
Effects of Snow Cover on Carbon Dioxide Emissions and Their δ13C Values of Temperate Forest Soils with and without Litter



Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Collection of Soil Columns
2.2. Setup of Incubation Experiments
2.3. Measurements of CO2 Emissions from Soil Columns and Their δ13C Values
2.4. Statistical Analysis
3. Results
3.1. Changes in Soil Moisture at the Different Stages of the Incubation
3.2. Effect of Snow Cover and the Presence of Litter on CO2 Emissions from Soil Columns
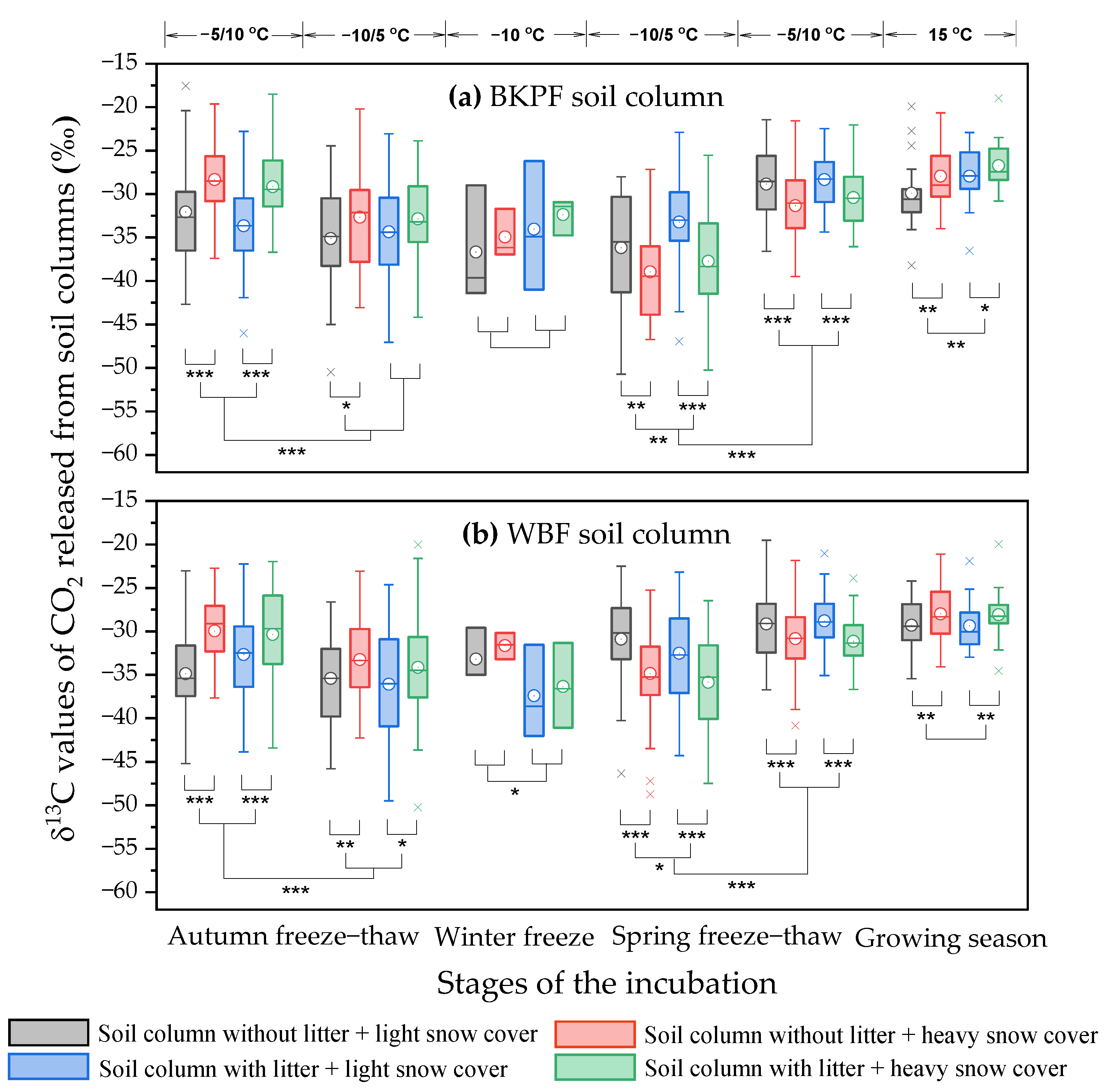
3.3. Effect of Snow Cover and the Presence of Litter on the δ13C Values of Soil-Respired CO2
3.4. Relationships among Soil CO2 Emissions, δ13C Values of Soil-Respired CO2, and Soil Moisture
4. Discussion
4.1. Effects of Snow Cover on Soil CO2 Emissions Vary with the Presence of Litter and Forest Stands
4.2. Changes in the δ13C Values of Soil-Respired CO2 and Influencing Mechanisms
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, T.; Barry, R.G.; Armstrong, R.L. Application of satellite remote sensing techniques to frozen ground studies. Polar Geogr. 2004, 28, 163–196. [Google Scholar] [CrossRef]
- Tarnocai, C.; Canadell, J.G.; Schuur, E.A.G.; Kuhry, P.; Mazhitova, G.; Zimov, S. Soil organic carbon pools in the northern circumpolar permafrost region. Glob. Biogeochem. Cycles 2009, 23, GB2023. [Google Scholar] [CrossRef]
- Brooks, P.D.; Grogan, P.; Templer, P.H.; Groffman, P.; Öquist, M.G.; Schimel, J. Carbon and nitrogen cycling in snow-covered environments. Geogr. Compass 2011, 5, 682–699. [Google Scholar] [CrossRef]
- Uchida, M.; Mo, W.; Nakatsubo, T.; Tsuchiya, Y.; Horikoshi, T.; Koizumi, H. Microbial activity and litter decomposition under snow cover in a cool-temperate broad-leaved deciduous forest. Agric. For. Meteorol. 2005, 134, 102–109. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Climate Change 2020: The Physical Science Basis; Cambridge University Press: New York, NY, USA, 2021. [Google Scholar]
- Brown, R.D.; Robinson, D.A. Northern Hemisphere spring snow cover variability and change over 1922–2010 including an assessment of uncertainty. Cryosphere 2011, 5, 219–229. [Google Scholar] [CrossRef]
- Kreyling, J.; Haei, M.; Laudon, H. Absence of snow cover reduces understory plant cover and alters plant community composition in boreal forests. Oecologia 2012, 168, 577–587. [Google Scholar] [CrossRef]
- Robroek, B.J.M.; Heijboer, A.; Jassey, V.E.J.; Hefting, M.M.; Rouwenhorst, T.G.; Buttler, A.; Bragazza, L. Snow cover manipulation effects on microbial community structure and soil chemistry in a mountain bog. Plant Soil 2013, 369, 151–164. [Google Scholar] [CrossRef]
- Wu, J.B.; Guan, D.X.; Wang, M.; Pei, T.F.; Han, S.J.; Jin, C.J. Year-round soil and ecosystem respiration in a temperate broad-leaved Korean pine forest. For. Ecol. Manag. 2006, 223, 35–44. [Google Scholar] [CrossRef]
- Liu, X.P.; Dong, W.X.; Wood, J.D.; Wang, Y.Y.; Li, X.X.; Zhang, Y.M.; Hu, C.S.; Gu, L.H. Aboveground and belowground contributions to ecosystem respiration in a temperate deciduous forest. Agric. For. Meteorol. 2022, 314, 108807. [Google Scholar] [CrossRef]
- Xu, X.K. Effect of freeze-thaw disturbance on soil C and N dynamics and GHG fluxes of East Asia forests: Review and future perspectives. Soil Sci. Plant Nutr. 2022, 68, 15–26. [Google Scholar] [CrossRef]
- Kalbitz, K.; Solinger, S.; Park, J.H.; Michalzik, B.; Matzner, E. Controls on the dynamics of dissolved organic matter in soils: A review. Soil Sci. 2000, 165, 277–304. [Google Scholar] [CrossRef]
- Wardle, D.A.; Walker, L.R.; Bardgett, R.D. Ecosystem properties and forest decline in contrasting long-term chronosequences. Science 2004, 305, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Ngao, J.; Epron, D.; Brechet, C.; Granier, A. Estimating the contribution of leaf litter decomposition to soil CO2 efflux in a beech forest using 13C-depleted litter. Glob. Chang. Biol. 2005, 11, 1768–1776. [Google Scholar] [CrossRef]
- Sakata, T.; Ishizuka, S.; Takahashi, M. Separation of soil respiration into CO2 emission sources using 13C natural abundance in a deciduous broad-leaved forest in Japan. Soil Sci. Plant Nutr. 2007, 53, 328–336. [Google Scholar] [CrossRef]
- Mauritz, M.; Celis, G.; Ebert, C.; Hutchings, J.; Ledman, J.; Natali, S.M.; Pegoraro, E.; Salmon, V.G.; Schadel, C.; Taylor, M. Using stable carbon isotopes of seasonal ecosystem respiration to determine permafrost carbon loss. J. Geophys. Res.-Biogeosci. 2019, 124, 46–60. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zou, J.L.; Meng, D.L.; Dang, S.N.; Zhou, J.H.; Osborne, B.; Ren, Y.Y.; Liang, T.; Yu, K.K. Effect of soil microorganisms and labile C availability on soil respiration in response to litter inputs in forest ecosystems: A meta-analysis. Ecol. Evol. 2020, 10, 13602–13612. [Google Scholar] [CrossRef]
- Kiikkila, O.; Kitunen, V.; Spetz, P.; Smolander, A. Characterization of dissolved organic matter in decomposing Norway spruce and silver birch litter. Eur. J. Soil Sci. 2012, 63, 476–486. [Google Scholar] [CrossRef]
- Wu, H.H.; Xu, X.K.; Cheng, W.G.; Fu, P.Q.; Li, F.Y. Antecedent soil moisture prior to freezing can affect quantity, composition and stability of soil dissolved organic matter during thaw. Sci. Rep. 2017, 7, 6380. [Google Scholar] [CrossRef] [PubMed]
- Hobara, S.; Osono, T.; Hirose, D.; Noro, K.; Hirota, M.; Benner, R. The roles of microorganisms in litter decomposition and soil formation. Biogeochemistry 2014, 118, 471–486. [Google Scholar] [CrossRef]
- Wu, H.H.; Xu, X.K.; Fu, P.Q.; Cheng, W.G.; Fu, C.S. Responses of soil WEOM quantity and quality to freeze-thaw and litter manipulation with contrasting soil water content: A laboratory experiment. Catena 2021, 198, 105058. [Google Scholar] [CrossRef]
- Tan, B.; Wu, F.Z.; Yang, W.Q.; He, X.H. Snow removal alters soil microbial biomass and enzyme activity in a Tibetan alpine forest. Appl. Soil Ecol. 2014, 76, 34–41. [Google Scholar] [CrossRef]
- Nakajima, M.; Cheng, W.G.; Tang, S.R.; Hori, Y.; Yaginuma, E.; Hattori, S.; Hanayama, S.; Tawaraya, K.; Xu, X.K. Modeling aerobic decomposition of rice straw during off-rice season in an Andisol paddy soil in a cold temperate region, Japan: Effects of soil temperature and moisture. Soil Sci. Plant Nutr. 2016, 62, 90–98. [Google Scholar] [CrossRef]
- Wu, H.H.; Xu, X.K.; Cheng, W.G.; Han, L. Dissolved organic matter and inorganic N jointly regulate greenhouse gases fluxes from forest soils with different moistures during a freeze-thaw period. Soil Sci. Plant Nutr. 2020, 66, 163–176. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, S.R.; Wang, J.X.; Sun, P.S.; Liu, W.G.; Hartley, D.S. Effects of seasonal snow on the growing season of temperate vegetation in China. Glob. Chang. Biol. 2013, 19, 2182–2195. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H. Impact of winter climate change on nitrogen biogeochemistry in forest ecosystems: A synthesis from Japanese case studies. Ecol. Indic. 2016, 65, 4–9. [Google Scholar] [CrossRef]
- Zhou, B.T.; Wang, Z.Y.; Shi, Y.; Xu, Y.; Han, Z.Y. Historical and future changes of snowfall events in China under warming background. J. Clim. 2018, 31, 5873–5889. [Google Scholar] [CrossRef]
- Wang, C.K.; Han, Y.; Chen, J.Q.; Wang, X.C.; Zhang, Q.Z.; Bond-Lamberty, B. Seasonality of Soil CO2 Efflux in a Temperate Forest: Biophysical Effects of Snowpack and Spring Freeze–Thaw Cycles. Agric. For. Meteorol. 2013, 177, 83–92. [Google Scholar] [CrossRef]
- Xu, X.K.; Duan, C.T.; Wu, H.H.; Li, T.S.; Cheng, W.G. Effect of intensity and duration of freezing on soil microbial biomass, extractable C and N pools, and N2O and CO2 emissions from forest soils in cold temperate region. Sci. China Earth Sci. 2016, 59, 156–169. [Google Scholar] [CrossRef]
- Schindlbacher, A.; Jandl, R.; Schindlbacher, S. Natural variations in snow cover do not affect the annual soil CO2 efflux from a mid-elevation temperate forest. Glob. Chang. Biol. 2014, 20, 622–632. [Google Scholar] [CrossRef]
- Gao, D.C.; Hagedorn, F.; Zhang, L.; Liu, J.; Qu, G.F.; Sun, J.F.; Peng, B.; Fan, Z.Z.; Zheng, J.Q.; Jiang, P.; et al. Small and transient response of winter soil respiration and microbial communities to altered snow depth in a mid-temperate forest. Appl. Soil Ecol. 2018, 130, 40–49. [Google Scholar] [CrossRef]
- Tang, S.R.; Cheng, W.G.; Hu, R.G.; Nakajima, M.; Guigue, J.; Kimani, S.M.; Sato, S.; Tawaraya, K.; Xu, X.K. Decomposition of soil organic carbon influenced by soil temperature and moisture in Andisol and Inceptisol paddy soils in a cold temperate region of Japan. J. Soils Sediments 2017, 17, 1843–1851. [Google Scholar] [CrossRef]
- Li, W.; Wu, J.; Bai, E.; Jin, C.; Wang, A.; Yuan, F.; Guan, D. Response of terrestrial carbon dynamics to snow cover change: A meta-analysis of experimental manipulation (II). Soil Biol. Biochem. 2016, 103, 388–393. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Hart, S.C. Consequences of manipulated snow cover on soil gaseous emission and N retention in the growing season: A meta-analysis. Ecosphere 2012, 3, 1–20. [Google Scholar] [CrossRef]
- Moyes, A.B.; Gaines, S.J.; Siegwolf, R.T.W.; Bowling, D.R. Diffusive fractionation complicates isotopic partitioning of autotrophic and heterotrophic sources of soil respiration. Plant Cell Environ. 2010, 33, 1804–1819. [Google Scholar] [CrossRef]
- Diao, H.Y.; Wang, A.Z.; Yuan, F.H.; Guan, D.X.; Wu, J.B. Autotrophic respiration modulates the carbon isotope composition of soil respiration in a mixed forest. Sci. Total Environ. 2022, 807, 150834. [Google Scholar] [CrossRef]
- Diao, H.Y.; Wang, A.Z.; Yuan, F.H.; Guan, D.X.; Dai, G.H.; Wu, J.B. Environmental effects on carbon isotope discrimination from assimilation to respiration in a coniferous and broad-leaved mixed forest of northeast China. Forests 2020, 11, 1156. [Google Scholar] [CrossRef]
- Zheng, J.P.; Guo, Z.L.; Xu, C.Y.; Fan, C.N. Seasonal dynamics of litter accumulation in major forest communities on the northern slope of Changbai Mountain, Northeast China. Acta Ecol. Sin. 2011, 31, 4299–4307. [Google Scholar]
- Ma, L.J.; Qin, D.H. Spatial-temporal characteristics of observed key parameters for snow cover in China during 1957–2009. J. Glaciol. Geocryol. 2012, 34, 1–11. [Google Scholar]
- Li, D.S.; Zheng, J.Q.; Wang, X.X.; Zheng, X.B.; Han, S.J. Effects of nitrogen addition and water manipulation on leaf litter decomposition. J. Beijing For. Univ. 2016, 38, 44–52. [Google Scholar]
- Yang, L.Y.; Li, W.H.; Wu, S.T. Fine root decomposition and nitrogen mineralization of the primitive Korean pine and broadleaved forests as well as its secondary forests in the Changbaishan Mountains, northeastern China. J. Beijing For. Univ. 2007, 29, 10–15. [Google Scholar]
- Wu, H.H.; Xu, X.K.; Duan, C.T.; Li, T.S.; Cheng, W.G. Synergistic effects of dissolved organic carbon and inorganic nitrogen on methane uptake in forest soils without and with freezing treatment. Sci. Rep. 2016, 6, 32555. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Liu, Q.J.; Xu, Z.Z.; Du, W.X.; Yu, J.; Meng, S.W.; Zhou, H.; Qin, L.H.; Shah, S. How can the shade intolerant Korean pine survive under dense deciduous canopy? For. Ecol. Manag. 2020, 457, 117735. [Google Scholar] [CrossRef]
- Hao, Z.Q.; Zhang, J.; Song, B.; Ye, J.; Li, B.H. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. For. Ecol. Manag. 2007, 252, 1–11. [Google Scholar] [CrossRef]
- Xu, X.K.; Han, L.; Wang, Y.S.; Inubushi, K. Influence of vegetation types and soil properties on microbial biomass carbon and metabolic quotients in temperate volcanic and tropical forest soils. Soil Sci. Plant Nutr. 2007, 53, 430–440. [Google Scholar] [CrossRef]
- Xu, X.K.; Yin, L.; Duan, C.T.; Jing, Y.S. Effect of N addition, moisture, and temperate on soil microbial respiration and microbial biomass in forest soil at different stages of litter decomposition. J. Soils Sediments 2016, 16, 1421–1439. [Google Scholar] [CrossRef]
- Keeling, C.D. The concentration and isotopic abundances of atmospheric carbon dioxide in rural areas. Geochim. Cosmochim. Acta 1958, 13, 322–334. [Google Scholar] [CrossRef]
- Keeling, C.D. The concentration and isotopic abundances of carbon dioxide in rural and marine air. Geochim. Cosmochim. Acta 1961, 24, 277–298. [Google Scholar] [CrossRef]
- Kammer, A.; Tuzson, B.; Emmenegger, L.; Knohl, A.; Mohn, J.; Hagedorn, F. Application of a quantum cascade laser-based spectrometer in a closed chamber system for real-time δ13C and δ18O measurements of soil-respired CO2. Agric. For. Meteorol. 2011, 151, 39–48. [Google Scholar] [CrossRef]
- Wen, X.F.; Meng, Y.; Zhang, X.Y.; Sun, X.M.; Lee, X. Evaluating calibration strategies for isotope ratio infrared spectroscopy for atmospheric 13CO2/12CO2 measurement. Atmos. Meas. Technol. 2013, 6, 1491–1501. [Google Scholar] [CrossRef]
- Chen, C.H.; Wei, J.; Wang, J.Y.; Guo, Q.J.; Wen, X.F. Disentangling diel variations in the carbon isotopic composition of soil CO2 efflux: Insights from a multi-chamber system and a non-steady model. J. Soil Sci. Plant Nutr. 2022, 22, 3096–3109. [Google Scholar] [CrossRef]
- Zhu, J.J.; Wu, Q.X.; Wu, F.Z.; Yue, K.; Ni, X.Y. Decline in carbon decomposition from litter after snow removal is driven by a delayed release of carbohydrates. Plant Soil 2022, 481, 83–95. [Google Scholar] [CrossRef]
- Groffman, P.M.; Driscoll, C.T.; Fahey, T.J.; Hardy, J.P.; Fitzhugh, R.D.; Tierney, G.L. Colder soils in a warmer world: A snow manipulation study in a northern hardwood forest ecosystem. Biogeochemistry 2001, 56, 135–150. [Google Scholar] [CrossRef]
- Brooks, P.; Williams, M.; Schmidt, S. Inorganic nitrogen and microbial biomass dynamics before and during spring snowmelt. Biogeochemistry 1998, 43, 1–15. [Google Scholar] [CrossRef]
- Lee, M.S.; Lee, J.S.; Koizumi, H. Temporal variation in CO2 efflux from soil and snow surfaces in a Japanese cedar (Cryptomeria japonica) Plantation, central Japan. Ecol. Res. 2008, 23, 777–785. [Google Scholar] [CrossRef]
- Gao, D.C.; Peng, B.; Fan, Z.Z.; Pei, G.T.; Bai, E. Different winter soil respiration between two mid-temperate plantation forests. For. Ecol. Manag. 2018, 409, 390–398. [Google Scholar] [CrossRef]
- Wu, Q.Q. Season-dependent effect of snow depth on soil microbial biomass and enzyme activity in a temperate forest in Northeast China. Catena 2020, 195, 104760. [Google Scholar] [CrossRef]
- Yohannes, Y.; Shibistova, O.; Abate, A.; Fetene, M.; Guggenberger, G. Soil CO2 efflux in an Afromontane forest of Ethiopia as driven seasonality and tree species. For. Ecol. Manag. 2011, 261, 1090–1098. [Google Scholar] [CrossRef]
- Philben, M.; Bowering, K.; Podrebarac, F.A.; Laganière, J.; Edwards, K.; Ziegler, S.E. Enrichment of 13C with depth in soil organic horizons is not explained by CO2 or DOC losses during decomposition. Geoderma 2022, 424, 116004. [Google Scholar] [CrossRef]
- Benner, R.; Fogel, M.L.; Sprague, E.K.; Hodson, R.E. Depletion of 13C in lignin and its implications for stable carbon isotope studies. Nature 1987, 329, 708–710. [Google Scholar] [CrossRef]
- Bowling, D.R.; Pataki, E.E.; Randerson, J.T. Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytol. 2008, 178, 24–40. [Google Scholar] [CrossRef]
- Kalbitz, K.; Glaser, B.; Bol, R. Clear-cutting of a Norway spruce stand: Implications for controls on the dynamics of dissolved organic matter in the forest floor. Eur. J. Soil Sci. 2004, 55, 401–413. [Google Scholar] [CrossRef]
- Xu, X.K.; Luo, X.B.; Jiang, S.H.; Xu, Z.J. Biodegradation of dissolved organic carbon in soil extracts and leachates from a temperate forest stand and its relationship to ultraviolet absorbance. Chin. Sci. Bull. 2012, 57, 912–920. [Google Scholar] [CrossRef]
- Bowling, D.R.; Massman, W.J.; Schaeffer, S.M.; Burns, S.P.; Monson, R.K.; Williams, M.W. Biological and physical influences on the carbon isotope content of CO2 in a subalpine forest snowpack, Niwot Ridge, Colorado. Biogeochemistry 2009, 95, 37–59. [Google Scholar] [CrossRef]
- Zobitz, J.M.; Keener, J.P.; Schnyder, H.; Bowling, D.R. Sensitivity analysis and quantification of uncertainty for isotopic mixing relationships in carbon cycle research. Agric. For. Meteorol. 2006, 136, 56–75. [Google Scholar] [CrossRef]
- Andrews, J.A.; Matamala, R.; Westover, K.M.; Schlesinger, W.H. Temperature effects on the diversity of soil heterotrophs and the δ13C of soil-respired CO2. Soil Biol. Biochem. 2000, 32, 699–706. [Google Scholar] [CrossRef]
- Monson, R.K.; Lipson, D.L.; Burns, S.P.; Turnipseed, A.A.; Delany, A.C.; Williams, M.W.; Schmidt, S.K. Winter forest soil respiration controlled by climate and microbial community composition. Nature 2006, 439, 711–714. [Google Scholar] [CrossRef]
- Aanderud, Z.T.; Jones, S.E.; Schoolmaster, D.R., Jr.; Fierer, N.; Lennon, J.T. Sensitivity of soil respiration and microbial communities to altered snowfall. Soil Biol. Biochem. 2013, 57, 217–227. [Google Scholar] [CrossRef]
- Dijkstra, P.; Ishizu, A.; Doucett, R.; Hart, S.C.; Schwartz, E.; Menyailo, O.V.; Hungate, B.A. 13C and 15N natural abundance of the soil microbial biomass. Soil Biol. Biochem. 2006, 38, 3257–3266. [Google Scholar] [CrossRef]
- Werth, M.; Kuzyakov, Y. 13C fractionation at the root–microorganisms–soil interface: A review and outlook for partitioning studies. Soil Biol. Biochem. 2010, 42, 1372–1384. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem. 2003, 35, 837–843. [Google Scholar] [CrossRef]
- Schadt, C.W.; Martin, A.P.; Lipson, D.A.; Schmidt, S.K. Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science 2003, 301, 1359–1361. [Google Scholar] [CrossRef]
- Buckeridge, K.M.; Grogan, P. Deepened snow alters soil microbial nutrient limitations in arctic birch hummock tundra. Appl. Soil Ecol. 2008, 39, 210–222. [Google Scholar] [CrossRef]
- Weintraub, M.N.; Scott-Denton, L.E.; Schmidt, S.K.; Monson, R.K. The effects of tree rhizodeposition on soil exoenzyme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem. Oecologia 2007, 154, 327–338. [Google Scholar] [CrossRef]
- Gauthier, A.; Amiotte-Suchet, P.; Nelson, P.N.; Léeêque, J.; Zeller, B.; Hénault, C. Dynamics of the water extractable organic carbon pool during mineralisation in soils from a Douglas fir plantation and an oak-beech forest—An incubation experiment. Plant Soil 2010, 330, 465–479. [Google Scholar] [CrossRef]
- Tang, S.R.; Yuan, P.; Tawaraya, K.; Tokida, T.; Fukuoka, M.; Yoshimoto, M.; Sakai, H.; Hasegawa, T.; Xu, X.K.; Cheng, W.G. Winter nocturnal warming affects the freeze-thaw frequency, soil aggregate distribution, the contents and decomposability of C and N in paddy fields. Sci. Total Environ. 2022, 802, 149870. [Google Scholar] [CrossRef]
- Böstrom, B.; Comstedt, D.; Ekblad, A. Can isotopic fractionation during respiration explain the 13C-enriched sporocarps of ectomycorrhizal and saprotrophic fungi? New Phytol. 2007, 177, 1012–1019. [Google Scholar] [CrossRef]
- Amundson, R.; Stern, L.; Baisden, T.; Wang, Y. The isotopic composition of soil and soil-respired CO2. Geoderma 1998, 82, 83–114. [Google Scholar] [CrossRef]
- Barbour, M.M.; Hunt, J.E.; Kodama, N.; Laubach, J.; McSeveny, T.M.; Rogers, G.N.D.; Tcherkez, G.; Wingate, L. Rapid changes in δ13C of ecosystem-respired CO2 after sunset are consistent with transient 13C enrichment of leaf respired CO2. New Phytol. 2011, 190, 990–1002. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Lv, S.J.; Lv, G.Y.; Xu, X.B.; Yao, H.Y.; Yu, Z.H.; Qiu, X.; Wang, Z.Y.; Wang, C.J. Effects of warming and precipitation on soil CO2 flux and its stable carbon isotope composition in the temperate desert steppe. Sustainability 2022, 14, 3351. [Google Scholar] [CrossRef]
- Phillips, C.L.; Nickerson, N.; Risk, D.; Kayler, Z.E.; Anderson, C.; Mix, A.; Bond, B.J. Soil moisture effects on the carbon isotope composition of soil respiration. Rapid Commun. Mass Spectrom. 2010, 24, 1271–1280. [Google Scholar] [CrossRef]
- Babur, E.; Dindaroglu, T.; Danish, S.; Haggblom, M.M.; Ozlu, E.; Gozukara, G.; Uslu, O.S. Spatial responses of soil carbon stocks, total nitrogen, and microbial indices to post-wildfire in the Mediterranean red pine forest. J. Environ. Manag. 2022, 320, 115939. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Levels of Snow Cover | Simulation Stages | ||
|---|---|---|---|
| Autumn Freeze–Thaw | Winter Freeze | Spring Freeze–Thaw | |
| Light snow cover | 35 g and 35 g on the first and 8th days of the incubation, respectively | 160 g on the 15th day of the incubation | 70 g on the 205th day of the incubation |
| Heavy snow cover | 150 g and 150 g on the first and 8th days of the incubation, respectively | 600 g on the 15th day of the incubation | 300 g on the 205th day of the incubation |
| Simulation Stage | Autumn Freeze–Thaw | Winter Freeze | Spring Freeze–Thaw | Growing Season | ||
|---|---|---|---|---|---|---|
| Duration (day) | 7 | 7 | 180 | 7 | 7 | 61 |
| Daily freeze–thaw alternation time (hour) | 12/12 | 24 (a) | 12/12 | 24 (a) | ||
| Temperature setting (°C) | −5/10 | −10/5 | −10 | −10/5 | −5/10 | 15 |
| Forest Type (FT) | Litter Cover (L) | Levels of Snow Cover (S) | Simulation Stage | ||||
|---|---|---|---|---|---|---|---|
| Autumn Freeze–Thaw | Winter Freeze | Spring Freeze–Thaw | Growing Season | Total | |||
| (g CO2-C m−2) | |||||||
| BKPF | Without | Light | 4.84 ± 0.11 | 8.43 ± 0.24 | 22.15 ± 1.55 | 128.67 ± 3.77 | 164.08 ± 5.21 |
| Heavy | 5.74 ± 0.26 | 10.44 ± 0.77 | 11.98 ± 0.49 | 139.46 ± 2.41 | 167.62 ± 2.30 | ||
| With | Light | 6.51 ± 0.31 | 16.02 ± 0.47 | 25.08 ± 0.30 | 161.92 ± 2.08 | 209.53 ± 2.34 | |
| Heavy | 8.58 ± 0.79 | 20.42 ± 1.68 | 17.13 ± 1.19 | 197.17 ± 13.47 | 243.30 ± 16.82 | ||
| WBF | Without | Light | 5.71 ± 0.11 | 15.03 ± 0.73 | 24.01 ± 1.51 | 154.06 ± 3.69 | 198.81 ± 5.59 |
| Heavy | 7.67 ± 0.14 | 19.73 ± 0.81 | 16.30 ± 0.47 | 182.48 ± 2.67 | 226.17 ± 3.69 | ||
| With | Light | 6.55 ± 0.65 | 27.04 ± 1.65 | 23.99 ± 2.05 | 157.92 ± 9.07 | 215.51 ± 12.40 | |
| Heavy | 8.20 ± 0.54 | 30.57 ± 1.77 | 17.83 ± 0.39 | 203.71 ± 3.14 | 260.31 ± 5.76 | ||
| Three−way factorial ANOVA with FT, L, and S as fixed factors (p-value) | |||||||
| FT | 0.338 | <0.001 | 0.396 | 0.064 | 0.025 | ||
| L | 0.032 | <0.001 | 0.167 | 0.005 | 0.002 | ||
| S | 0.018 | 0.040 | <0.001 | 0.004 | 0.033 | ||
| FT × L | 0.227 | 0.434 | 0.337 | 0.084 | 0.154 | ||
| FT × S | 0.801 | 0.786 | 0.531 | 0.442 | 0.469 | ||
| L × S | 0.733 | 0.856 | 0.576 | 0.259 | 0.326 | ||
| FT × L × S | 0.564 | 0.594 | 0.922 | 0.845 | 0.789 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Wu, H.; Yue, J.; Tang, S.; Cheng, W. Effects of Snow Cover on Carbon Dioxide Emissions and Their δ13C Values of Temperate Forest Soils with and without Litter. Forests 2023, 14, 1384. https://doi.org/10.3390/f14071384
Xu X, Wu H, Yue J, Tang S, Cheng W. Effects of Snow Cover on Carbon Dioxide Emissions and Their δ13C Values of Temperate Forest Soils with and without Litter. Forests. 2023; 14(7):1384. https://doi.org/10.3390/f14071384
Chicago/Turabian StyleXu, Xingkai, Haohao Wu, Jin Yue, Shuirong Tang, and Weiguo Cheng. 2023. "Effects of Snow Cover on Carbon Dioxide Emissions and Their δ13C Values of Temperate Forest Soils with and without Litter" Forests 14, no. 7: 1384. https://doi.org/10.3390/f14071384
APA StyleXu, X., Wu, H., Yue, J., Tang, S., & Cheng, W. (2023). Effects of Snow Cover on Carbon Dioxide Emissions and Their δ13C Values of Temperate Forest Soils with and without Litter. Forests, 14(7), 1384. https://doi.org/10.3390/f14071384