Community Abundance of Resprouting in Woody Plants Reflects Fire Return Time, Intensity, and Type


Abstract
1. Introduction
2. Methods
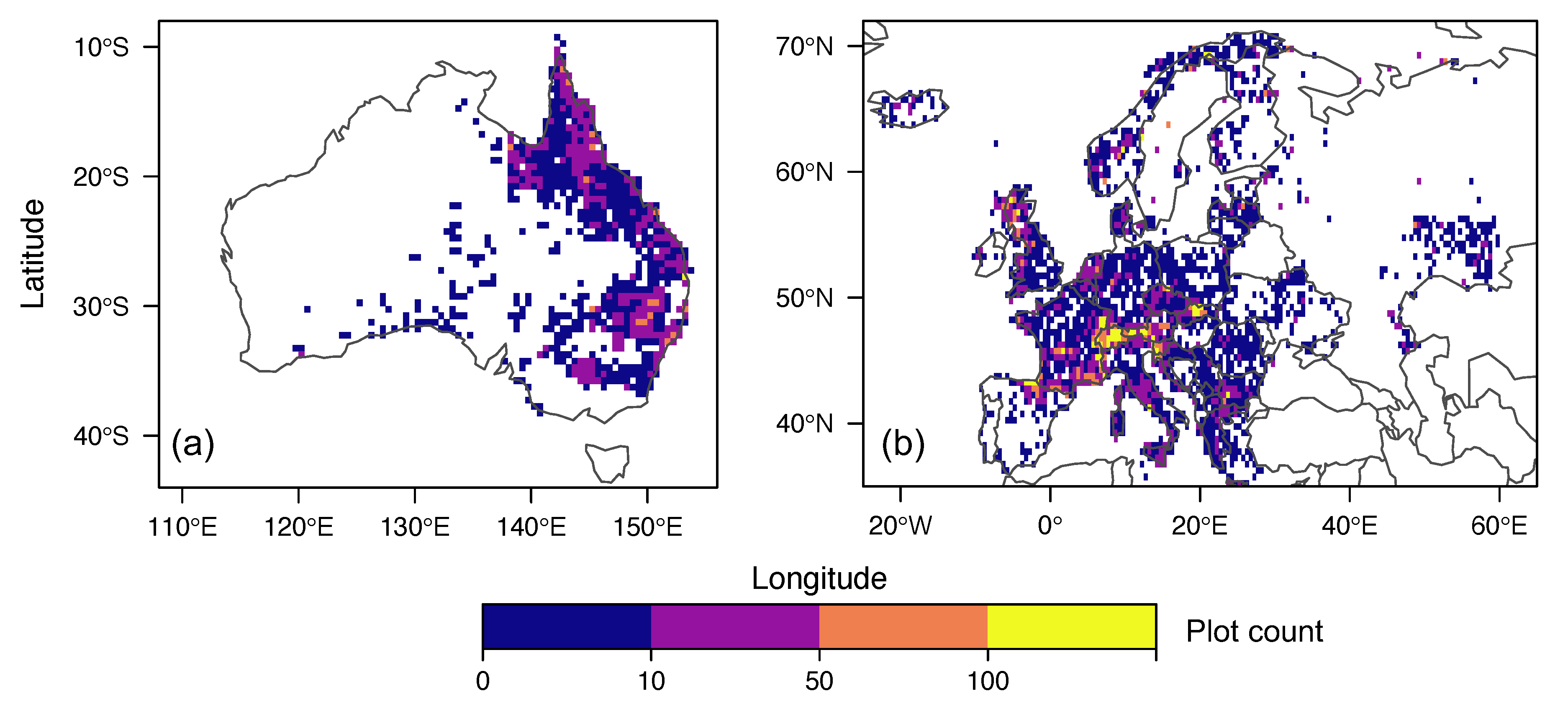
2.1. Species Cover and Resprouting Information
2.2. Fire Return Interval
2.3. Analysis of Relationships between the Incidence of Resprouters and Fire Properties
3. Results
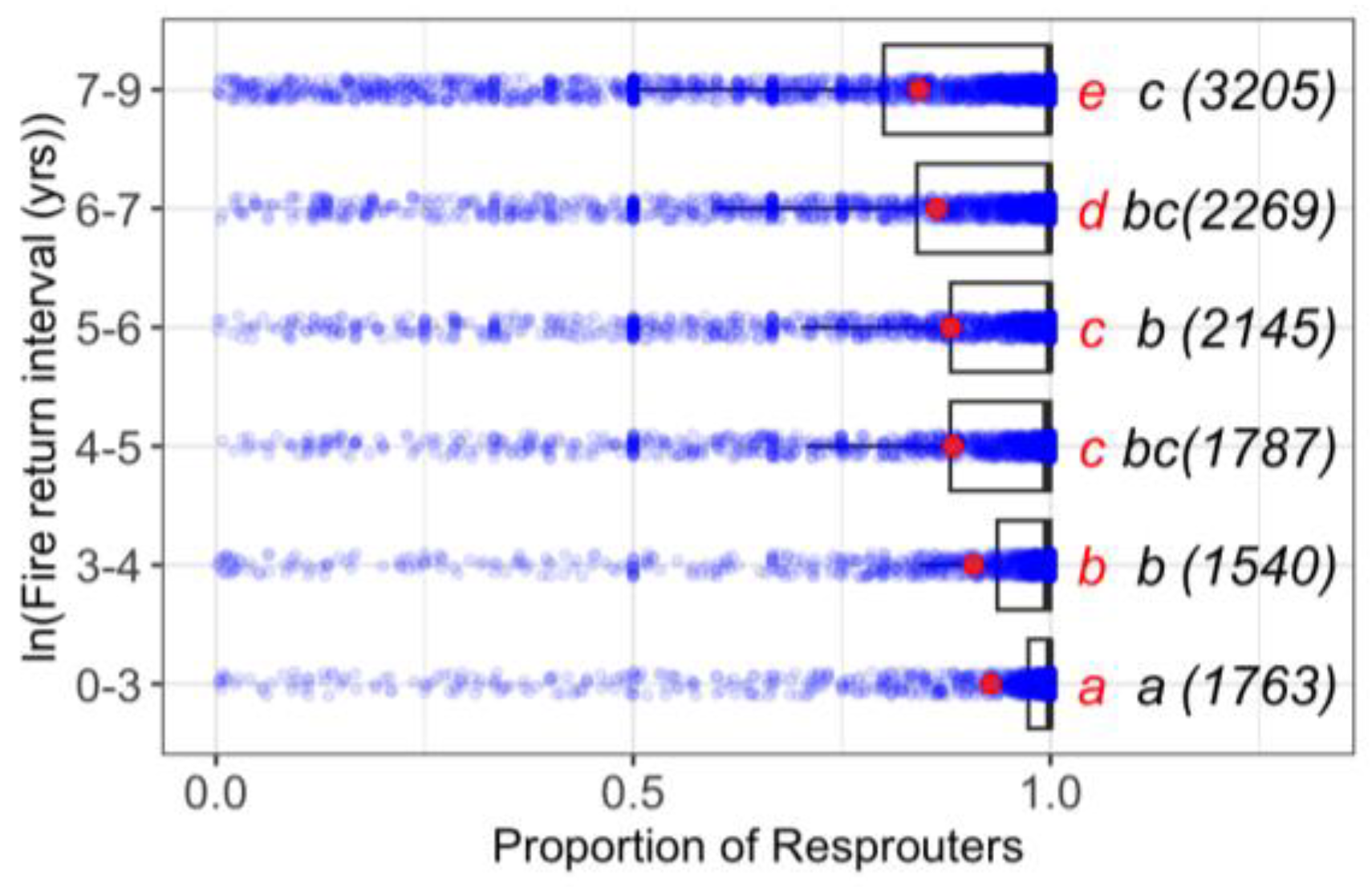
3.1. Relationship between the Fire Return Interval and the Incidence of Resprouting
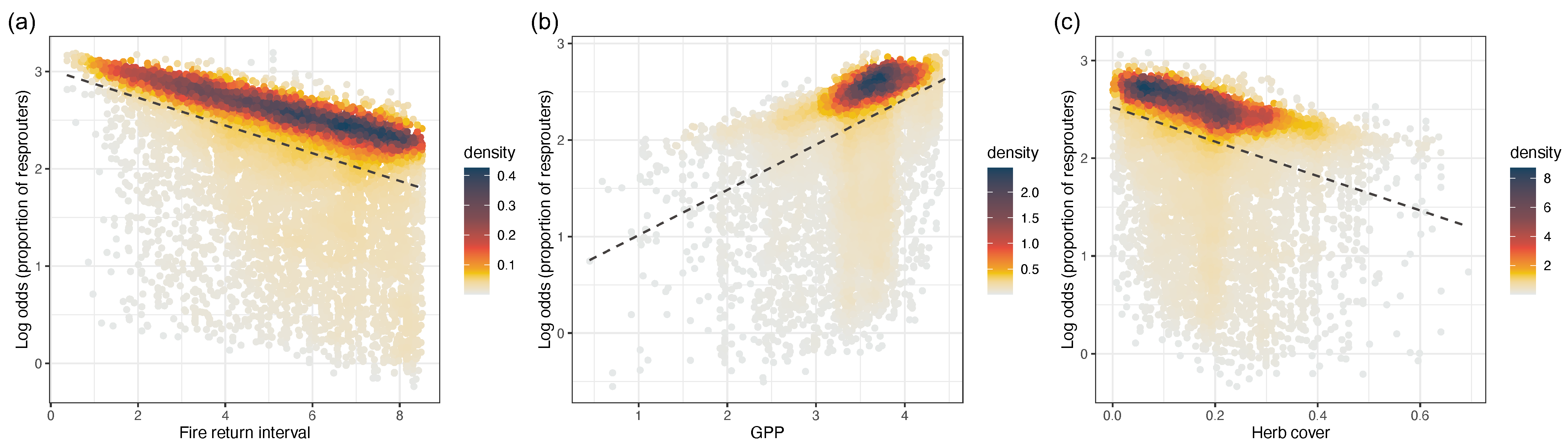
3.2. Relationships between the Incidence of Resprouting, GPP, and Grass Cover
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dupuy, J.-L.; Fargeon, H.; Martin-Stpaul, N.; Pimont, F.; Ruffault, J.; Guijarro, M.; Hernando, C.; Madrigal, J.; Fernandes, P. Climate change impact on future wildfire danger and activity in southern Europe: A review. Ann. For. Sci. 2020, 77, 35. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef]
- Harrison, S.P.; Marlon, J.R.; Bartlein, P.J. Fire in the Earth system. In Changing Climates, Earth Systems and Society; Dodson, J., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 21–48. [Google Scholar]
- Liu, H.; Randerson, J.T.; Lindfors, J.; Chapin, F.S. Changes in the surface energy budget after fire in boreal ecosystems of interior Alaska: An annual perspective. J. Geophys. Res. 2005, 110, D13101. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant Functional Traits in Relation to Fire in Crown-Fire Ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G. Evolutionary Ecology of Fire. Annu. Rev. Ecol. Evol. Syst. 2022, 53, 203–225. [Google Scholar] [CrossRef]
- Lamont, B.B.; He, T.; Yan, Z. Evolutionary history of fire-stimulated resprouting, flowering, seed release and germination. Biol. Rev. Camb. Philos. Soc. 2019, 94, 903–928. [Google Scholar] [CrossRef] [PubMed]
- Enright, N.J.; Fontaine, J.B.; Lamont, B.B.; Miller, B.P.; Westcott, V.C. Resistance and resilience to changing climate and fire regime depend on plant functional traits. J. Ecol. 2014, 102, 1572–1581. [Google Scholar] [CrossRef]
- Harrison, S.P.; Prentice, I.C.; Bloomfield, K.J.; Dong, N.; Forkel, M.; Forrest, M.; Ningthoujam, R.K.; Pellegrini, A.; Shen, Y.; Baudena, M.; et al. Understanding and modelling wildfire regimes: An ecological perspective. Environ. Res. Lett. 2021, 16, 125008. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Epicormic Resprouting in Fire-Prone Ecosystems. Trends Plant Sci. 2017, 22, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Clarke, P.J.; Lawes, M.J.; Murphy, B.P.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.; Enright, N.J.; Fontaine, J.B.; Gosper, C.R.; Radford, I.; et al. A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci. Total Environ. 2015, 534, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Pratt, R.B.; Keeley, J.E.; Jacobsen, A.L.; Ramirez, A.R.; Vilagrosa, A.; Paula, S.; Kaneakua-Pia, I.N.; Davis, S.D. Towards understanding resprouting at the global scale. New Phytol. 2016, 209, 945–954. [Google Scholar] [CrossRef]
- Thonicke, K.; Spessa, A.; Prentice, I.C.; Harrison, S.P.; Dong, L.; Carmona-Moreno, C. The influence of vegetation, fire spread and fire behaviour on biomass burning and trace gas emissions: Results from a process-based model. Biogeosciences 2010, 7, 1991–2011. [Google Scholar] [CrossRef]
- Higgins, S.I.; Bond, W.J.; Trollope, W.S.W. Fire, resprouting and variability: A recipe for grass-tree coexistence in savanna. J. Ecol. 2000, 88, 213–229. [Google Scholar] [CrossRef]
- Kelley, D.I.; Harrison, S.P.; Prentice, I.C. Improved simulation of fire–vegetation interactions in the Land surface Processes and eXchanges dynamic global vegetation model (LPX-Mv1). Geosci. Model Dev. 2014, 7, 2411–2433. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Kattge, J.; Bönisch, G.; Díaz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; Tautenhahn, S.; Werner, G.D.A.; Aakala, T.; Abedi, M.; et al. TRY plant trait database—Enhanced coverage and open access. Glob. Chang. Biol. 2020, 26, 119–188. [Google Scholar] [CrossRef]
- Falster, D.; Gallagher, R.; Wenk, E.H.; Sauquet, H.; Wright, I.J.; Indiarto, D.; Andrew, S.C.; Baxter, C.; Lawson, J.; Allen, S.; et al. AusTraits, a curated plant trait database for the Australian flora. Sci. Data 2021, 8, 254. [Google Scholar] [CrossRef]
- Tavşanoğlu, Ç.; Pausas, J.G. A functional trait database for Mediterranean Basin plants. Sci. Data 2018, 5, 180135. [Google Scholar] [CrossRef]
- Sabatini, F.M.; Lenoir, J.; Hattab, T.; Arnst, E.A.; Chytrý, M.; Dengler, J.; De Ruffray, P.; Hennekens, S.M.; Jandt, U.; Jansen, F.; et al. sPlotOpen—An environmentally balanced, open-access, global dataset of vegetation plots. Glob. Ecol. Biogeogr. 2021, 30, 1740–1764. [Google Scholar] [CrossRef]
- Russell Smith, J.; Gardener, M.R.; Brock, C.; Brennan, K.; Yates, C.P.; Grace, B. Fire persistence traits can be used to predict vegetation response to changing fire regimes at expansive landscape scales—An Australian example. J. Biogeogr. 2012, 39, 1657–1668. [Google Scholar] [CrossRef]
- Giglio, L.; Schroeder, W.; Justice, C.O. The collection 6 MODIS active fire detection algorithm and fire products. Remote Sens. Environ. 2016, 178, 31–41. [Google Scholar] [CrossRef]
- Giglio, L.; Boschetti, L.; Roy, D.; Hoffmann, A.A.; Humber, M.; Hall, J.V. Collection 6 Modis Burned Area Product User’s Guide Version 1.0; NASA EOSDIS Land Processes DAAC: Sioux Falls, SD, USA, 2016. [Google Scholar]
- Lizundia-Loiola, J.; Otón, G.; Ramo, R.; Chuvieco, E. A spatio-temporal active-fire clustering approach for global burned area mapping at 250 m from MODIS data. Remote Sens. Environ. 2020, 236, 111493. [Google Scholar] [CrossRef]
- Otón, G.; Ramo, R.; Lizundia-Loiola, J.; Chuvieco, E. Global Detection of Long-Term (1982–2017) Burned Area with AVHRR-LTDR Data. Remote Sens. 2019, 11, 2079. [Google Scholar] [CrossRef]
- Archibald, S.; Lehmann, C.E.R.; Gómez-Dans, J.L.; Bradstock, R.A. Defining pyromes and global syndromes of fire regimes. Proc. Natl. Acad. Sci. USA 2013, 110, 6442–6447. [Google Scholar] [CrossRef] [PubMed]
- Hantson, S.; Arneth, A.; Harrison, S.P.; Kelley, D.I.; Prentice, I.C.; Rabin, S.S.; Archibald, S.; Mouillot, F.; Arnold, S.R.; Artaxo, P.; et al. The status and challenge of global fire modelling. Biogeosciences 2016, 13, 3359–3375. [Google Scholar] [CrossRef]
- Haas, O.; Prentice, I.C.; Harrison, S.P. Global environmental controls on wildfire burnt area, size, and intensity. Environ. Res. Lett. 2022, 17, 065004. [Google Scholar] [CrossRef]
- Casals, P.; Rios, A.I. Burning intensity and low light availability reduce resprouting ability and vigor of Buxus sempervirens L. after clearing. Sci. Total Environ. 2018, 627, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Meunier, J.; Holoubek, N.S.; Johnson, Y.; Kuhman, T.; Strobel, B. Effects of fire seasonality and intensity on resprouting woody plants in prairie-forest communities. Restor. Ecol. 2021, 29, e13451. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review and suggested usage. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Stocker, B.D.; Wang, H.; Smith, N.G.; Harrison, S.P.; Keenan, T.F.; Sandoval, D.; Davis, T.; Prentice, I.C. P-model v1.0: An optimality-based light use efficiency model for simulating ecosystem gross primary production. Geosci. Model Dev. 2020, 13, 1545–1581. [Google Scholar] [CrossRef]
- Cai, W.; Prentice, I.C. Recent trends in gross primary production and their drivers: Analysis and modelling at flux-site and global scales. Environ. Res. Lett. 2020, 15, 124050. [Google Scholar] [CrossRef]
- Wang, H.; Prentice, I.C.; Keenan, T.F.; Davis, T.W.; Wright, I.J.; Cornwell, W.K.; Evans, B.J.; Peng, C. Towards a universal model for carbon dioxide uptake by plants. Nat. Plants 2017, 3, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Weedon, G.P.; Balsamo, G.; Bellouin, N.; Gomes, S.; Best, M.J.; Viterbo, P. The WFDEI meteorological forcing data set: WATCH Forcing Data methodology applied to ERA-Interim reanalysis data. Water Resour. Res. 2014, 50, 7505–7514. [Google Scholar] [CrossRef]
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (LAI) 3g and fraction of photosynthetically active radiation (FPAR) 3g derived from global inventory modeling and mapping studies (GIMMS) normalized difference vegetation index (NDVI3g) for the period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar]
- Still, C.J.; Berry, J.A.; Collatz, G.J.; Defries, R.S. Global distribution of C3 and C4 vegetation: Carbon cycle implications. Glob. Biogeochem. Cycles 2003, 17, 6-1–6-14. [Google Scholar] [CrossRef]
- European Space Agency (ESA). Land Cover CCI Product User Guide Version 2; European Space Agency: Paris, France, 2017. [Google Scholar]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman and Hall: New York, NY, USA, 1989. [Google Scholar]
- Nelder, J.A.; Wedderburn, R.W. Generalized linear models. J. R. Stat. Soc. Ser. A (Gen.) 1972, 135, 370–384. [Google Scholar] [CrossRef]
- O’Brien, R.M. A Caution Regarding Rules of Thumb for Variance Inflation Factors. Qual. Quant. 2007, 41, 673–690. [Google Scholar] [CrossRef]
- Larsen, W.A.; McCleary, S.J. The use of partial residual plots in regression analysis. Technometrics 1972, 14, 781–790. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Moris, J.V.; Berretti, R.; Bono, A.; Sino, R.; Minotta, G.; Garbarino, M.; Motta, R.; Vacchiano, G.; Maringer, J.; Conedera, M.; et al. Resprouting in European beech confers resilience to high-frequency fire. For. Int. J. For. Res. 2022, cpac018. [Google Scholar] [CrossRef]
- Fairman, T.A.; Bennett, L.T.; Nitschke, C.R. Short-interval wildfires increase likelihood of resprouting failure in fire-tolerant trees. J. Environ. Manag. 2019, 231, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Cury, R.T.d.S.; Balch, J.K.; Brando, P.M.; Andrade, R.B.; Scervino, R.P.; Torezan, J.M.D. Higher fire frequency impaired woody species regeneration in a south-eastern Amazonian forest. J. Trop. Ecol. 2020, 36, 190–198. [Google Scholar] [CrossRef]
- Thomsen, A.M.; Ooi, M.K.J. Shifting season of fire and its interaction with fire severity: Impacts on reproductive effort in resprouting plants. Ecol. Evol. 2022, 12, e8717. [Google Scholar] [CrossRef] [PubMed]
- Schutz, A.E.N.; Bond, W.J.; Cramer, M.D. Juggling carbon: Allocation patterns of a dominant tree in a fire-prone savanna. Oecologia 2009, 160, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Lawes, M.J.; Crisp, M.D.; Clarke, P.J.; Murphy, B.P.; Midgley, J.J.; Russell-Smith, J.; Nano, C.E.M.; Bradstock, R.A.; Enright, N.J.; Fontaine, J.B.; et al. Appraising widespread resprouting but variable levels of postfire seeding in Australian ecosystems: The effect of phylogeny, fire regime and productivity. Aust. J. Bot. 2022, 70, 114–130. [Google Scholar] [CrossRef]
- Vesk, P.A.; Westoby, M. Sprouting Ability across Diverse Disturbances and Vegetation Types Worldwide. J. Ecol. 2004, 92, 310–320. [Google Scholar] [CrossRef]
- Nicolle, D. A classification and census of regenerative strategies in the eucalypts (Angophora, Corymbia and Eucalyptus—Myrtaceae), with special reference to the obligate seeders. Aust. J. Bot. 2006, 54, 391–407. [Google Scholar] [CrossRef]
- Hartmann, H.; Bahn, M.; Carbone, M.; Richardson, A.D. Plant carbon allocation in a changing world—Challenges and progress: Introduction to a Virtual Issue on carbon allocation. New Phytol. 2020, 227, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.; Bartlein, P. Records from the Past, Lessons for the Future: What the Palaeorecord Implies about Mechanisms of Global Change. In The Future of the World’s Climate; Elsevier: Amsterdam, The Netherlands, 2012; pp. 403–436. [Google Scholar]
- Fritz, S.C. The climate of the Holocene and its landscape and biotic impacts. Tellus B Chem. Phys. Meteorol. 2013, 65, 20602. [Google Scholar] [CrossRef]
- Kaufman, D.; Mckay, N.; Routson, C.; Erb, M.; Dätwyler, C.; Sommer, P.S.; Heiri, O.; Davis, B. Holocene global mean surface temperature, a multi-method reconstruction approach. Sci. Data 2020, 7, 201. [Google Scholar] [CrossRef] [PubMed]
- Daniau, A.L.; Bartlein, P.J.; Harrison, S.P.; Prentice, I.C.; Brewer, S.; Friedlingstein, P.; Harrison-Prentice, T.I.; Inoue, J.; Izumi, K.; Marlon, J.R.; et al. Predictability of biomass burning in response to climate changes. Glob. Biogeochem. Cycles 2012, 26, GB4007. [Google Scholar] [CrossRef]
- Marlon, J.R.; Bartlein, P.J.; Daniau, A.-L.; Harrison, S.P.; Maezumi, S.Y.; Power, M.J.; Tinner, W.; Vanniére, B. Global biomass burning: A synthesis and review of Holocene paleofire records and their controls. Quat. Sci. Rev. 2013, 65, 5–25. [Google Scholar] [CrossRef]
- Cracknell, A.P. Advanced Very High Resolution Radiometer AVHRR; CRC Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Young, N.E.; Anderson, R.S.; Chignell, S.M.; Vorster, A.G.; Lawrence, R.; Evangelista, P.H. A survival guide to Landsat preprocessing. Ecology 2017, 98, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Wulder, M.A.; Loveland, T.R.; Roy, D.P.; Crawford, C.J.; Masek, J.G.; Woodcock, C.E.; Allen, R.G.; Anderson, M.C.; Belward, A.S.; Cohen, W.B. Current status of Landsat program, science, and applications. Remote Sens. Environ. 2019, 225, 127–147. [Google Scholar] [CrossRef]
- Niklasson, M.; Granström, A. Numbers and sizes of fires: Long-term spatially explicit fire history in a Swedish boreal landscape. Ecology 2000, 81, 1484–1499. [Google Scholar] [CrossRef]
- Stambaugh, M.C.; Marschall, J.M.; Abadir, E.R.; Jones, B.C.; Brose, P.H.; Dey, D.C.; Guyette, R.P. Wave of fire: An anthropogenic signal in historical fire regimes across central Pennsylvania, USA. Ecosphere 2018, 9, e02222. [Google Scholar] [CrossRef]
- Harrison, S.P.; Villegas-Diaz, R.; Cruz-Silva, E.; Gallagher, D.; Kesner, D.; Lincoln, P.; Shen, Y.; Sweeney, L.; Colombaroli, D.; Ali, A.; et al. The Reading Palaeofire Database: An expanded global resource to document changes in fire regimes from sedimentary charcoal records. Earth Syst. Sci. Data 2022, 14, 1109–1124. [Google Scholar] [CrossRef]
- Moore, N.A.; Camac, J.S.; Morgan, J.W. Effects of drought and fire on resprouting capacity of 52 temperate Australian perennial native grasses. New Phytol. 2019, 221, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Viedma, O.; Meliá, J.; Segarra, D.; Garcia-Haro, J. Modeling rates of ecosystem recovery after fires by using landsat TM data. Remote Sens. Environ. 1997, 61, 383–398. [Google Scholar] [CrossRef]
- Zeppel, M.J.B.; Harrison, S.P.; Adams, H.D.; Kelley, D.I.; Li, G.; Tissue, D.T.; Dawson, T.E.; Fensham, R.; Medlyn, B.E.; Palmer, A.; et al. Drought and resprouting plants. New Phytol. 2015, 206, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Cragg, J.G.; Uhler, R.S. The Demand for Automobiles. Can J. Econ. 1970, 3, 386–406. [Google Scholar] [CrossRef]
- Shen, Y. Code and Data for Resprouting Analysis. 2023. Available online: https://figshare.com/articles/software/Code_and_data_for_resprouting_analysis/21899487/1 (accessed on 19 April 2023).
- Cai, W. Global Monthly Gross Primary Production. 2023. Available online: https://zenodo.org/record/7513533#.ZD-LlezMI6E (accessed on 19 April 2023). [CrossRef]
- Pausas, J.G.; Tavşanoğlu, Ç. BROT Plant Functional Trait Database: Data File. 2018. Available online: https://springernature.figshare.com/articles/dataset/BROT_plant_functional_trait_database_Data_file/5280868/1 (accessed on 19 April 2023). [CrossRef]
- Chuvieco, E.; Pettinari, M.L.; Otón, G. ESA Fire Climate Change Initiative (Fire_cci): AVHRR-LTDR Burned Area Grid Product, Version 1.1; European Space Agency: Paris, France, 2020. [Google Scholar] [CrossRef]
- Haas, O. Scripts and Input Files. 2023. Available online: https://figshare.com/articles/dataset/Scripts_and_input_files/19071044/1 (accessed on 19 April 2023). [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Woody Species | R+ | R− | R? | Percentage of Known Species | |
|---|---|---|---|---|---|
| Australia | 1890 | 969 (0) | 259 (1) | 661 | 65.03% |
| Europe | 913 | 194 (83) | 32 (38) | 566 | 38.01% |
| Estimate Coefficient | Standard Error | z-Value | VIF | |
|---|---|---|---|---|
| (Intercept) | 1.58 | 0.19 | 8.39 *** | |
| Fire return interval | −0.14 | 0.02 | −9.43 *** | 1.01 |
| GPP | 0.45 | 0.05 | 9.73 *** | 1.02 |
| Grass cover | −1.70 | 0.23 | −7.37 *** | 1.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Cai, W.; Prentice, I.C.; Harrison, S.P. Community Abundance of Resprouting in Woody Plants Reflects Fire Return Time, Intensity, and Type. Forests 2023, 14, 878. https://doi.org/10.3390/f14050878
Shen Y, Cai W, Prentice IC, Harrison SP. Community Abundance of Resprouting in Woody Plants Reflects Fire Return Time, Intensity, and Type. Forests. 2023; 14(5):878. https://doi.org/10.3390/f14050878
Chicago/Turabian StyleShen, Yicheng, Wenjia Cai, I. Colin Prentice, and Sandy P. Harrison. 2023. "Community Abundance of Resprouting in Woody Plants Reflects Fire Return Time, Intensity, and Type" Forests 14, no. 5: 878. https://doi.org/10.3390/f14050878
APA StyleShen, Y., Cai, W., Prentice, I. C., & Harrison, S. P. (2023). Community Abundance of Resprouting in Woody Plants Reflects Fire Return Time, Intensity, and Type. Forests, 14(5), 878. https://doi.org/10.3390/f14050878