
Soil Depth Can Modify the Contribution of Root System Architecture to the Root Decomposition Rate

Abstract
1. Introduction
2. Materials and Methods
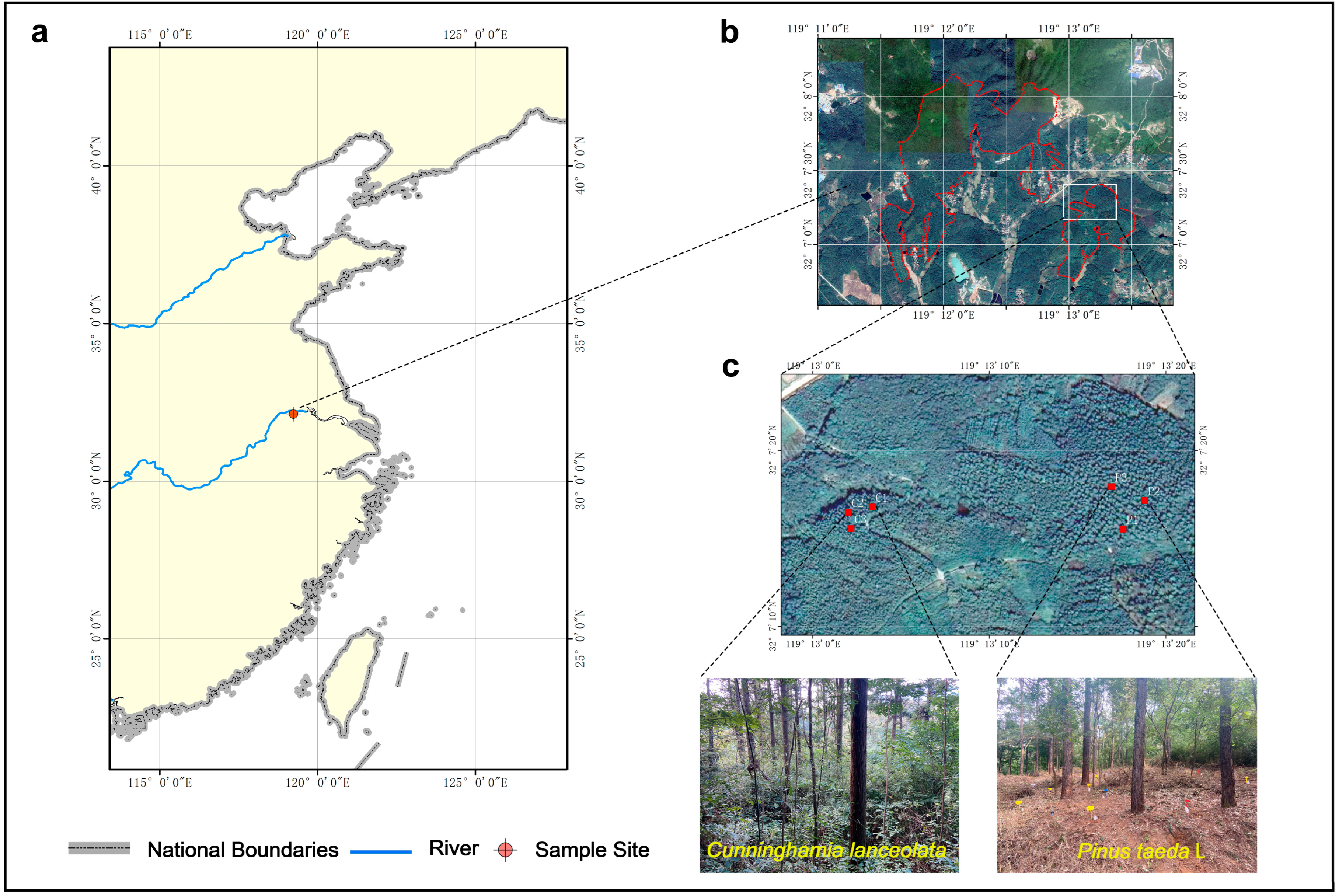
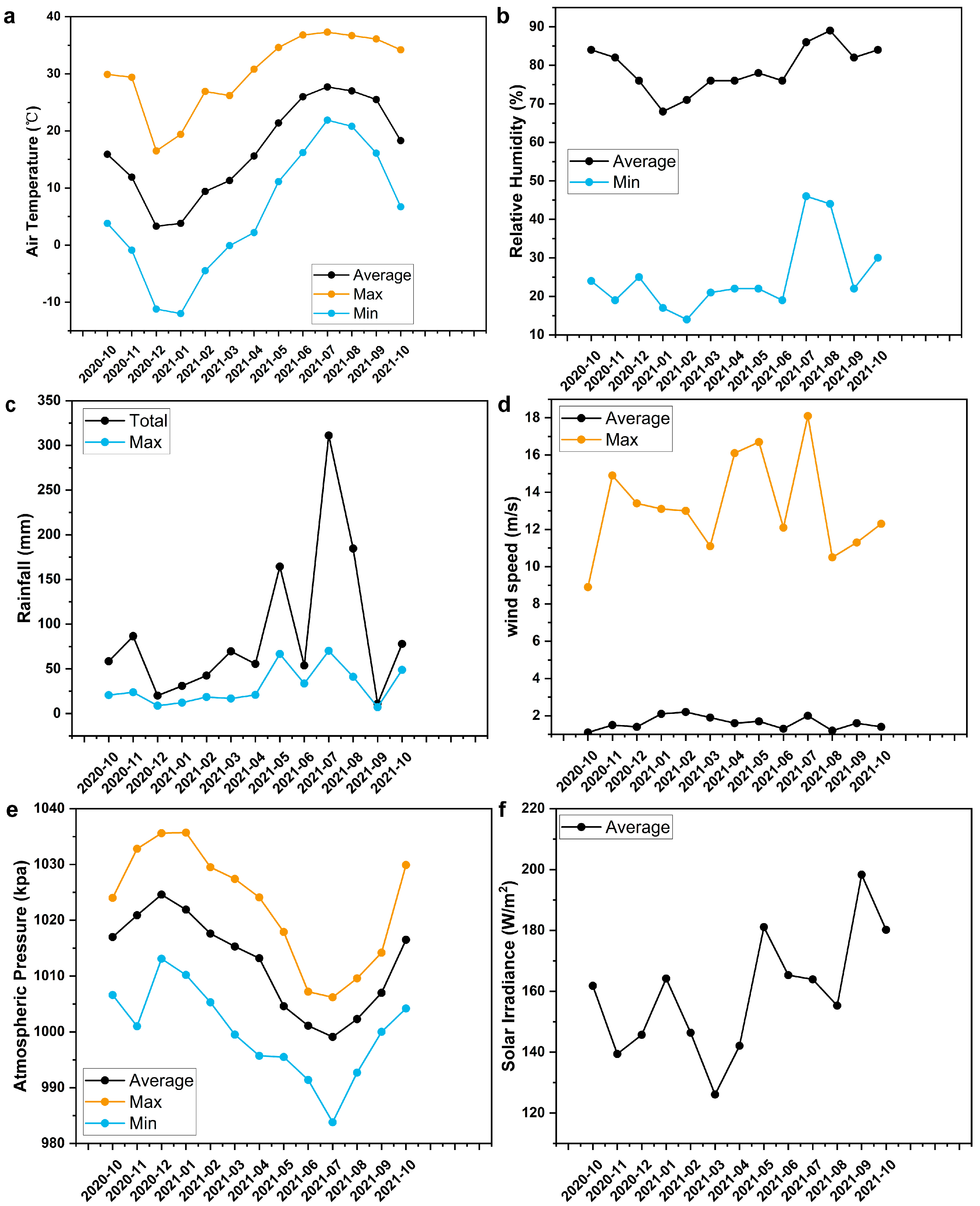
2.1. Study Site
2.2. Experimental Design
2.3. RSA
2.4. Root Decomposition Rate
2.5. Statistical Analysis
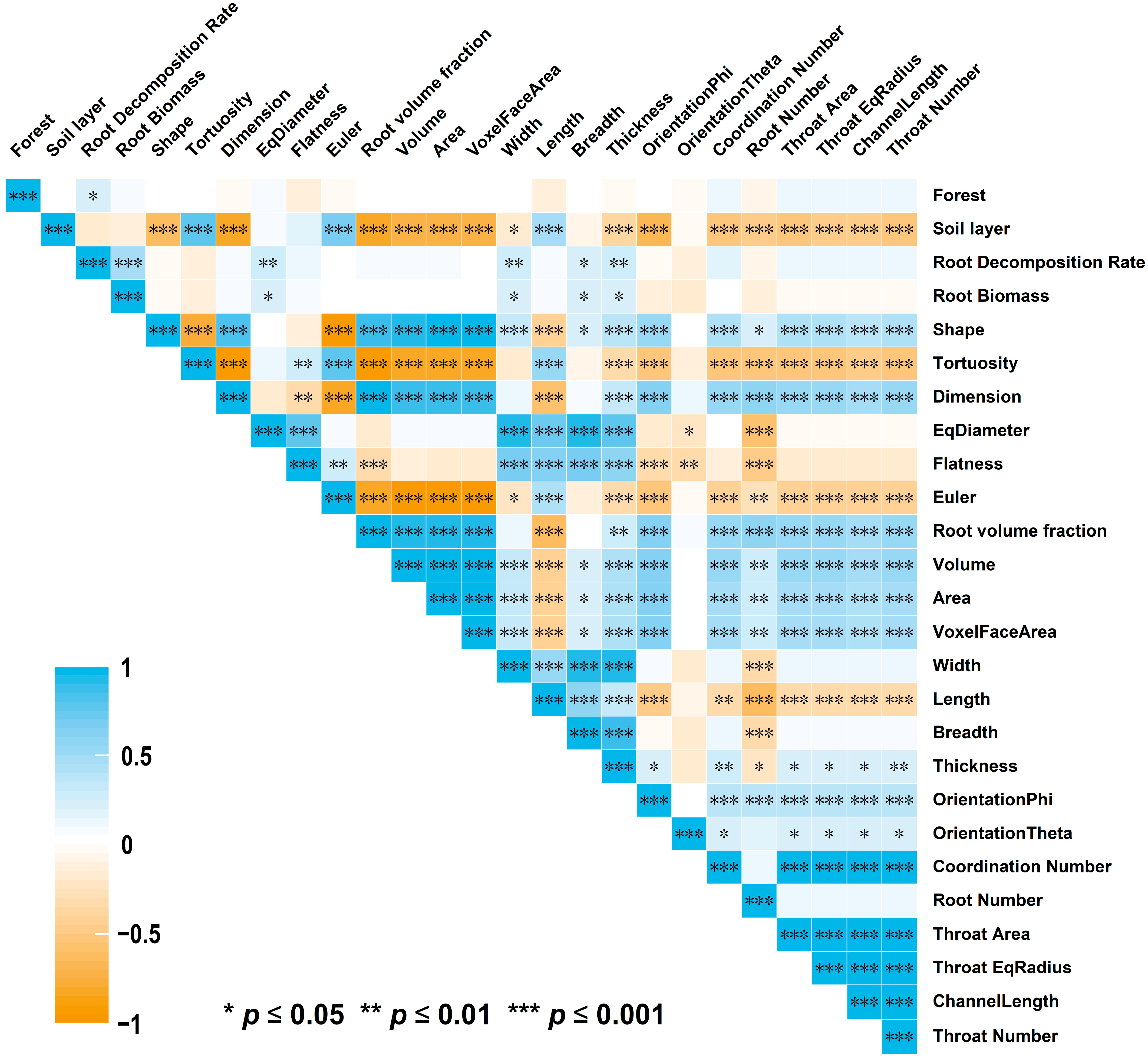
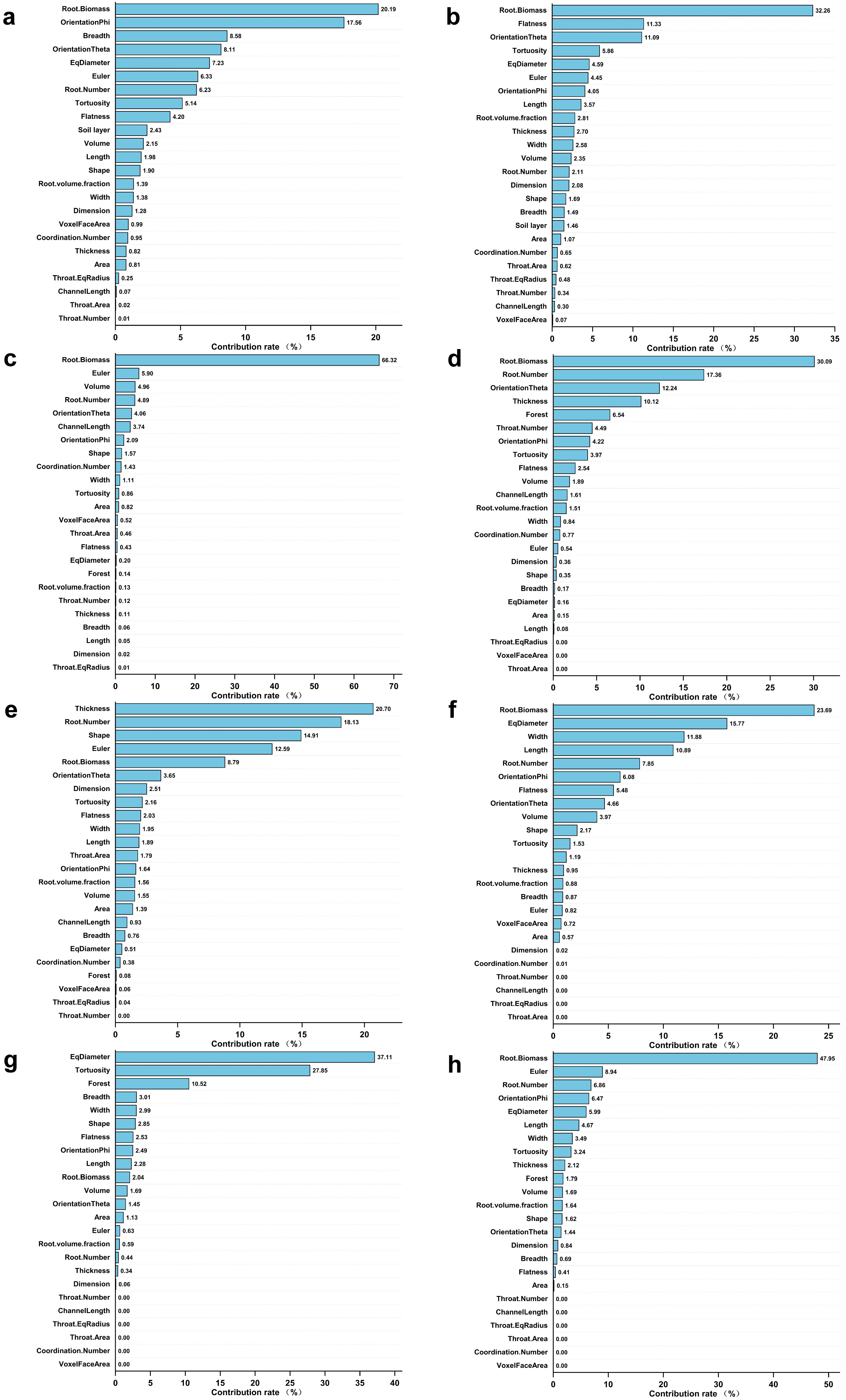
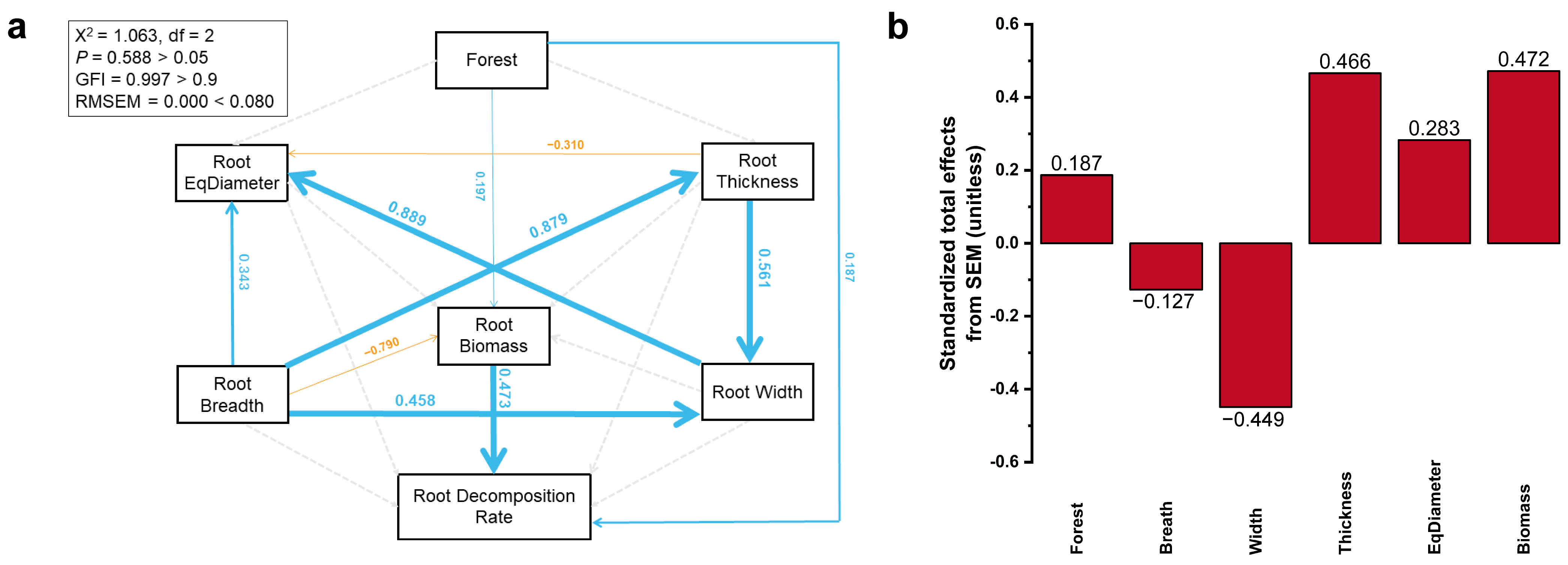
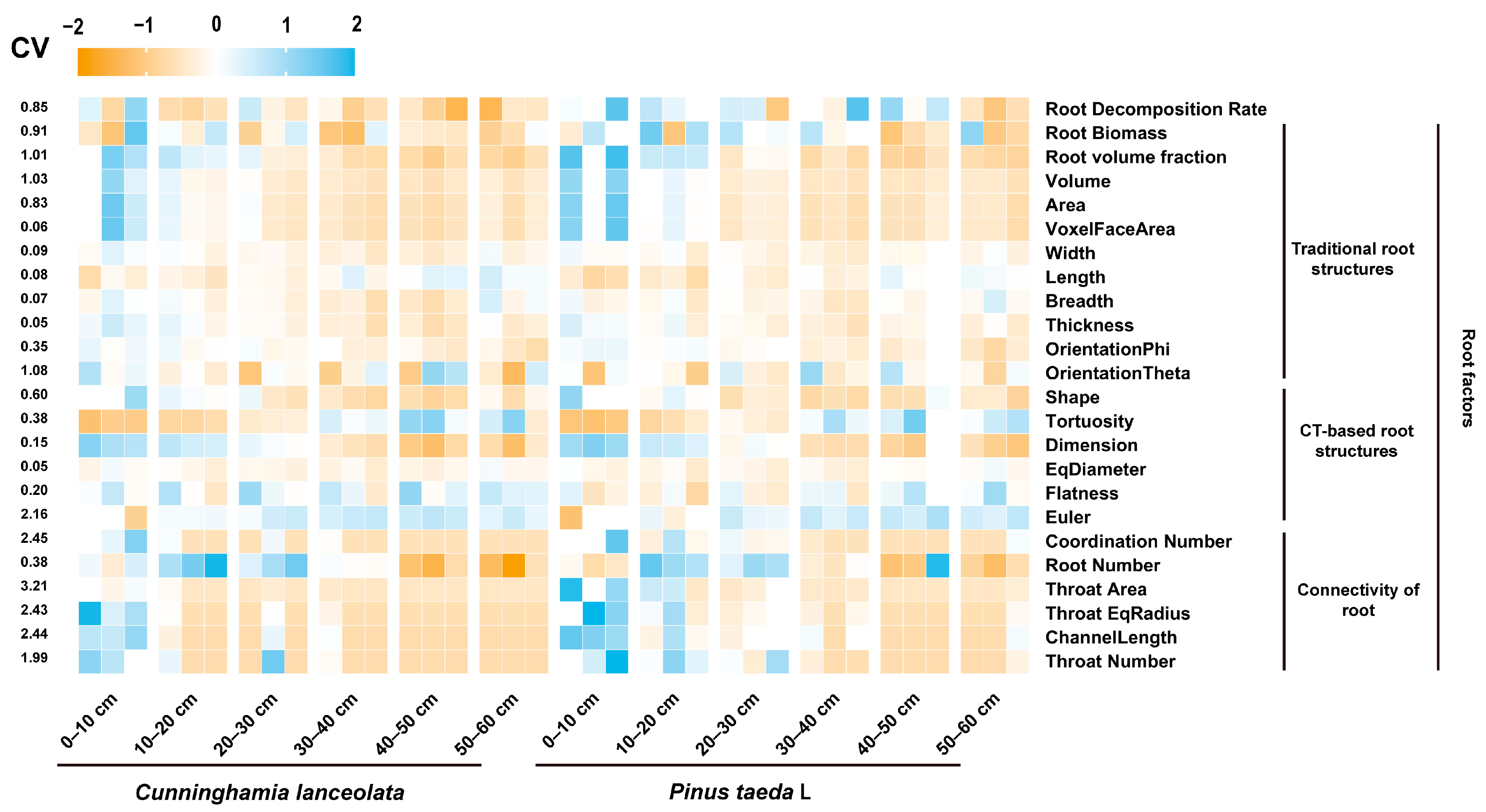
3. Results
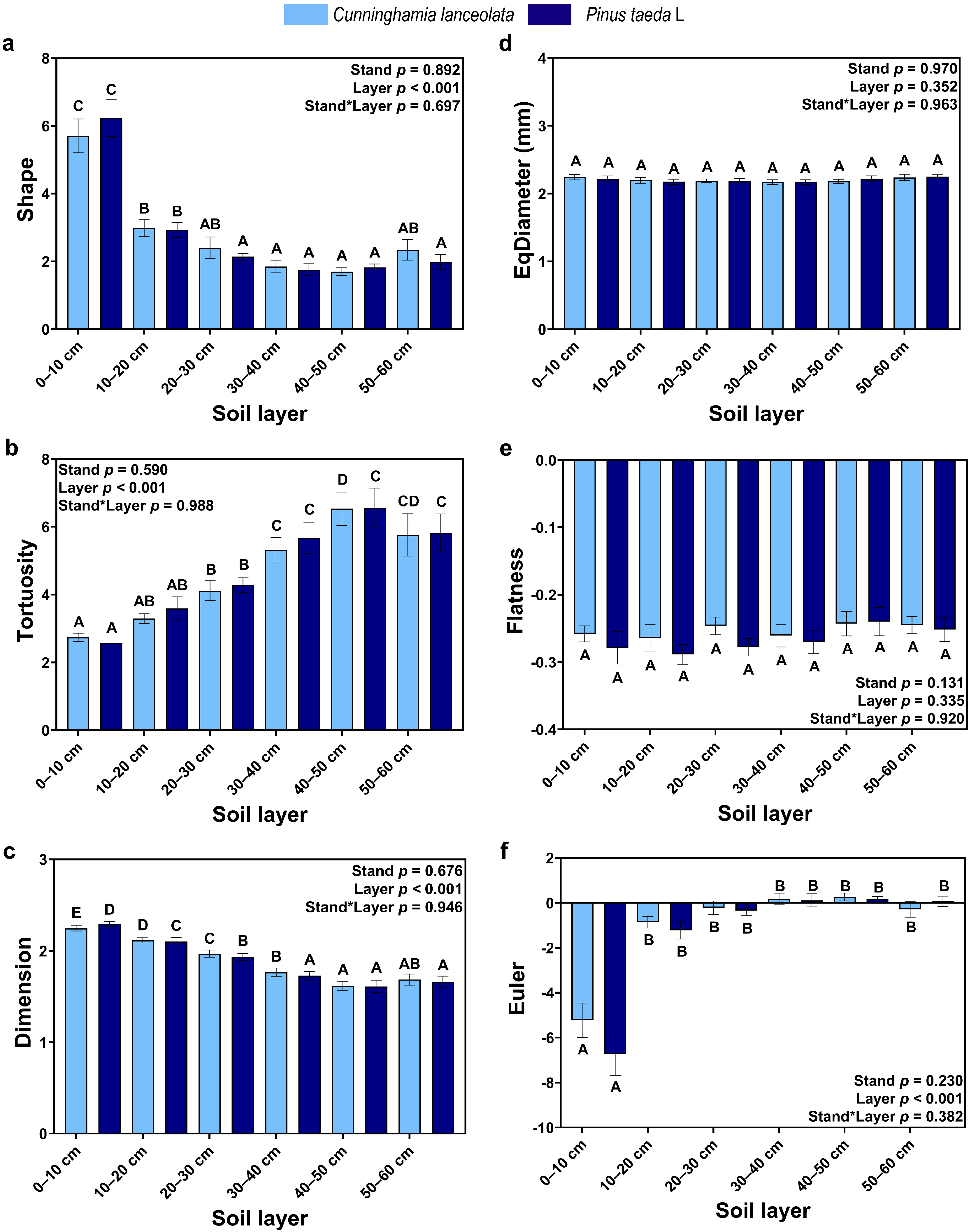
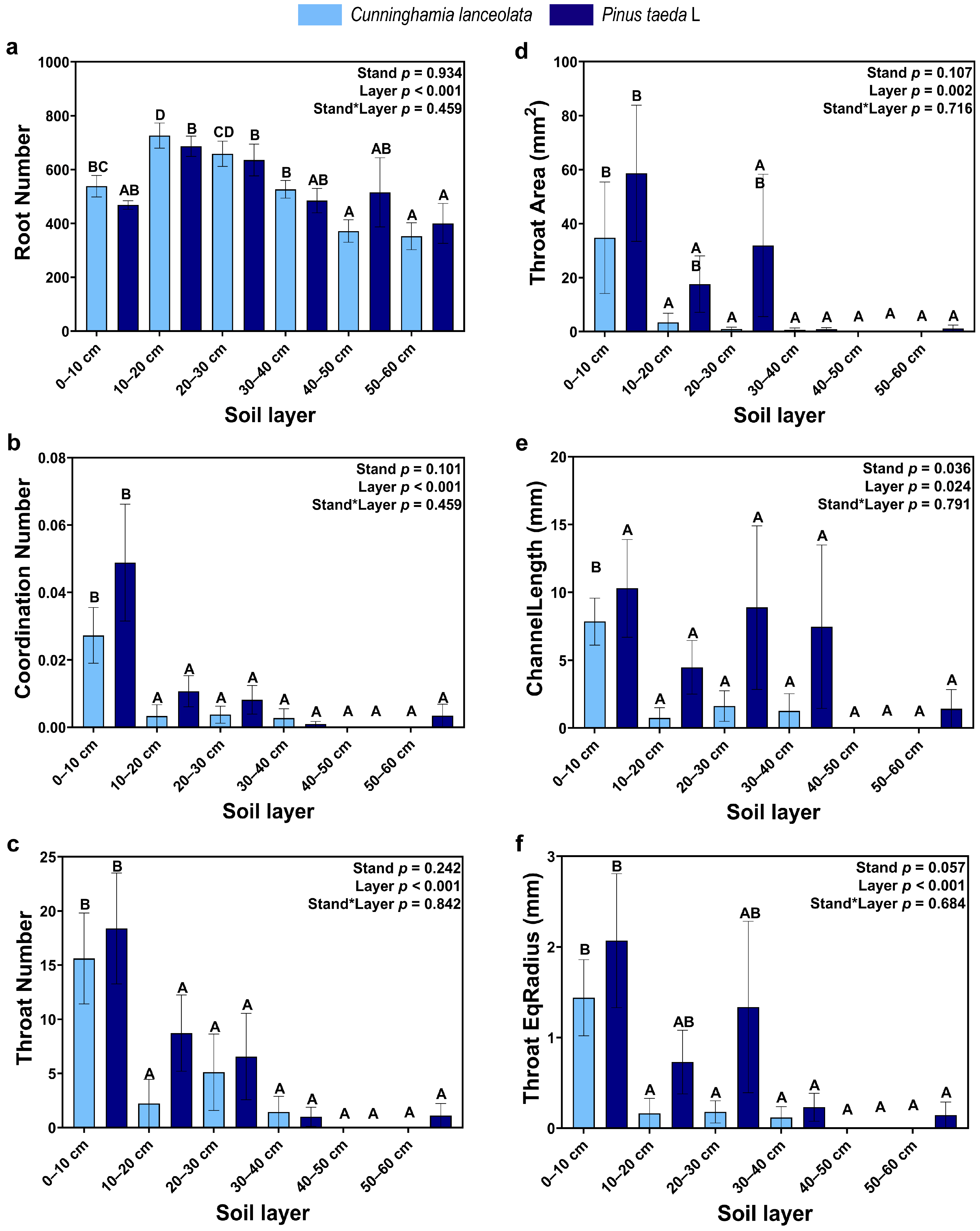
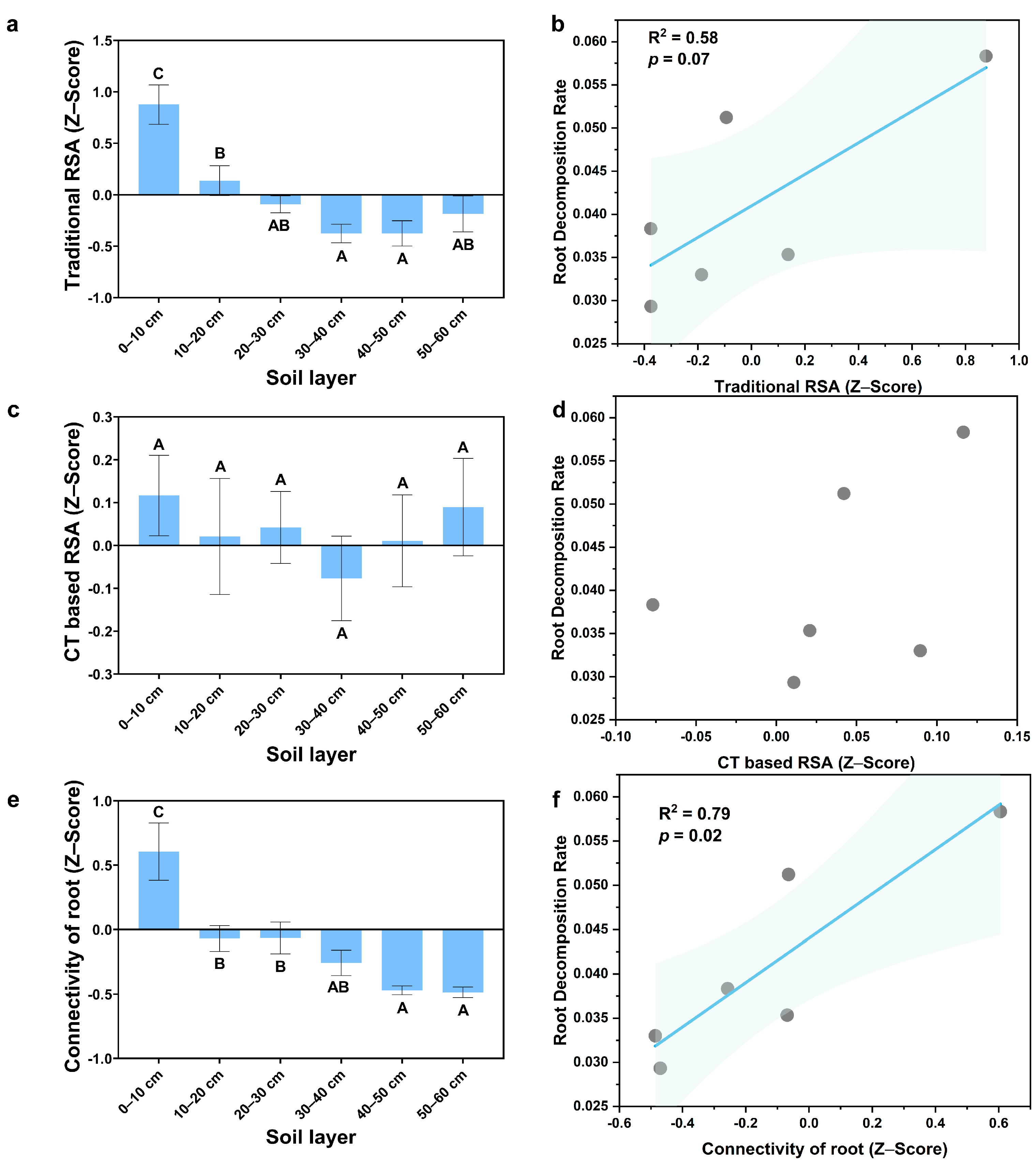
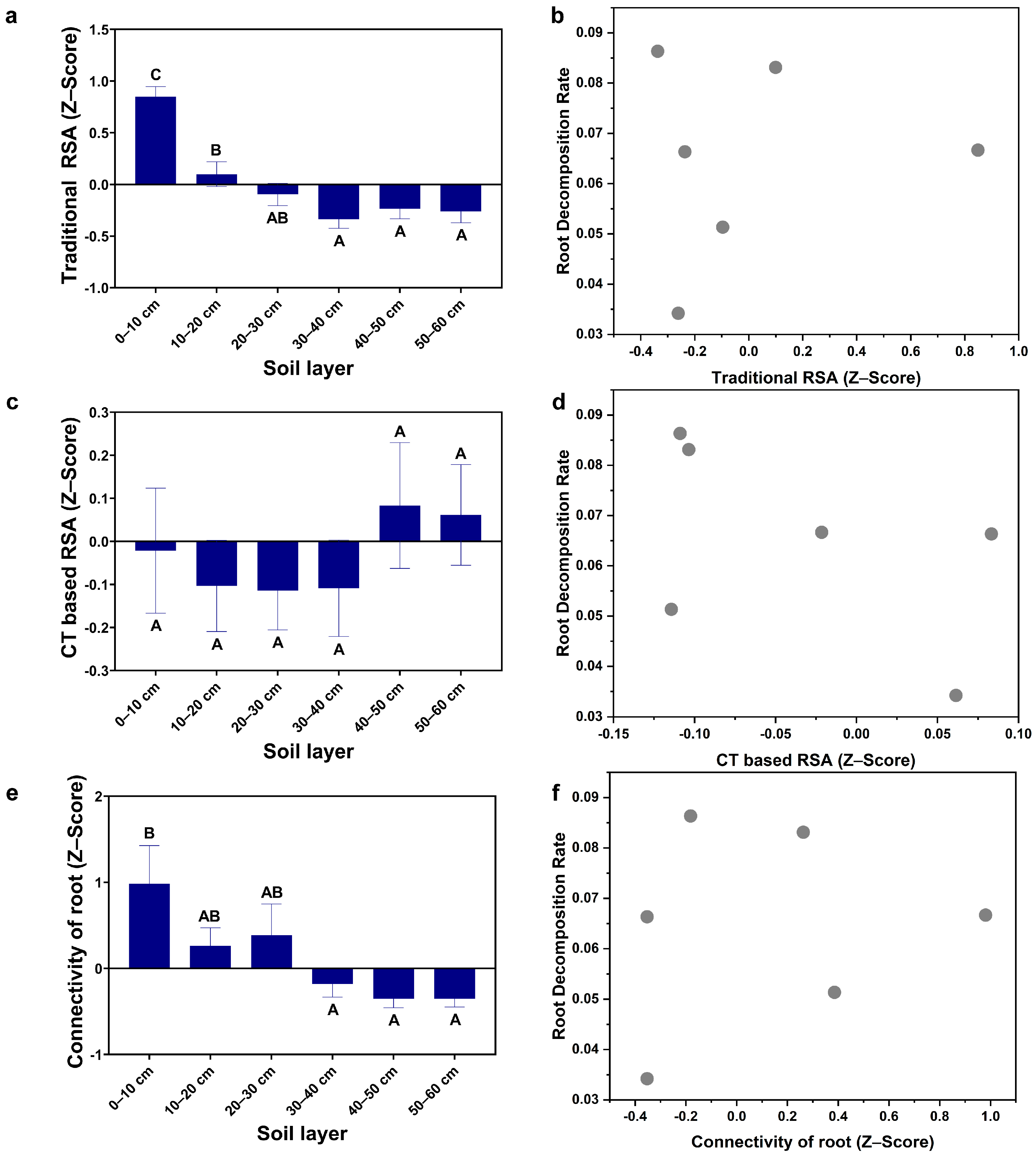
3.1. The Vertical Distribution of RSA
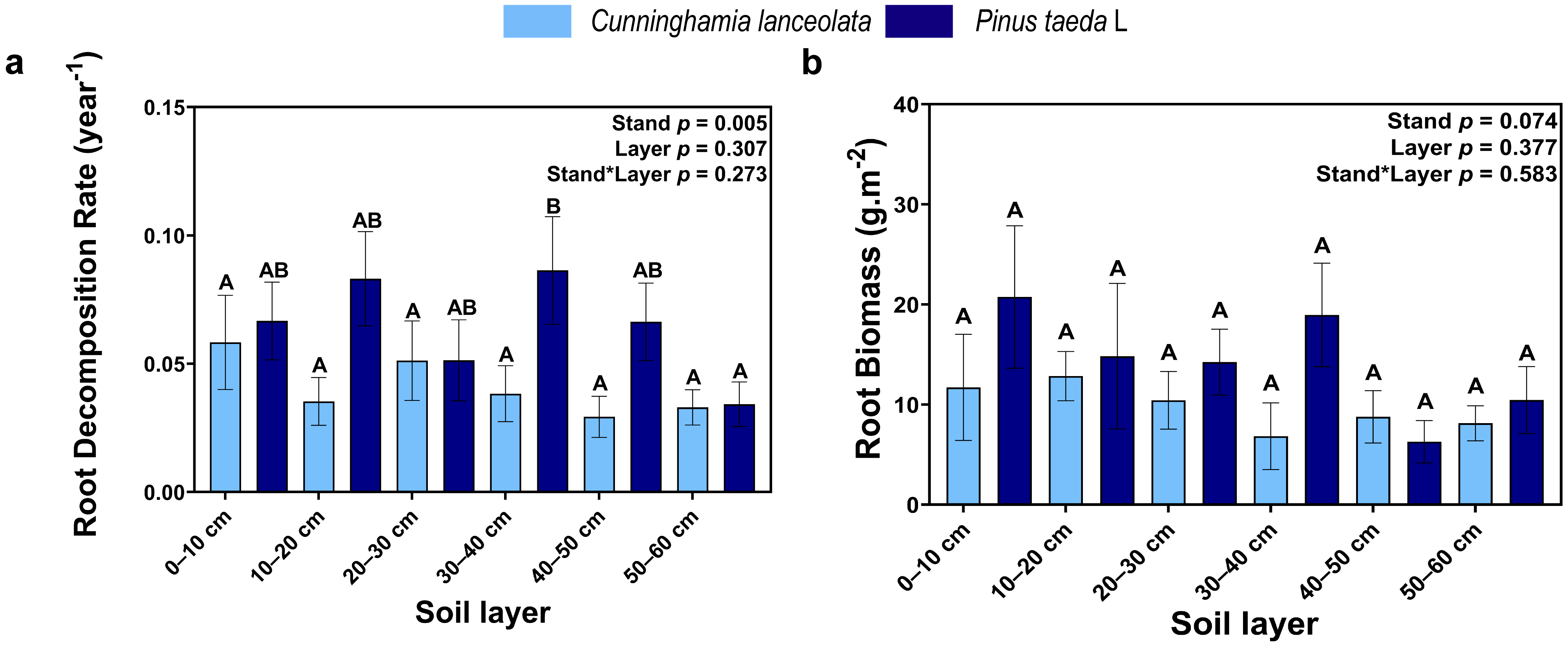
3.2. Root Decomposition Rate and Root Biomass
3.3. Major Factors Affecting the Root Decomposition Rate in Different Stand Types and Soil Depths
3.4. SEM Analysis
4. Discussion
4.1. Vertical Distribution of the RSA
4.2. Vertical Distribution of the Root Decomposition Rate
4.3. Major Factors Affecting the Root Decomposition Rate in Different Stand Types and at Different Soil Depths
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Age (Years) | Place | Aspect | Slope | ASL (m) | Density (Trees ha−2) | Mean DBH (cm) | Mean TH (m) | Mean CD (m) | Crown Density | Intrusion Ratio |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C. lanceolata C1 | 43 | 32°07′17″ N 119°13′03″ E | 347° N | 14° | 100 | 975 | 22.19 (4.83) | 12.77 (4.33) | 6.12 (2.16) | 0.82 | 8% |
| C. lanceolata C2 | 43 | 32°07′17″ N 119°13′02″ E | 350° N | 15° | 100 | 875 | 25.38 (6.72) | 14.39 (4.80) | 8.11 (3.26) | 0.89 | 12% |
| C. lanceolata C3 | 43 | 32°07′15″ N 119°13′03″ E | 214° SW | 16° | 100 | 750 | 24.06 (4.75) | 14.43 (3.19) | 6.93 (2.20) | 0.93 | 20% |
| P. taeda L. P1 | 39 | 32°07′15″ N 119°13′16″ E | 203° SW | 13° | 110 | 675 | 28.80 (5.62) | 15.11 (3.95) | 7.93 (2.72) | 0.54 | 9% |
| P. taeda L. P2 | 39 | 32°07′17″ N 119°13′18″ E | 207° SW | 10° | 110 | 500 | 29.91 (6.52) | 13.13 (3.15) | 7.75 (2.22) | 0.61 | 17% |
| P. taeda L. P3 | 39 | 32°07′17″ N 119°13′17″ E | 330° NW | 7° | 110 | 525 | 28.61 (4.98) | 14.00 (2.66) | 9.62 (1.83) | 0.63 | 13% |
| Parameters | Explanatory Notes |
|---|---|
| The volume fraction of roots | The volume of roots/volume of undisturbed soil core (The range is 0 to 1) |
| Length | Maximum Feret diameter The Feret diameter is defined as the distance between two tangent planes of a particle in a given direction |
| Breadth | Largest distance between two parallel lines touching the object without intersecting it and lying in a plane orthogonal to the maximum Feret diameter |
| Width | Minimum Feret Diameter |
| Thickness | The largest segment that touches the object by its endpoints and lying in a plane orthogonal to the maximum Feret diameter and orthogonal to the breadth diameter |
| OrientationPhi | Phi orientation of the particle in degrees [0, +90], computed with the inertia moments. It defines, with OrientationTheta, the eigenvector of the largest eigenvalue of the covariance matrix |
| OrientationTheta | Theta orientation of the particle in degrees [−180, 180], computed with the inertia moments. It defines, with OrientationPhi, the eigenvector of the largest eigenvalue of the covariance matrix |
| Shape | The shape factor is defined as shape = area3/(36 × π × volume2) |
| Tortuosity | Tortuosity is defined as the ratio between the length of the path and the distance between its ends along the z-axis. In our case, the distance between the ends of the curve is given by the number of planes along the z-axis |
| Fractal dimension | A fractal dimension is a number greater than 2 and strictly lower than 3. The result is 2 in the case of standard geometric surfaces. Applied to 3D images, the fractal dimension is an effective indicator to measure and compare the roughness of a surface. It is also a good indicator to evaluate how the curve fills the space. The less smooth the surface, the greater the fractal dimension. It can also be interpreted as a quantification of how complex the surface is and how it fills the space |
| Flatness | The ratio of the smallest to the medium eigenvalue of the covariance matrix |
| EqDiameter | The equivalent diameter is the diameter of the sphere of the same volume |
| VoxelFaceArea | This value is the sum of voxel surfaces that are on the outside of each connected component |
| Euler | Connectedness indicator. It is an indicator of the connectivity of a 3D complex structure |
References
- Teramoto, S.; Takayasu, S.; Kitomi, Y.; Arai-Sanoh, Y.; Tanabata, T.; Uga, Y. High-Throughput Three-Dimensional Visualization of Root System Architecture of Rice Using X-ray Computed Tomography. Plant Methods 2020, 16, 66. [Google Scholar] [CrossRef]
- Gregory, P.J. Plant Roots: Growth, Activity and Interactions with the Soil; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 978-1-4051-7308-7. [Google Scholar]
- Barton, C.V.M.; Montagu, K.D. Detection of Tree Roots and Determination of Root Diameters by Ground Penetrating Radar under Optimal Conditions. Tree Physiol. 2004, 24, 1323–1331. [Google Scholar] [CrossRef]
- Bert, D.; Danjon, F. Carbon Concentration Variations in the Roots, Stem and Crown of Mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Montagnoli, A.; Terzaghi, M.; Miali, A.; Chiatante, D.; Dumroese, R.K. Unusual Late-Fall Wildfire in a Pre-Alpine Fagus Sylvatica Forest Reduced Fine Roots in the Shallower Soil Layer and Shifted Very Fine-Root Growth to Deeper Soil Depth. Sci. Rep. 2023, 13, 6380. [Google Scholar] [CrossRef] [PubMed]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon Allocation in Forest Ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Onelli, E.; Scippa, G.S.; Chiatante, D. Seasonality of Fine Root Dynamics and Activity of Root and Shoot Vascular Cambium in a Quercus ilex L. Forest (Italy). For. Ecol. Manag. 2019, 431, 26–34. [Google Scholar] [CrossRef]
- Kondori, A.A.; Vajari, K.A.; Feizian, M.; Montagnoli, A.; Di Iorio, A. Gap Size in Hyrcanian Forest Affects the Lignin and N Concentrations of the Oriental Beech (Fagus orientalis Lipsky) Fine Roots but Does Not Change Their Morphological Traits in the Medium Term. Forests 2021, 12, 137. [Google Scholar] [CrossRef]
- Bontpart, T.; Concha, C.; Giuffrida, M.V.; Robertson, I.; Admkie, K.; Degefu, T.; Girma, N.; Tesfaye, K.; Haileselassie, T.; Fikre, A.; et al. Affordable and Robust Phenotyping Framework to Analyse Root System Architecture of Soil-Grown Plants. Plant J. 2020, 103, 2330–2343. [Google Scholar] [CrossRef]
- Rogers, E.D.; Monaenkova, D.; Mijar, M.; Nori, A.; Goldman, D.I.; Benfey, P.N. X-ray Computed Tomography Reveals the Response of Root System Architecture to Soil Texture. Plant Physiol. 2016, 171, 2028–2040. [Google Scholar] [CrossRef]
- Fang, S.; Yan, X.; Liao, H. 3D Reconstruction and Dynamic Modeling of Root Architecture in Situ and Its Application to Crop Phosphorus Research. Plant J. 2009, 60, 1096–1108. [Google Scholar] [CrossRef]
- Bauerle, T.L.; Centinari, M. Assessment of Root System Development among Four Ornamental Tree Species through Time via X-ray Computed Tomography. HortScience 2014, 49, 44–50. [Google Scholar] [CrossRef]
- Pang, Y.; Tian, J.; Yang, H.; Zhang, K.; Wang, D. Responses of Fine Roots at Different Soil Depths to Different Thinning Intensities in a Secondary Forest in the Qinling Mountains, China. Biology 2022, 11, 351. [Google Scholar] [CrossRef]
- Hodge, A. The Plastic Plant: Root Responses to Heterogeneous Supplies of Nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Montagnoli, A.; Lasserre, B.; Terzaghi, M.; Byambadorj, S.-O.; Nyam-Osor, B.; Scippa, G.S.; Chiatante, D. Fertilization Reduces Root Architecture Plasticity in Ulmus pumila Used for Afforesting Mongolian Semi-Arid Steppe. Front. Plant Sci. 2022, 13, 878299. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; Nie, Y.; Bai, S.H.; Zhou, L.; Shao, J.; Cheng, W.; Wang, J.; Hu, F.; Fu, Y. Drought-Induced Changes in Root Biomass Largely Result from Altered Root Morphological Traits: Evidence from a Synthesis of Global Field Trials. Plant Cell Environ. 2018, 41, 2589–2599. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Chen, H.Y.H. Positive Species Mixture Effects on Fine Root Turnover and Mortality in Natural Boreal Forests. Soil Biol. Biochem. 2018, 121, 130–137. [Google Scholar] [CrossRef]
- Nyam-Osor, B.; Byambadorj, S.-O.; Park, B.B.; Terzaghi, M.; Scippa, G.S.; Stanturf, J.A.; Chiatante, D.; Montagnoli, A. Root Biomass Distribution of Populus sibirica and Ulmus pumila Afforestation Stands Is Affected by Watering Regimes and Fertilization in the Mongolian Semi-Arid Steppe. Front. Plant Sci. 2021, 12, 638828. [Google Scholar] [CrossRef] [PubMed]
- Montagnoli, A.; Chiatante, D.; Godbold, D.L.; Koike, T.; Rewald, B.; Dumroese, R.K. Editorial: Modulation of Growth and Development of Tree Roots in Forest Ecosystems. Front. Plant Sci. 2022, 13, 850163. [Google Scholar] [CrossRef]
- Kaestner, A.; Schneebeli, M.; Graf, F. Visualizing Three-Dimensional Root Networks Using Computed Tomography. Geoderma 2006, 136, 459–469. [Google Scholar] [CrossRef]
- Peng, S.; Chen, H.Y.H. Global Responses of Fine Root Biomass and Traits to Plant Species Mixtures in Terrestrial Ecosystems. Glob. Ecol. Biogeogr. 2021, 30, 289–304. [Google Scholar] [CrossRef]
- Pierret, A.; Doussan, C.; Capowiez, Y.; Bastardie, F.; Pagès, L. Root Functional Architecture: A Framework for Modeling the Interplay between Roots and Soil. Vadose Zone J. 2007, 6, 269–281. [Google Scholar] [CrossRef]
- de Graaff, M.-A.; Six, J.; Jastrow, J.D.; Schadt, C.W.; Wullschleger, S.D. Variation in Root Architecture among Switchgrass Cultivars Impacts Root Decomposition Rates. Soil Biol. Biochem. 2013, 58, 198–206. [Google Scholar] [CrossRef]
- Lontoc-Roy, M.; Dutilleul, P.; Prasher, S.O.; Han, L.; Brouillet, T.; Smith, D.L. Advances in the Acquisition and Analysis of CT Scan Data to Isolate a Crop Root System from the Soil Medium and Quantify Root System Complexity in 3-D Space. Geoderma 2006, 137, 231–241. [Google Scholar] [CrossRef]
- Ding, Y.; Leppälammi-Kujansuu, J.; Helmisaari, H.-S. Fine Root Longevity and Below- and Aboveground Litter Production in a Boreal Betula Pendula Forest. For. Ecol. Manag. 2019, 431, 17–25. [Google Scholar] [CrossRef]
- Li, A.; Zhu, L.; Xu, W.; Liu, L.; Teng, G. Recent Advances in Methods for in Situ Root Phenotyping. PeerJ 2022, 10, e13638. [Google Scholar] [CrossRef]
- Dupuy, L.X.; Fourcaud, T.; Lac, P.; Stokes, A. A Generic 3D Finite Element Model of Tree Anchorage Integrating Soil Mechanics and Real Root System Architecture. Am. J. Bot. 2007, 94, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Chen, J.; Cui, X.; Fan, B.; Lin, H. Application of Ground Penetrating Radar for Coarse Root Detection and Quantification: A Review. Plant Soil 2013, 362, 1–23. [Google Scholar] [CrossRef]
- Smith, A.; Astrup, R.; Raumonen, P.; Liski, J.; Krooks, A.; Kaasalainen, S.; Åkerblom, M.; Kaasalainen, M. Tree Root System Characterization and Volume Estimation by Terrestrial Laser Scanning and Quantitative Structure Modeling. Forests 2014, 5, 3274–3294. [Google Scholar] [CrossRef]
- Jose, S.; Gillespie, A.R.; Seifert, J.R.; Pope, P.E. Comparison of Minirhizotron and Soil Core Methods for Quantifying Root Biomass in a Temperate Alley Cropping System. Agrofor. Syst. 2001, 52, 161–168. [Google Scholar] [CrossRef]
- Saltveit, M.E.; Young, E. A Method for Studying the Three-Dimensional Distribution of Roots Grown in an Artificial Medium. J. Am. Soc. Horticult. Sci. 1983, 108, 1023–1025. [Google Scholar] [CrossRef]
- Burr-Hersey, J.E.; Mooney, S.J.; Bengough, A.G.; Mairhofer, S.; Ritz, K. Developmental Morphology of Cover Crop Species Exhibit Contrasting Behaviour to Changes in Soil Bulk Density, Revealed by X-ray Computed Tomography. PLoS ONE 2017, 12, e0181872. [Google Scholar] [CrossRef]
- Hu, X.; Li, X.-Y.; Zhao, Y.-D.; Cheng, Y.-Q.; Gao, Z.; Yang, Z.-G. Identification of Water Flow through Non-Root Soil Macropores and along Roots in Shrub-Encroached Grassland. Eur. J. Soil Sci. 2022, 73, e13260. [Google Scholar] [CrossRef]
- Suresh, N.; Stephens, S.A.; Adams, L.; Beck, A.N.; McKinney, A.L.; Varga, T. Extracting Metrics for Three-Dimensional Root Systems: Volume and Surface Analysis from In-Soil X-ray Computed Tomography Data. J. Vis. Exp. 2016, 110, 53788. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources 2014: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014; ISBN 978-92-5-108369-7.
- Dornbush, M.E.; Isenhart, T.M.; Raich, J.W. Quantifying Fine-Root Decomposition: An Alternative to Buried Litterbags. Ecology 2002, 83, 2985–2990. [Google Scholar] [CrossRef]
- Li, X.; Zheng, X.; Zhou, Q.; McNulty, S.; King, J.S. Measurements of Fine Root Decomposition Rate: Method Matters. Soil Biol. Biochem. 2022, 164, 108482. [Google Scholar] [CrossRef]
- Katuwal, S.; Norgaard, T.; Moldrup, P.; Lamandé, M.; Wildenschild, D.; de Jonge, L.W. Linking Air and Water Transport in Intact Soils to Macropore Characteristics Inferred from X-ray Computed Tomography. Geoderma 2015, 237–238, 9–20. [Google Scholar] [CrossRef]
- Olson, J.S. Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Aulen, M.; Shipley, B.; Bradley, R. Prediction of in Situ Root Decomposition Rates in an Interspecific Context from Chemical and Morphological Traits. Ann. Bot. 2012, 109, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Delgado-Baquerizo, M.; Luo, X.; Liu, Y.; Van Nostrand, J.D.; Chen, W.; Zhou, J.; Huang, Q. Soil Aggregate Size-Dependent Relationships between Microbial Functional Diversity and Multifunctionality. Soil Biol. Biochem. 2021, 154, 108143. [Google Scholar] [CrossRef]
- Li, C.; Yu, Z.; Lin, J.; Meng, M.; Zhao, Y.; Jia, Z.; Peng, X.; Liu, X.; Zhang, J. Forest Conversion and Soil Depth Can Modify the Contributions of Organic and Inorganic Colloids to the Stability of Soil Aggregates. Forests 2022, 13, 546. [Google Scholar] [CrossRef]
- Chen, W.; Chen, Y.; Siddique, K.H.M.; Li, S. Root Penetration Ability and Plant Growth in Agroecosystems. Plant Physiol. Biochem. 2022, 183, 160–168. [Google Scholar] [CrossRef]
- Materechera, S.A.; Alston, A.M.; Kirby, J.M.; Dexter, A.R. Influence of Root Diameter on the Penetration of Seminal Roots into a Compacted Subsoil. Plant Soil 1992, 144, 297–303. [Google Scholar] [CrossRef]
- Popova, L.; van Dusschoten, D.; Nagel, K.A.; Fiorani, F.; Mazzolai, B. Plant Root Tortuosity: An Indicator of Root Path Formation in Soil with Different Composition and Density. Ann. Bot. 2016, 118, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, Y.; An, Z.; Li, R.; Xu, Y.; Zhang, P.; Yang, Y.; Wang, T. Deep Root Information “Hidden in the Dark”: A Case Study on the 21-m Soil Profile of Robinia pseudoacacia in the Critical Zone of the Chinese Loess Plateau. CATENA 2022, 213, 106121. [Google Scholar] [CrossRef]
- Hicks Pries, C.E.; Sulman, B.N.; West, C.; O’Neill, C.; Poppleton, E.; Porras, R.C.; Castanha, C.; Zhu, B.; Wiedemeier, D.B.; Torn, M.S. Root Litter Decomposition Slows with Soil Depth. Soil Biol. Biochem. 2018, 125, 103–114. [Google Scholar] [CrossRef]
- Han, S.H.; Kim, S.; Chang, H.; Kim, H.-J.; Khamzina, A.; Son, Y. Soil Depth- and Root Diameter-Related Variations Affect Root Decomposition in Temperate Pine and Oak Forests. J. Plant Ecol. 2019, 12, 871–881. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Cao, J.; Fu, S.; Wang, J.; Lambers, H.; Liu, Z. Divergent Responses of Fine Root Decomposition to Removal of Understory Plants and Overstory Trees in Subtropical Eucalyptus Urophylla Plantations. Plant Soil 2022, 476, 639–652. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Laskowski, M.J.; Burton, A.J.; Lessard, V.C.; Zak, D.R. Variation in Sugar Maple Root Respiration with Root Diameter and Soil Depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Birouste, M.; Kazakou, E.; Blanchard, A.; Roumet, C. Plant Traits and Decomposition: Are the Relationships for Roots Comparable to Those for Leaves? Ann. Bot. 2012, 109, 463–472. [Google Scholar] [CrossRef]
- Donovan, L.A.; Mason, C.M.; Bowsher, A.W.; Goolsby, E.W.; Ishibashi, C.D.A. Ecological and Evolutionary Lability of Plant Traits Affecting Carbon and Nutrient Cycling. J. Ecol. 2014, 102, 302–314. [Google Scholar] [CrossRef]
- Smith, S.W.; Woodin, S.J.; Pakeman, R.J.; Johnson, D.; van der Wal, R. Root Traits Predict Decomposition across a Landscape-Scale Grazing Experiment. New Phytol. 2014, 203, 851–862. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, X.; Tao, N.; Ao, D.; Zeng, W.; Qian, Y.; Zeng, H. Effects of Litter Types, Microsite and Root Diameters on Litter Decomposition in Pinus sylvestris Plantations of Northern China. Plant Soil 2014, 374, 677–688. [Google Scholar] [CrossRef]
- Djukic, I.; Kepfer-Rojas, S.; Schmidt, I.K.; Larsen, K.S.; Beier, C.; Berg, B.; Verheyen, K.; Caliman, A.; Paquette, A.; Gutiérrez-Girón, A.; et al. Early Stage Litter Decomposition across Biomes. Sci. Total Environ. 2018, 628–629, 1369–1394. [Google Scholar] [CrossRef] [PubMed]
- Harmon, M.E.; Silver, W.L.; Fasth, B.; Chen, H.; Burke, I.C.; Parton, W.J.; Hart, S.C.; Currie, W.S. Lidet Long-Term Patterns of Mass Loss during the Decomposition of Leaf and Fine Root Litter: An Intersite Comparison. Glob. Chang. Biol. 2009, 15, 1320–1338. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Oleksyn, J.; Eissenstat, D.M.; Reich, P.B. Fine Root Decomposition Rates Do Not Mirror Those of Leaf Litter among Temperate Tree Species. Oecologia 2010, 162, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lin, L.; Wang, H.; Zhang, Z.; Shangguan, Z.; Feng, X.; He, J.-S. Simulating Warmer and Drier Climate Increases Root Production but Decreases Root Decomposition in an Alpine Grassland on the Tibetan Plateau. Plant Soil 2021, 458, 59–73. [Google Scholar] [CrossRef]
- Santonja, M.; Milcu, A.; Fromin, N.; Rancon, A.; Shihan, A.; Fernandez, C.; Baldy, V.; Hättenschwiler, S. Temporal Shifts in Plant Diversity Effects on Carbon and Nitrogen Dynamics During Litter Decomposition in a Mediterranean Shrubland Exposed to Reduced Precipitation. Ecosystems 2019, 22, 939–954. [Google Scholar] [CrossRef]
- Guerrero-Ramírez, N.R.; Craven, D.; Messier, C.; Potvin, C.; Turner, B.L.; Handa, I.T. Root Quality and Decomposition Environment, but Not Tree Species Richness, Drive Root Decomposition in Tropical Forests. Plant Soil 2016, 404, 125–139. [Google Scholar] [CrossRef]
- Prieto, I.; Stokes, A.; Roumet, C. Root Functional Parameters Predict Fine Root Decomposability at the Community Level. J. Ecol. 2016, 104, 725–733. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of Changing Biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef]
- Ohashi, M.; Makita, N.; Katayama, A.; Kume, T.; Matsumoto, K.; Kumagai, T.; Endo, I.; Kho, L.K. Characteristics of Root Decomposition Based on in Situ Experiments in a Tropical Rainforest in Sarawak, Malaysia: Impacts of Root Diameter and Soil Biota. Plant Soil 2019, 436, 439–448. [Google Scholar] [CrossRef]
- Camiré, C.; Côté, B.; Brulotte, S. Decomposition of Roots of Black Alder and Hybrid Poplar in Short-Rotation Plantings: Nitrogen and Lignin Control. Plant Soil 1991, 138, 123–132. [Google Scholar] [CrossRef]
- Zhuang, L.; Yang, W.; Wu, F.; Tan, B.; Zhang, L.; Yang, K.; He, R.; Li, Z.; Xu, Z. Diameter-Related Variations in Root Decomposition of Three Common Subalpine Tree Species in Southwestern China. Geoderma 2018, 311, 1–8. [Google Scholar] [CrossRef]
- Sun, T.; Mao, Z.; Han, Y. Slow Decomposition of Very Fine Roots and Some Factors Controlling the Process: A 4-Year Experiment in Four Temperate Tree Species. Plant Soil 2013, 372, 445–458. [Google Scholar] [CrossRef]
- Wang, P.; Guo, J.; Xu, X.; Yan, X.; Zhang, K.; Qiu, Y.; Zhao, Q.; Huang, K.; Luo, X.; Yang, F.; et al. Soil Acidification Alters Root Morphology, Increases Root Biomass but Reduces Root Decomposition in an Alpine Grassland. Environ. Pollut. 2020, 265, 115016. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B. The World-Wide ‘Fast–Slow’ Plant Economics Spectrum: A Traits Manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Sun, T.; Dong, L.; Zhang, L.; Wu, Z.; Wang, Q.; Li, Y.; Zhang, H.; Wang, Z. Early Stage Fine-Root Decomposition and Its Relationship with Root Order and Soil Depth in a Larix Gmelinii Plantation. Forests 2016, 7, 234. [Google Scholar] [CrossRef]








| Forest Type | Soil Layer | 4 Months | 8 Months | 12 Months | Root Decomposition Rate (Year−1) |
|---|---|---|---|---|---|
| Cunninghamia lanceolata | 0–10 cm | 8.17% (±8.19%) Aa | 5.78% (±3.80%) Ba | 4.93% (±4.37%) Aa | 0.058 (±0.06) A |
| 10–20 cm | 11.00% (±8.16%) Ab | 4.66% (±3.73%) Bab | 4.23% (±3.22%) Aa | 0.035 (±0.028) A | |
| 20–30 cm | 8.10% (±8.16%) Aa | 3.83% (±3.73%) ABa | 4.33% (±3.22%) Aa | 0.051 (±0.047) A | |
| 30–40 cm | 8.37% (±6.55%) Ab | 3.33% (±1.44%) ABa | 2.11% (±1.41%) Aa | 0.038 (±0.033) A | |
| 40–50 cm | 8.32% (±8.47%) Aa | 1.25% (±1.36%) Aa | 4.45% (±7.29%) Aa | 0.029 (±0.024) A | |
| 50–60 cm | 8.91% (±8.95%) Ab | 1.65% (±1.13%) Aa | 1.05% (±1.47%) Aa | 0.033 (±0.021) A | |
| Pinus taeda | 0–10 cm | 14.79% (±9.28%) Ab | 4.8% (±2.66%) ABa | 9.92% (±6.04%) Bab | 0.067 (±0.046) AB |
| 10–20 cm | 12.00% (±8.73%) Aa | 8.84% (±5.91%) Ba | 5.74% (±2.75%) ABa | 0.083 (±0.055) AB | |
| 20–30 cm | 11.31% (±10.92%) Aa | 7.91% (±7.53%) Ba | 4.41% (±3.99%) ABa | 0.051 (±0.047) AB | |
| 30–40 cm | 7.18% (±9.77%) Aa | 3.97% (±3.24%) ABa | 2.07% (±2.64%) Aa | 0.086 (±0.063) B | |
| 40–50 cm | 4.40% (±3.54%) Aa | 1.65% (±1.71%) Aa | 2.76% (±1.98%) Aa | 0.066 (±0.045) B | |
| 50–60 cm | 5.82% (±6.34%) Aa | 5.87% (±6.21%) ABa | 4.14% (±9.37%) ABa | 0.034 (±0.026) A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Liu, X.; Lian, J.; Cheng, X.; Wang, G.G.; Zhang, J. Soil Depth Can Modify the Contribution of Root System Architecture to the Root Decomposition Rate. Forests 2023, 14, 1092. https://doi.org/10.3390/f14061092
Tang Y, Liu X, Lian J, Cheng X, Wang GG, Zhang J. Soil Depth Can Modify the Contribution of Root System Architecture to the Root Decomposition Rate. Forests. 2023; 14(6):1092. https://doi.org/10.3390/f14061092
Chicago/Turabian StyleTang, Yingzhou, Xin Liu, Jingwei Lian, Xuefei Cheng, G. Geoff Wang, and Jinchi Zhang. 2023. "Soil Depth Can Modify the Contribution of Root System Architecture to the Root Decomposition Rate" Forests 14, no. 6: 1092. https://doi.org/10.3390/f14061092
APA StyleTang, Y., Liu, X., Lian, J., Cheng, X., Wang, G. G., & Zhang, J. (2023). Soil Depth Can Modify the Contribution of Root System Architecture to the Root Decomposition Rate. Forests, 14(6), 1092. https://doi.org/10.3390/f14061092