
Acacia Density, Edaphic, and Climatic Factors Shape Plant Assemblages in Regrowth Montane Forests in Southeastern Australia

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Site Selection
2.3. Field Data Collection
2.4. Climate, Landform, and Soil
2.5. Statistical Analysis
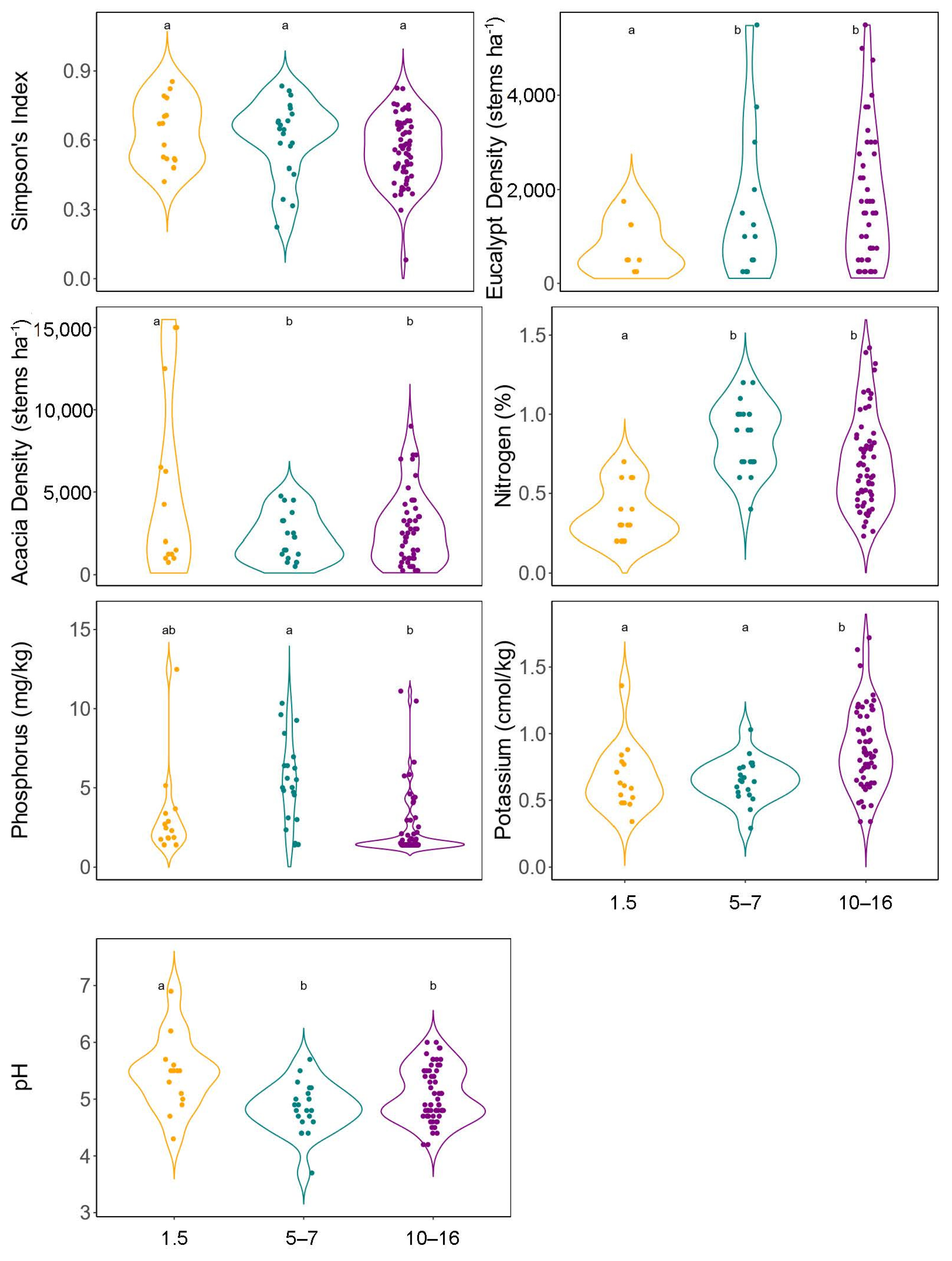
3. Results
3.1. Plant Community Composition and Response to Environmental Predictors
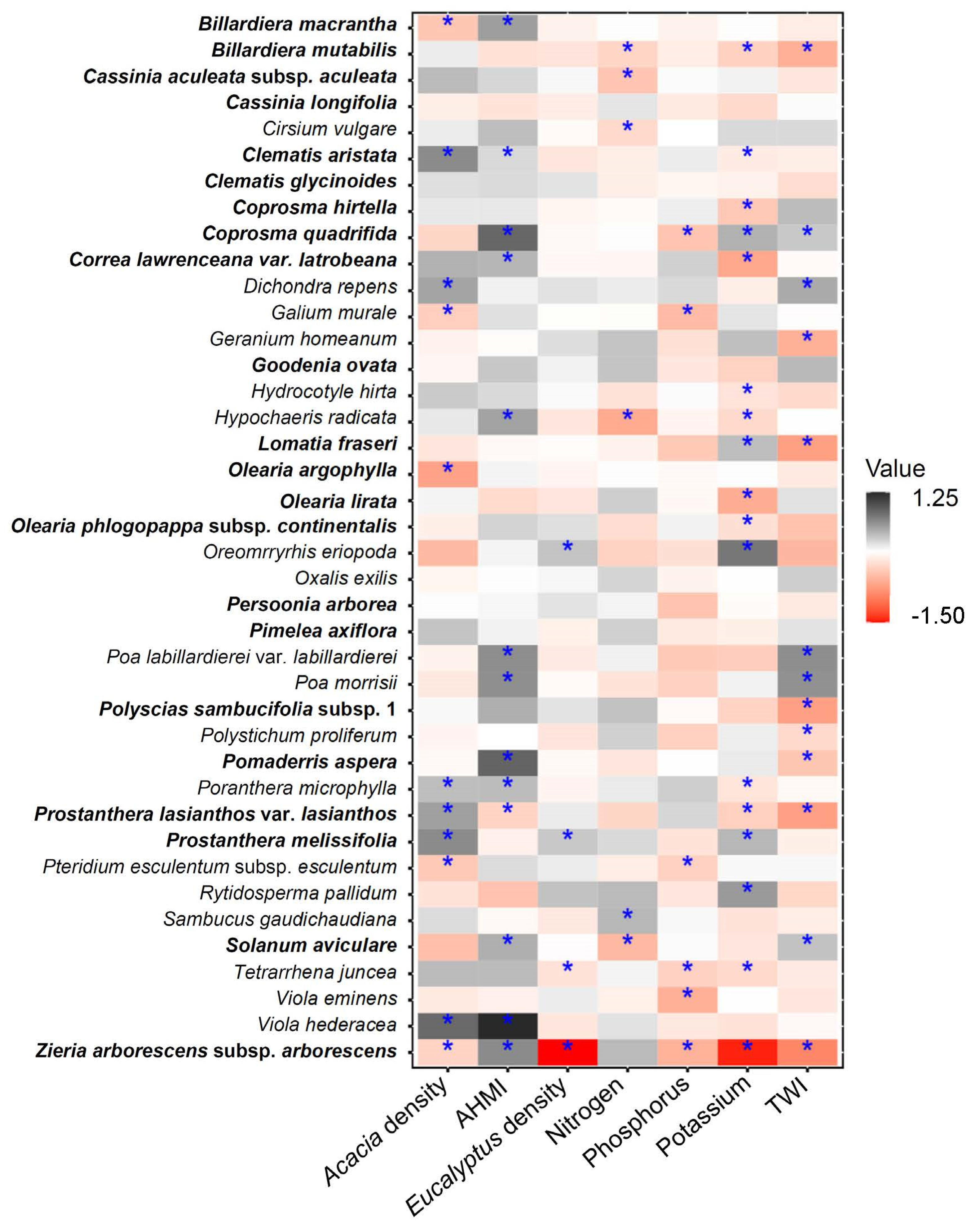
3.2. Species-Specific Responses to Environmental Predictors
4. Discussion
4.1. Canopy Structure and Density in Mediating Resource Availability
4.2. Species Responses
4.3. Effect of Soil Nutrients on Plant Community Composition
4.4. Effect of Moisture Variables on the Plant Community Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, G.E. Concluding remarks in Populations studies: Animal Ecology and Demography. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor: New York, NY, USA, 1957; pp. 415–427. [Google Scholar]
- Austin, M.P. A silent clash of paradigms: Some inconsistencies in community ecology. Oikos 1999, 86, 170–178. [Google Scholar] [CrossRef]
- Eilts, J.A.; Mittelbach, G.G.; Reynolds, H.L.; Gross, K.L. Resource heterogeneity, soil fertility, and species diversity: Effects of clonal species on plant communities. Am. Nat. 2011, 177, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Forrester, D.I. The spatial and temporal dynamics of species interactions in mixed-species forests: From pattern to process. For. Ecol. Manag. 2014, 312, 282–292. [Google Scholar] [CrossRef]
- Gosper, C.R.; Yates, C.J.; Prober, S.M. Floristic diversity in fire-sensitive eucalypt woodlands shows a “U”-shaped relationship with time since fire. J. Appl. Ecol. 2013, 50, 1187–1196. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.; Thomas, S.C.; Shahi, C. Linking resource availability and heterogeneity to understorey species diversity through succession in boreal forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Su, X.; Wang, M.; Huang, Z.; Fu, S.; Chen, H.Y. Forest understorey vegetation: Colonization and the Availability and Heterogeneity of resources. Forests 2019, 10, 944. [Google Scholar] [CrossRef]
- Reich, P.B.; Frelich, L.E.; Voldseth, R.A.; Bakken, P.; Adair, E.C. Understorey diversity in southern boreal forests is regulated by productivity and its indirect impacts on resource availability and heterogeneity. J. Ecol. 2012, 100, 539–545. [Google Scholar] [CrossRef]
- Veldman, J.W.; Brudvig, L.A.; Damschen, E.I.; Orrock, J.L.; Mattingly, W.B.; Walker, J.L. Fire frequency, agricultural history and the multivariate control of pine savanna understorey plant diversity. J. Veg. Sci. 2014, 25, 1438–1449. [Google Scholar] [CrossRef]
- Siefert, A.; Ravenscroft, C.; Althoff, D.; Alvarez-Yépiz, J.C.; Carter, B.E.; Glennon, K.L.; Heberling, J.M.; Jo, I.S.; Pontes, A.; Sauer, A.; et al. Scale dependence of vegetation-environment relationships: A meta-analysis of multivariate data. J. Veg. Sci. 2012, 23, 942–951. [Google Scholar] [CrossRef]
- Chick, M.P.; Nitschke, C.R.; Cohn, J.S.; Penman, T.D.; York, A. Factors influencing above-ground and soil seed bank vegetation diversity at different scales in a quasi-Mediterranean ecosystem. J. Veg. Sci. 2018, 29, 684–694. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology 2010, 91, 1931–1938. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.K.; Waeber, P.O.; Simard, S.W.; Innes, J.L.; Nitschke, C.R. Multiple factors influence plant richness and diversity in the cold and dry boreal forest of southwest Yukon, Canada. Plant. Ecol. 2016, 217, 505–519. [Google Scholar] [CrossRef]
- Barbier, S.; Gosselin, F.; Balandier, P. Influence of tree species on understory vegetation diversity and mechanisms involved—A critical review for temperate and boreal forests. For. Ecol. Manag. 2008, 254, 1–15. [Google Scholar] [CrossRef]
- Kasel, S.; Bennett, L.T.; Aponte, C.; Fedrigo, M.; Nitschke, C.R. Environmental heterogeneity promotes floristic turnover in temperate forests of south-eastern Australia more than dispersal limitation and disturbance. Landsc. Ecol. 2017, 32, 1613–1629. [Google Scholar] [CrossRef]
- Harvey, B.J.; Holzman, B.A. Divergent successional pathways of stand development following fire in a California closed-cone pine forest. J. Veg. Sci. 2014, 25, 88–99. [Google Scholar] [CrossRef]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Clements, F.E. Plant Succession: An Analysis of the Development of Vegetation; Carnegie Institution of Washington: Washington, DC, USA, 1916. [Google Scholar]
- Egler, F.E. Vegetation science concepts I. Initial floristic composition, a factor in old-field vegetation development with 2 figs. Plant. Ecol. 1954, 4, 412–417. [Google Scholar] [CrossRef]
- Cole, D.; Rapp, M. Elemental cycling in forest ecosystems. In Dynamic Properties of Forest Ecosystems; Reichle, D.E., Ed.; International Biological Programme; Cambridge University Press: Cambridge, UK, 1981; pp. 341–409. [Google Scholar]
- Wilson, S.D.; Tilman, D. Plant competition and resource availability in response to disturbance and fertilization. Ecology 1993, 74, 599–611. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J. A Review of Processes Behind Diversity-Productivity Relationships in Forests. Curr. Rep. 2016, 2, 45–61. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef]
- Bartels, S.F.; Chen, H.Y. Interactions between overstorey and understorey vegetation along an overstorey compositional gradient. J. Veg. Sci. 2013, 24, 543–552. [Google Scholar] [CrossRef]
- Bartemucci, P.; Messier, C.; Canham, C.D. Overstory influences on light attenuation patterns and understory plant community diversity and composition in southern boreal forests of Quebec. Can. J. For. Res. 2006, 36, 2065–2079. [Google Scholar] [CrossRef]
- Bratton, S.P. Resource division in an understory herb community: Responses to temporal and microtopographic gradients. Am. Nat. 1976, 110, 679–693. [Google Scholar] [CrossRef]
- Ough, K. Regeneration of Wet Forest flora a decade after clear-felling or wildfire-is there a difference? Aust. J. Bot. 2001, 49, 645–664. [Google Scholar] [CrossRef]
- Blair, D.P.; McBurney, L.M.; Blanchard, W.; Banks, S.C.; Lindenmayer, D.B. Disturbance gradient shows logging affects plant functional groups more than fire. Ecol. Appl. 2016, 26, 2280–2301. [Google Scholar] [CrossRef]
- Bowd, E.J.; Lindenmayer, D.B.; Banks, S.C.; Blair, D.P. Logging and fire regimes alter plant communities. Ecol. Appl. 2018, 28, 826–841. [Google Scholar] [CrossRef]
- Trouvé, R.; Sherriff, R.M.; Holt, L.M.; Baker, P.J. Differing regeneration patterns after catastrophic fire and clearfelling: Implications for future stand dynamics and forest management. For. Ecol. Manag. 2021, 498, 119555. [Google Scholar] [CrossRef]
- Kasel, S.; Nitschke, C.R.; Baker, S.C.; Pryde, E.C. Concurrent assessment of functional types in extant vegetation and soil seed banks informs environmental constraints and mechanisms of plant community turnover in temperate forests of south-eastern Australia. For. Ecol. Manag. 2022, 519, 14. [Google Scholar] [CrossRef]
- Pulsford, S.A.; Lindenmayer, D.B.; Driscoll, D.A. A succession of theories: Purging redundancy from disturbance theory. Biol. Rev. 2016, 91, 148–167. [Google Scholar] [CrossRef]
- Attiwill, P.M. Ecological disturbance and the conservative management of eucalypt forests in Australia. For. Ecol. Manag. 1994, 63, 301–346. [Google Scholar] [CrossRef]
- ABARES. Australia’s State of the Forest Report 2018; ABARES: Canberra, ACT, Australia, 2018. [Google Scholar]
- Stewart, S.B.; Nitschke, C.R. Improving temperature interpolation using MODIS LST and local topography: A comparison of methods in south east Australia. Int. J. Climatol. 2017, 37, 3098–3110. [Google Scholar] [CrossRef]
- Keenan, R.J.; Nitschke, C. Forest management options for adaptation to climate change: A case study of tall, wet eucalypt forests in Victoria’s Central Highlands region. Aust. For. 2016, 79, 96–107. [Google Scholar] [CrossRef]
- Booth, T.H.; Pryor, L.D. Climatic requirements of some commercially important eucalypt species. For. Ecol. Manag. 1991, 43, 47–60. [Google Scholar] [CrossRef]
- Adams, M.A.; Attiwill, P.M. Nutrient cycling and nitrogen mineralization in eucalypt forests of south-eastern Australia. Plant. Soil. 1986, 92, 341–362. [Google Scholar] [CrossRef]
- Ashton, D.H. Studies on the Autecology of Eucalyptus regnans F.v. M. Ph.D. Thesis, The University of Melbourne, Melbourne, VIC, Australia, 1956. [Google Scholar]
- Squire, R.; Geary, P.; Lutze, M. The East Gippsland Silvicultural Systems Project. I: The establishment of the project in lowland forest. Aust. For. 2006, 69, 167–181. [Google Scholar] [CrossRef]
- Dignan, P. Wood Production in Mountain Ash Forests: Implications of Alternative Systems for Harvesting Operations; VSP Technical Report. No. 22; Department of Conservation and Natural Resources: East Melbourne, VIC, Australia, 1993. [Google Scholar]
- Squire, R.O.; Campbell, R.G.; Wareing, K.J.; Featherston, G.R. The mountain ash forests of Victoria: Ecology, silviculture and management for wood production. In Forest Management in Australia, Proceedings of the Conference of the Institute of Foresters of Australia, Perth, WA, Australia, 18–22 September 1987; McKinnell, F.H., Hopkins, E.R., Fox, J.E.P., Eds.; Surrey Beatty & Sons: Chipping Norton, NSW, Australia, 1991; pp. 36–57. [Google Scholar]
- Keenan, R.J.; Kimmins, J.P. The ecological effects of clear-cutting. Environ. Rev. 1993, 1, 121–144. [Google Scholar] [CrossRef]
- Fedrigo, M.; Stewart, S.B.; Roxburgh, S.H.; Kasel, S.; Bennett, L.T.; Vickers, H.; Nitschke, C.R. Predictive Ecosystem Mapping of South-Eastern Australian Temperate Forests Using Lidar-Derived Structural Profiles and Species Distribution Models. Remote. Sens. 2019, 11, 93. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Yanchuk, A.; O’neill, G.; Aitken, S.N. Use of response functions in selecting lodgepole pine populations for future climates. Glob. Change Biol. 2006, 12, 2404–2416. [Google Scholar] [CrossRef]
- Raduła, M.W.; Szymura, T.H.; Szymura, M. Topographic wetness index explains soil moisture better than bioindication with Ellenberg’s indicator values. Ecol. Indic. 2018, 85, 172–179. [Google Scholar] [CrossRef]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods: Australasia; CSIRO Publishing: Collingwood, VIC, Australia, 2010. [Google Scholar]
- Rayment, G.E.; Higginson, F.R. Australian Laboratory Handbook of Soil and Water Chemical Methods; Inkata Press: Port Melbourne, VIC, Australia, 1992. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. mvabund—An R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Hui, F.K.C. boral—Bayesian Ordination and Regression Analysis of Multivariate Abundance Data in r. Methods Ecol. Evol. 2016, 7, 744–750. [Google Scholar] [CrossRef]
- Björk, J.R.; Hui, F.K.C.; O’Hara, R.B.; Montoya, J.M. Uncovering the drivers of host-associated microbiota with joint species distribution modelling. Mol. Ecol. 2018, 27, 2714–2724. [Google Scholar] [CrossRef] [PubMed]
- Niku, J.; Hui, F.K.C.; Taskinen, S.; Warton, D.I.; Goslee, S. gllvm: Fast analysis of multivariate abundance data with generalized linear latent variable models in R. Methods Ecol. Evol. 2019, 10, 2173–2182. [Google Scholar] [CrossRef]
- Warton, D.I.; Blanchet, F.G.; O’Hara, R.B.; Ovaskainen, O.; Taskinen, S.; Walker, S.C.; Hui, F.K.C. So Many Variables: Joint Modeling in Community Ecology. Trends Ecol. Evol. 2015, 30, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Ovaskainen, O.; Tikhonov, G.; Norberg, A.; Guillaume Blanchet, F.; Duan, L.; Dunson, D.; Roslin, T.; Abrego, N. How to make more out of community data? A conceptual framework and its implementation as models and software. Ecol. Lett. 2017, 20, 561–576. [Google Scholar] [CrossRef]
- Di Stefano, J. How Much Power Is Enough? Against the Development of an Arbitrary Convention for Statistical Power Calculations. Funct. Ecol. 2003, 17, 707–709. [Google Scholar] [CrossRef]
- Watanabe, S.; Opper, M. Asymptotic equivalence of Bayes cross validation and widely applicable information criterion in singular learning theory. J. Mach. Learn. Res. 2010, 11, 3571–3594. [Google Scholar]
- Singh, A.; Wagner, B.; Kasel, S.; Baker, P.J.; Nitschke, C.R. Canopy Composition and Spatial Configuration Influences Beta Diversity in Temperate Regrowth Forests of Southeastern Australia. Drones 2023, 7, 155. [Google Scholar] [CrossRef]
- Fairman, T.A.; Nitschke, C.R.; Bennett, L.T. Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. Int. J. Wildland Fire 2016, 25, 831–848. [Google Scholar] [CrossRef]
- Bowd, E.J.; McBurney, L.; Lindenmayer, D.B. The characteristics of regeneration failure and their potential to shift wet temperate forests into alternate stable states. For. Ecol. Manag. 2023, 529, 120673. [Google Scholar] [CrossRef]
- Anderson, M.C. The geometry of leaf distribution in some south-eastern Australian forests. Agric. Meteorol. 1981, 25, 195–206. [Google Scholar] [CrossRef]
- King, D.A. The functional significance of leaf angle in Eucalyptus. Aust. J. Bot. 1997, 45, 619–639. [Google Scholar] [CrossRef]
- Bauhus, J.; Van Winden, A.P.; Nicotra, A.B. Aboveground interactions and productivity in mixed-species plantations of Acacia mearnsii and Eucalyptus globulus. Can. J. For. Res. 2004, 34, 686–694. [Google Scholar] [CrossRef]
- Thomas, S.C.; Halpern, C.B.; Falk, D.A.; Liguori, D.A.; Austin, K.A. Plant diversity in managed forests: Understory responses to thinning and fertilization. Ecol. Appl. 1999, 9, 864–879. [Google Scholar] [CrossRef]
- Baker, S.C.; Kasel, S.; van Galen, L.G.; Jordan, G.J.; Nitschke, C.R.; Pryde, E.C. Identifying regrowth forests with advanced mature forest values. For. Ecol. Manag. 2019, 433, 73–84. [Google Scholar] [CrossRef]
- May, B. Silver Wattle (Acacia dealbata): Its Role in the Ecology of the Mountain Ash Forest and the Effect of Alternative Silvicultural Systems on Its Regeneration. Ph.D. Thesis, The University of Melbourne, Melbourne, VIC, Australia, 1999. [Google Scholar]
- Trouvé, R.; Nitschke, C.R.; Andrieux, L.; Willersdorf, T.; Robinson, A.P.; Baker, P.J. Competition drives the decline of a dominant midstorey tree species. Habitat implications for an endangered marsupial. For. Ecol. Manag. 2019, 447, 26–34. [Google Scholar] [CrossRef]
- Ashton, D.H. The root and shoot development of Eucalyptus regnans F. Muell. Aust. J. Bot. 1975, 23, 867–887. [Google Scholar] [CrossRef]
- Ashton, D.H. The development of even-aged stands of Eucalyptus regnans F. Muell. in central Victoria. Aust. J. Bot. 1976, 24, 397–414. [Google Scholar] [CrossRef]
- Ashton, D.H. The big ash forest, Wallaby Creek, Victoria—Changes during one lifetime. Aust. J. Bot. 2000, 48, 1–26. [Google Scholar] [CrossRef]
- Forrester, D.I. Growth responses to thinning, pruning and fertiliser application in Eucalyptus plantations: A review of their production ecology and interactions. For. Ecol. Manag. 2013, 310, 336–347. [Google Scholar] [CrossRef]
- White, D.J.; Vesk, P.A. Fire and legacy effects of logging on understorey assemblages in wet-sclerophyll forests. Aust. J. Bot. 2019, 67, 341–357. [Google Scholar] [CrossRef]
- Cadiz, G.O.; Cawson, J.G.; Penman, T.D.; York, A.; Duff, T.J. Environmental factors associated with the abundance of forest wiregrass (Tetrarrhena juncea), a flammable understorey grass in productive forests. Aust. J. Bot. 2000, 68, 37–48. [Google Scholar] [CrossRef]
- Penman, T.D.; Binns, D.L.; Shiels, R.J.; Allen, R.M.; Kavanagh, R.P. Changes in understorey plant species richness following logging and prescribed burning in shrubby dry sclerophyll forests of south-eastern Australia. Austral Ecol. 2008, 33, 197–210. [Google Scholar] [CrossRef]
- Ashwell, D. The Importance of Tetrarrhena juncea in the Ecology of Eucalyptus regnans Stands. Master’s Thesis, The University of Melbourne, Parkville, VIC, Australia, 1985. [Google Scholar]
- Cadiz, G.O.; Cawson, J.G.; Duff, T.J.; Penman, T.D.; York, A.; Farrell, C. Independent effects of drought and shade on growth, biomass allocation and leaf morphology of a flammable perennial grass Tetrarrhena juncea R.Br. Plant. Ecol. 2021, 222, 877–895. [Google Scholar] [CrossRef]
- Younis, S.; Kasel, S. Do Fire Cues Enhance Germination of Soil Seed Stores across an Ecotone of Wet Eucalypt Forest to Cool Temperate Rainforest in the Central Highlands of South-Eastern Australia? Fire 2023, 6, 138. [Google Scholar] [CrossRef]
- Wang, L. The soil seed bank and understorey regeneration in Eucalyptus regnans forest, Victoria. Aust. J. Ecol. 1997, 22, 404–411. [Google Scholar] [CrossRef]
- Ashton, D. The seasonal growth of Eucalyptus regnans F. Muell. Aust. J. Bot. 1975, 23, 239–252. [Google Scholar] [CrossRef]
- Vickers, H.; Kasel, S.; Duff, T.; Nitschke, C. Recruitment and growth dynamics of a temperate forest understorey species following wildfire in southeast Australia. Dendrochronologia 2021, 67, 125829. [Google Scholar] [CrossRef]
- Murphy, A.; Ough, K. Regenerative strategies of understorey flora following clearfell logging in the Central Highlands, Victoria. Aust. For. 1997, 60, 90–98. [Google Scholar] [CrossRef]
- Ough, K.; Murphy, A. Decline in tree fern abundance after clearfell harvesting. For. Ecol. Manag. 2004, 199, 153–163. [Google Scholar] [CrossRef]
- Rab, M.A. Soil physical and hydrological properties following logging and slash burning in the Eucalyptus regnans forest of southeastern Australia. For. Ecol. Manag. 1996, 84, 159–176. [Google Scholar] [CrossRef]
- Weston, P.; Cameron, D. Persoonia arborea . In The IUCN Red List of Threatened Species 2020; IUCN: Gland, Switzerland, 2020; pp. 1–8. [Google Scholar]
- Gullan, P. A Rare Plant That Is Locally Abundant. Available online: http://www.viridans.com/RAREPL/locallyabundant.htm (accessed on 2 April 2023).
- Emery, N.J.; Offord, C.A. Managing Persoonia (Proteaceae) species in the landscape through a better understanding of their seed biology and ecology. Cunninghamia J. Plant. Ecol. East Aust. 2018, 18, 89–107. [Google Scholar]
- Adams, M.; Attiwill, P.; Polglase, P. Availability of nitrogen and phosphorus in forest soils in northeastern Tasmania. Biol. Fertil. Soils 1989, 8, 212–218. [Google Scholar] [CrossRef]
- Beadle, N. Soil phosphate and the delimitation of plant communities in eastern Australia. Ecology 1954, 35, 370–375. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Turner, M.G.; Mack, M.C.; Chapin, F.S. Postfire Soil N Cycling in Northern Conifer Forests Affected by Severe, Stand-Replacing Wildfires. Ecosystems 2005, 8, 163–181. [Google Scholar] [CrossRef]
- Wan, S.; Hui, D.; Luo, Y. Fire effects on nitrogen pools and dynamics in terrestrial ecosystems: A meta-analysis. Ecol. Appl. 2001, 11, 1349–1365. [Google Scholar] [CrossRef]
- Weston, C.J.; Attiwill, P.M. Effects of fire and harvesting on nitrogen transformations and ionic mobility in soils of Eucalyptus regnans forests of south-eastern Australia. Oecologia 1990, 83, 20–26. [Google Scholar] [CrossRef]
- Kreft, H.; Jetz, W. Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 5925–5930. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.J.; Audino, L.D.; Whitacre, J.; Eck, J.L.; Wenzel, J.W.; Queenborough, S.A.; Comita, L.S. Species associations structured by environment and land-use history promote beta-diversity in a temperate forest. Ecology 2015, 96, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Plue, J.; De Frenne, P.; Acharya, K.; Brunet, J.; Chabrerie, O.; Decocq, G.; Diekmann, M.; Graae, B.J.; Heinken, T.; Hermy, M.; et al. Where does the community start, and where does it end? Including the seed bank to reassess forest herb layer responses to the environment. J. Veg. Sci. 2017, 28, 424–435. [Google Scholar] [CrossRef]
- Baker, P.J.; Nitschke, C.R.; Trouve, R.; Robinson, A.P. Forest stand dynamics drive a conservation conundrum for the critically endangered Leadbeater’s Possum. In Forests as Complex Social and Ecological Systems: A Festschrift for Chadwick D. Oliver; Baker, P.J., Larsen, D.R., Saxena, A., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 93–113. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Predictors | Wald Statistic | p Value |
|---|---|---|
| Annual heat moisture index | 231.91 | 0.001 |
| Topographic wetness index | 121.44 | 0.001 |
| Phosphorus | 104.28 | 0.001 |
| Acacia density | 96.28 | 0.001 |
| Potassium | 78.71 | 0.008 |
| Nitrogen | 72.02 | 0.014 |
| Eucalyptus density | 56.96 | 0.230 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, A.; Kasel, S.; Hui, F.K.C.; Trouvé, R.; Baker, P.J.; Nitschke, C.R. Acacia Density, Edaphic, and Climatic Factors Shape Plant Assemblages in Regrowth Montane Forests in Southeastern Australia. Forests 2023, 14, 1166. https://doi.org/10.3390/f14061166
Singh A, Kasel S, Hui FKC, Trouvé R, Baker PJ, Nitschke CR. Acacia Density, Edaphic, and Climatic Factors Shape Plant Assemblages in Regrowth Montane Forests in Southeastern Australia. Forests. 2023; 14(6):1166. https://doi.org/10.3390/f14061166
Chicago/Turabian StyleSingh, Anu, Sabine Kasel, Francis K. C. Hui, Raphaël Trouvé, Patrick J. Baker, and Craig R. Nitschke. 2023. "Acacia Density, Edaphic, and Climatic Factors Shape Plant Assemblages in Regrowth Montane Forests in Southeastern Australia" Forests 14, no. 6: 1166. https://doi.org/10.3390/f14061166
APA StyleSingh, A., Kasel, S., Hui, F. K. C., Trouvé, R., Baker, P. J., & Nitschke, C. R. (2023). Acacia Density, Edaphic, and Climatic Factors Shape Plant Assemblages in Regrowth Montane Forests in Southeastern Australia. Forests, 14(6), 1166. https://doi.org/10.3390/f14061166