of three different soil types on the interaction of two strains of Trichoderma harzianum with Brassica rapa subsp. sylvestris cv. esculenta, under soil mineral fertilization. Geoderma 2019, 350, 11–18. [Google Scholar] [CrossRef]
- Kahkashan, P.; Najat, B.; Iram, S.; Iffat, S. Influence of soil type on the growth parameters, essential oil yield and biochemical contents of Mentha arvensis L. J. Essent. Oil Bear. Plant 2016, 19, 76–81. [Google Scholar] [CrossRef]
- Xie, J.; Yan, Q.; Yuan, J.; Li, R.; Lü, X.; Liu, S.; Zhu, J. Temporal effects of thinning on the leaf C:N:P stoichiometry of regenerated broadleaved trees in larch plantations. Forests 2020, 11, 54. [Google Scholar] [CrossRef]
- Yang, S.; Shi, J.; Chen, L.; Zhang, J.; Zhang, D.; Xu, Z.; Xiao, J.; Zhu, P.; Liu, Y.; Lin, T.; et al. Physiological and biomass partitioning shifts to water stress under distinct soil types in Populus deltoides saplings. J. Plant Ecol. 2020, 13, 545–553. [Google Scholar] [CrossRef]
- Zhi, X.; Jiang, M.; Wang, Y.; Guo, Z.; Gong, W.; Gao, S. Responses of carbon, nitrogen and phosphorus stoichiometry in different root orders of Zanthoxylum armatum seedlings to four soil types. J. Elem. 2021, 26, 953–969. [Google Scholar] [CrossRef]
- Xie, J.; Fang, H.; Zhang, Q.; Chen, M.; Xu, X.; Pan, J.; Gao, Y.; Fang, X.; Guo, X.; Zhang, L. Understory plant functional types alter stoichiometry correlations between litter and soil in Chinese fir plantations with N and P addition. Forests 2019, 10, 742. [Google Scholar] [CrossRef]
- Hu, Y.F.; Shu, X.Y.; He, J.; Zhang, Y.L.; Xiao, H.H.; Tang, X.Y.; Gu, Y.F.; Lan, T.; Xia, J.G.; Ling, J.; et al. Storage of C, N, and P affected by afforestation with Salix cupularis in an alpine semiarid desert ecosystem. Land Degrad. Dev. 2018, 29, 188–198. [Google Scholar] [CrossRef]
- Zhu, D.; Hui, D.; Wang, M.; Yang, Q.; Yu, S. Light and competition alter leaf stoichiometry of introduced species and native mangrove species. Sci. Total Environ. 2020, 738, 140301. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y.H. Plant mixture balances terrestrial ecosystem C:N:P stoichiometry. Nat. Commun. 2021, 12, 4562. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gou, X.; Zhang, F.; Bian, R.; Yin, D. Spatial patterns in the C:N:P stoichiometry in Qinghai spruce and the soil across the Qilian Mountains, China. Catena 2021, 196, 104814. [Google Scholar] [CrossRef]
- Shi, L.; Li, Q.; Fu, X.; Kou, L.; Dai, X.; Wang, H. Foliar, root and rhizospheric soil C:N:P stoichiometries of overstory and understory species in subtropical plantations. Catena 2021, 198, 105020. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Pan, F.; Li, D.; Chen, H.; Wang, K. Changes in nitrogen and phosphorus limitation during secondary succession in a karst region in southwest China. Plant Soil 2015, 391, 77–91. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Fang, Y.; Ma, R.; Lal, R.; An, S.; Huang, Y. Impact of vegetation restoration on plants and soil C:N:P stoichiometry on the Yunwu Mountain Reserve of China. Ecol. Eng. 2017, 109, 92–100. [Google Scholar] [CrossRef]
- Jing, H.; Zhou, H.; Wang, G.; Xue, S.; Liu, G.; Duan, M. Nitrogen addition changes the stoichiometry and growth rate of different organs in Pinus tabuliformis seedlings. Front. Plant Sci. 2017, 8, 1922. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, J.; Zhao, X.; Chao, Z.; Wang, Y.; Zhang, X.; Wang, D. The linkages of plant, litter and soil C:N:P stoichiometry and nutrient stock in different secondary mixed forest types in the Qinling Mountains, China. PeerJ 2020, 8, e9274. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Q.; Chang, F.; Pan, H.; Liu, X.-F.; Li, H.; Hu, D.-Y.; Yang, S. Production and fuel properties of biodiesel from Firmiana platanifolia L.f. as a potential non-food oil source. Ind. Crop. Prod. 2015, 76, 768–771. [Google Scholar] [CrossRef]
- Lv, E.; Dou, T.; Ding, S.; Lu, J.; Li, Z.; Yi, W.; Li, J.; Ding, J. Membrane dehydration-enhanced esterification for biodiesel production from a potential feedstock of Firmiana platanifolia L.f. seed oil. Chem. Eng. Res. Des. 2020, 153, 1–7. [Google Scholar] [CrossRef]
- Sun, S.; Guo, J.; Duan, X. Biodiesel preparation from Phoenix tree seed oil using ethanol as acyl acceptor. Ind. Crop. Prod. 2019, 137, 270–275. [Google Scholar] [CrossRef]
- Sun, S.; Li, X. Physicochemical properties and fatty acid profile of Phoenix tree seed and its oil. J. Am. Oil Chem. Soc. 2016, 93, 1111–1114. [Google Scholar] [CrossRef]
- Zhi, X.; Gao, S.; Zou, J.; Ma, N.; Liu, T.; Wang, H.R.; Hu, Y.-Y.; Yao, Y.; Rui, W.H. Responses of the growth and nutrient stoichiometry in Ricinus communis seedlings on four soil types. J. Elem. 2022, 27, 223–238. [Google Scholar] [CrossRef]
- Dong, R.; Yang, S.; Wang, X.; Xie, L.; Ma, Y.; Wang, Y.; Zhang, L.; Zhang, M.; Qin, J. C:N:P stoichiometry in plant, soil and microbe in Sophora moorcroftiana shrubs across three sandy dune types in the middle reaches of the Yarlung Zangbo River. Front. Plant Sci. 2023, 13, 1060686. [Google Scholar] [CrossRef]
- Hu, L.; Ade, L.; Wu, X.; Zi, H.; Luo, X.; Wang, C. Changes in soil C:N:P stoichiometry and microbial structure along soil depth in two forest soils. Forests 2019, 10, 113. [Google Scholar] [CrossRef]
- Schulz, V.S.; Munz, S.; Stolzenburg, K.; Hartung, J.; Weisenburger, S.; Graeff-Honninger, S. Impact of different shading levels on growth, yield and quality of potato (Solanum tuberosum L.). Agronomy 2019, 9, 330. [Google Scholar] [CrossRef]
- Li, S.S.; Liu, C.; Tan, X.; Tan, B.; He, Y.X.; Li, N.W. Interactive effects of light and nitrogen on pakchoi (Brassica chinensis L.) growth and soil enzyme activity in an underground environment. Agronomy 2020, 10, 1772. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, W.; Ding, Y.; Zhang, J.; Cambula, E.D.; Weng, F.; Liu, Z.; Ding, C.; Tang, S.; Chen, L.; et al. Shading contributes to the reduction of stem mechanical strength by decreasing cell wall synthesis in japonica rice (Oryza sativa L.). Front. Plant Sci. 2017, 8, 881. [Google Scholar] [CrossRef]
- Quevedo-Rojas, A.; García-Núñez, C.; Jerez-Rico, M.; Jaimez, R.E.; Schwarzkopf, T. Leaf acclimation strategies to contrasting light conditions in saplings of different shade tolerance in a tropical cloud forest. Funct. Plant Biol. 2018, 45, 968–982. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Xiao, W.; Shi, Z.; Zeng, L.; Lei, L. Combined effects of drought and shading on growth and non-structural carbohydrates in Pinus massoniana Lamb. Seedlings. Forests 2020, 11, 18. [Google Scholar] [CrossRef]
- Yin, J.; Lin, F.; De Lombaerde, E.; Mao, Z.; Liu, S.; Ye, J.; Fang, S.; Wang, X. The effects of light, conspecific density and soil fungi on seedling growth of temperate tree species. For. Ecol. Manag. 2023, 529, 120683. [Google Scholar] [CrossRef]
- Dan, T.H.; Brix, H. Effects of soil type and water saturation on growth, nutrient and mineral content of the perennial forage shrub Sesbania sesban. Agrofor. Syst. 2017, 91, 173–184. [Google Scholar] [CrossRef]
- Lin, J.; Zhang, R.; Hu, Y.; Song, Y.; Hänninen, H.; Wu, J. Interactive effects of drought and shading on Torreya grandis seedlings: Physiological and growth responses. Trees-Struct. Funct. 2019, 33, 951–961. [Google Scholar] [CrossRef]
- Li, H.L.; Crabbe, M.J.C.; Xu, F.L.; Wang, W.L.; Ma, L.H.; Niu, R.L.; Gao, X.; Li, X.X.; Zhang, P.; Ma, X.; et al. Seasonal variations in carbon, nitrogen and phosphorus concentrations and C: N: P stoichiometry in different organs of a Larix principis-rupprechtii Mayr. plantation in the Qinling Mountains, China. PLoS ONE 2017, 12, e0185163. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Yang, K.; Chen, G.; Xian, J.; Chang, H. Divergent adaptations of leaf functional traits to light intensity across common urban plant species in Lanzhou, northwestern China. Front. Plant Sci. 2023, 14, 1000647. [Google Scholar] [CrossRef]
- Puglielli, G.; Laanisto, L.; Poorter, H.; Niinemets, U. Global patterns of biomass allocation in woody species with different tolerances of shade and drought: Evidence for multiple strategies. New Phytol. 2021, 229, 308–322. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, W.; Xu, M.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | pH | SOC (g·kg−1) | BD (g·cm−3) | TN (g·kg−1) | TP (g·kg−1) | TK (g·kg−1) |
|---|---|---|---|---|---|---|
| B1 | 4.34 ± 0.13 b | 42.53 ± 1.31 a | 1.24 ± 0.08 b | 2.33 ± 0.18 a | 0.24 ± 0.01 a | 4.99 ± 0.13 a |
| B2 | 5.14 ± 0.16 a | 16.64 ± 0.66 c | 1.30 ± 0.06 a | 1.14 ± 0.07 b | 0.07 ± 0.00 b | 3.15 ± 0.08 b |
| B3 | 4.79 ± 0.12 a | 33.50 ± 0.64 b | 1.32 ± 0.07 a | 1.89 ± 0.11 a | 0.08 ± 0.00 b | 2.21 ± 0.09 c |
| Treatment | SH (cm) | BLD (cm) | SLA (cm2·g−1) | R/T | Rob(g·cm−1) |
|---|---|---|---|---|---|
| A1B1 | 7.10 ± 0.39 C | 0.399 ± 0.025 Ab | 56.17 ± 3.07 Bb | 0.319 ± 0.016 A | 7.31 ± 0.32 Aa |
| A1B2 | 7.25 ± 0.32 C | 0.411 ± 0.019 Aa | 59.53 ± 3.80 Ba | 0.313 ± 0.017 A | 6.90 ± 0.17 Ab |
| A1B3 | 7.08 ± 0.15 C | 0.387 ± 0.023 Ac | 52.71 ± 3.71 Bb | 0.307 ± 0.119 A | 7.77 ± 0.38 Aa |
| A2B1 | 8.20 ± 0.18 B | 0.385 ± 0.021 Aa | 71.71 ± 3.66 Bb | 0.160 ± 0.014 B | 6.23 ± 0.49 Ba |
| A2B2 | 8.83 ± 0.21 B | 0.372 ± 0.014 Ab | 75.45 ± 2.64 Ba | 0.164 ± 0.008 B | 5.67 ± 0.36 Ba |
| A2B3 | 8.73 ± 0.26 B | 0.367 ± 0.018 Ac | 63.68 ± 3.94 Bb | 0.162 ± 0.113 B | 6.24 ± 0.16 Ba |
| A3B1 | 9.50 ± 0.19 A | 0.391 ± 0.022 Aa | 66.70 ± 1.94 Bb | 0.171 ± 0.016 B | 6.88 ± 0.24 Ba |
| A3B2 | 9.30 ± 0.17 B | 0.401 ± 0.019 Aa | 71.57 ± 3.53 Ba | 0.167 ± 0.020 B | 6.77 ± 0.14 Ba |
| A3B3 | 9.33 ± 0.22 AB | 0.389 ± 0.017 Aa | 63.79 ± 2.61 Bb | 0.160 ± 0.117 B | 6.92 ± 0.06 Ba |
| A4B1 | 10.40 ± 0.25 A | 0.277 ± 0.013 Ba | 168.05 ± 3.84 Aa | 0.131 ± 0.009 B | 1.95 ± 0.10 Ca |
| A4B2 | 10.58 ± 0.20 A | 0.263 ± 0.009 Ba | 154.48 ± 3.04 Ab | 0.125 ± 0.015 B | 1.75 ± 0.12 Ca |
| A4B3 | 10.03 ± 0.17 A | 0.269 ± 0.011 Ba | 151.12 ± 3.44 Ab | 0.120 ± 0.116 B | 1.83 ± 0.07 Ca |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhi, X.; Song, Y.; Yu, D.; Qian, W.; He, M.; Lin, X.; Zhang, D.; Gao, S. Early Growth Characterization and C:N:P Stoichiometry in Firmiana simplex Seedlings in Response to Shade and Soil Types. Forests 2023, 14, 1481. https://doi.org/10.3390/f14071481
Zhi X, Song Y, Yu D, Qian W, He M, Lin X, Zhang D, Gao S. Early Growth Characterization and C:N:P Stoichiometry in Firmiana simplex Seedlings in Response to Shade and Soil Types. Forests. 2023; 14(7):1481. https://doi.org/10.3390/f14071481
Chicago/Turabian StyleZhi, Ximin, Yi Song, Deshui Yu, Wenzhang Qian, Min He, Xi Lin, Danju Zhang, and Shun Gao. 2023. "Early Growth Characterization and C:N:P Stoichiometry in Firmiana simplex Seedlings in Response to Shade and Soil Types" Forests 14, no. 7: 1481. https://doi.org/10.3390/f14071481
APA StyleZhi, X., Song, Y., Yu, D., Qian, W., He, M., Lin, X., Zhang, D., & Gao, S. (2023). Early Growth Characterization and C:N:P Stoichiometry in Firmiana simplex Seedlings in Response to Shade and Soil Types. Forests, 14(7), 1481. https://doi.org/10.3390/f14071481