
Plant Traits of Tilia tomentosa Moench, Fraxinus excelsior L., and Pinus nigra J.F.Arnold as a Proxy of Urbanization


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Urbanization Intensity Classification
2.3. Plant Traits
2.3.1. Concentrations of Potentially Toxic Elements (PTEs) in Tree Leaves/Needles
2.3.2. Leaf Traits
2.3.3. Tree Physiology
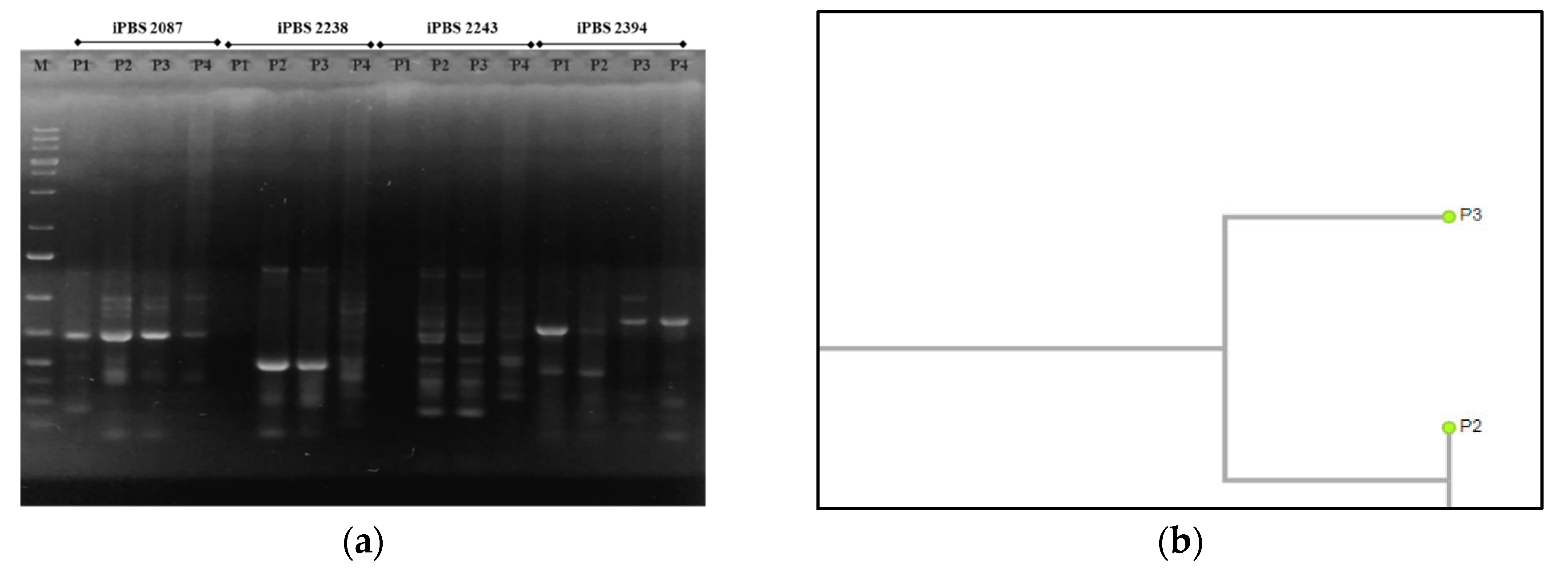
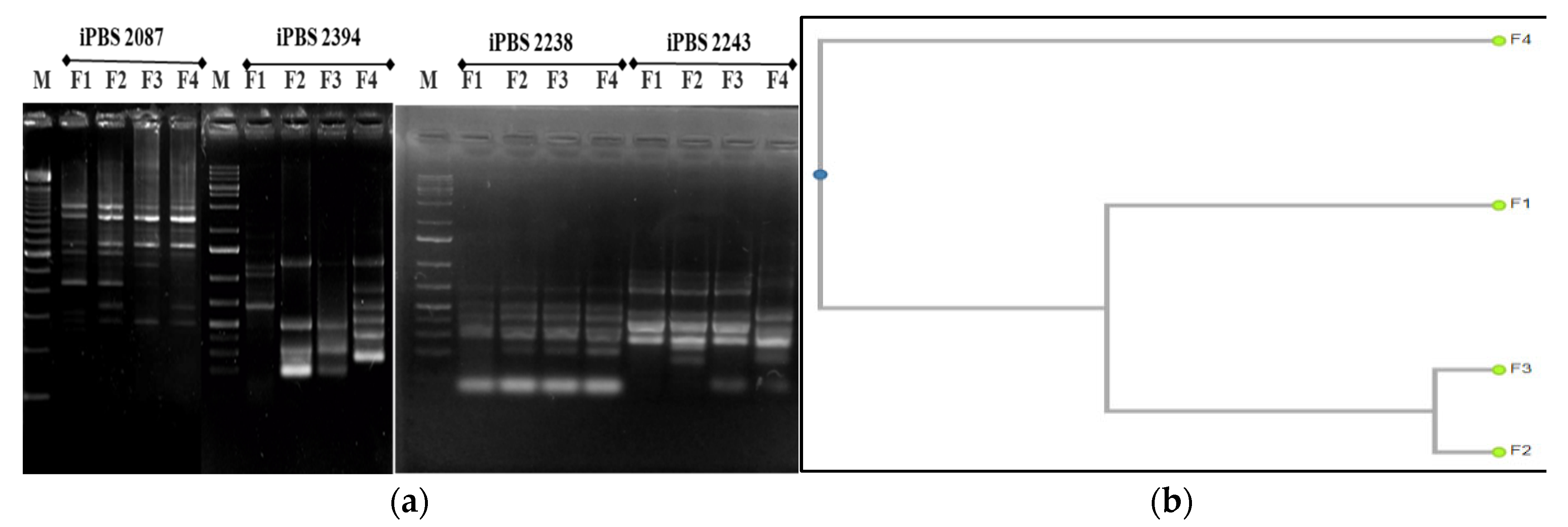
2.3.4. Molecular Markers
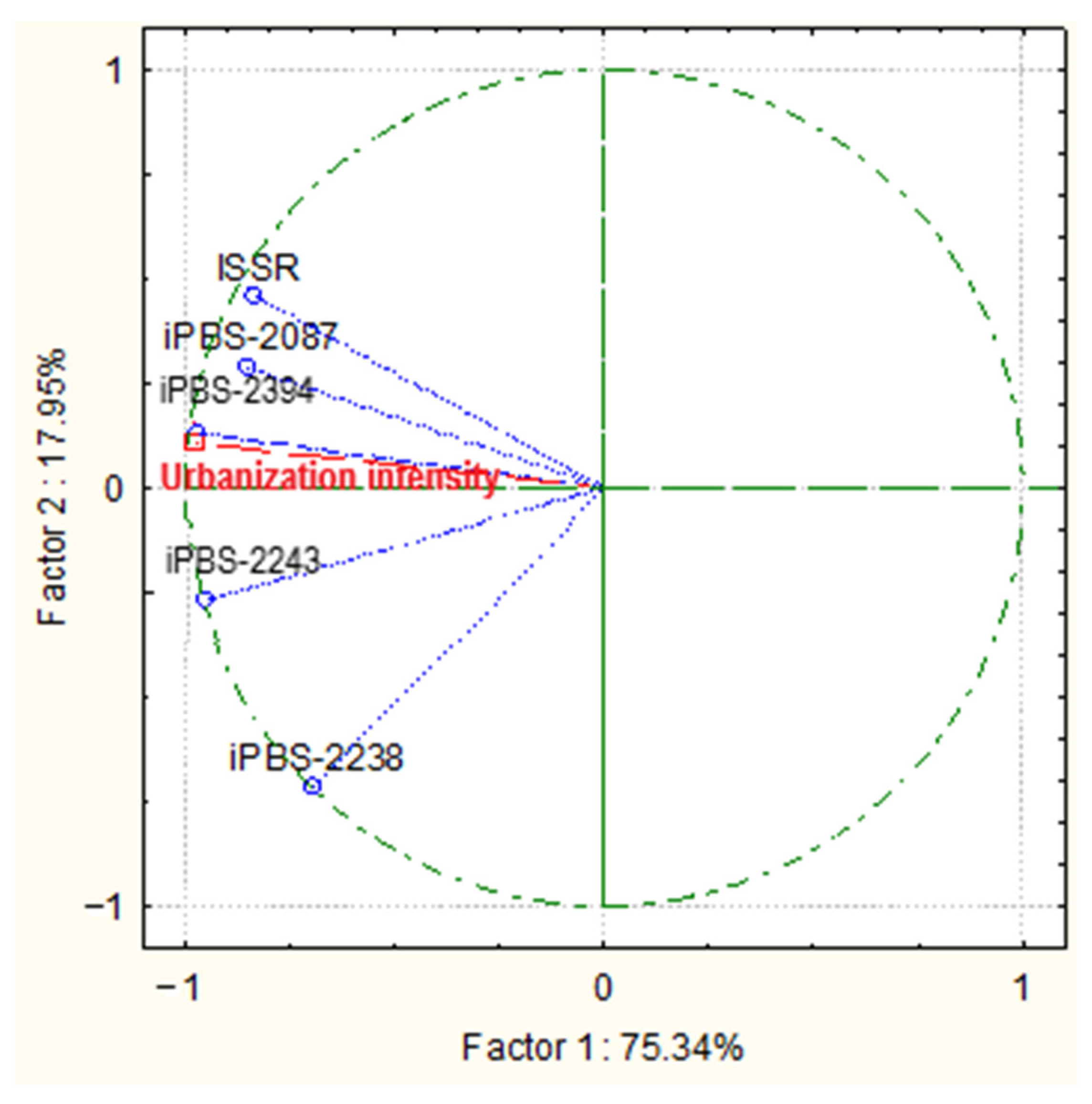
2.4. Data Processing
3. Results
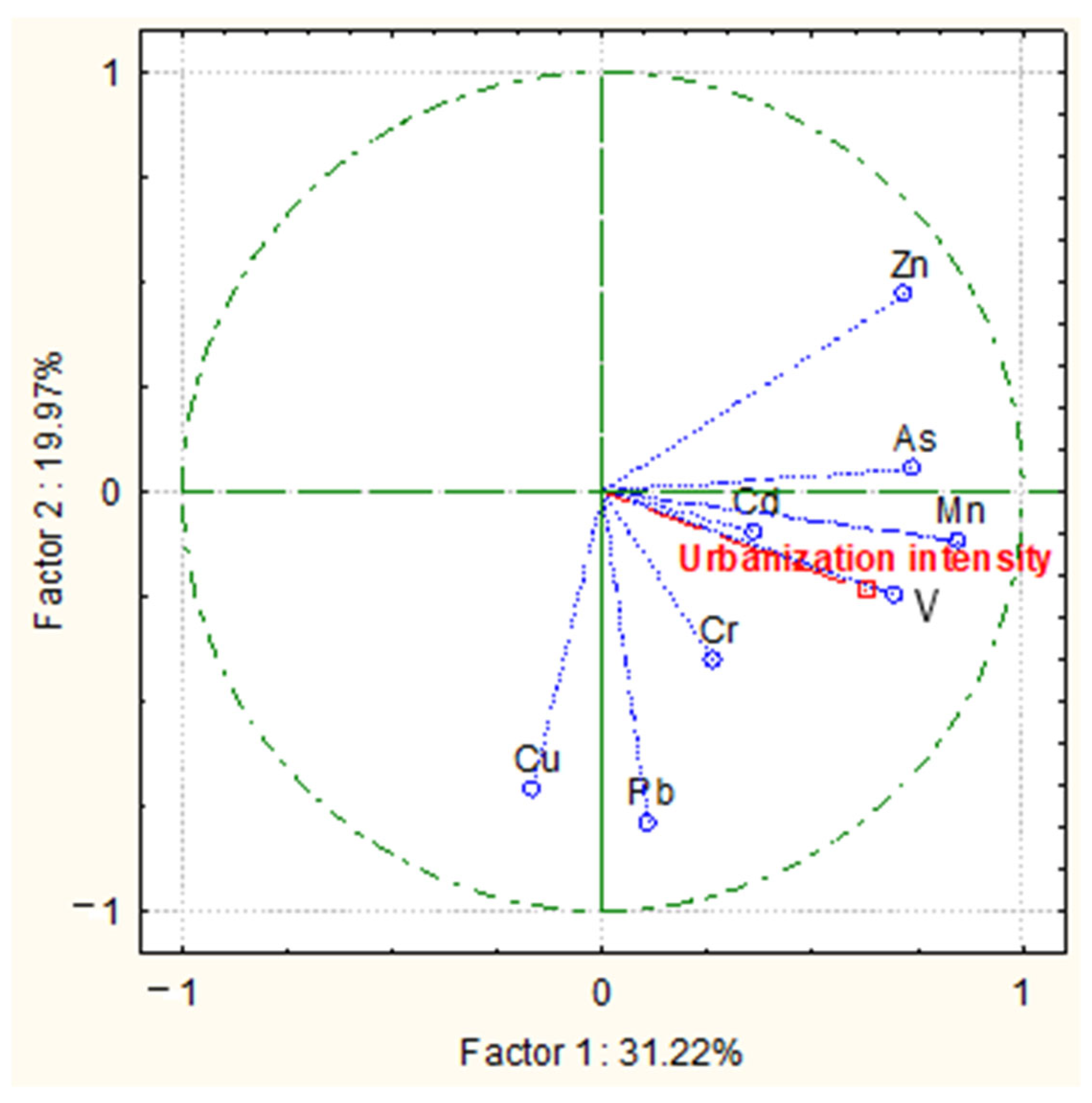
3.1. Concentrations of PTEs in Tree Leaves/Needles
- Plot 1—Mn > Zn > Cu > Pb > Cr > As > V > Cd
- Plot 2—Mn > Zn > Cu > Pb > As > Cr > V > Cd
- Plot 3—Mn > Zn > Cu > Pb > V > As > Cr > Cd
- Plot 4—Mn > Zn > Cu > Pb > Cr > V > As > Cd.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Plant Location | As mg/kg | Cd mg/kg | Cr mg/kg | Cu mg/kg | Mn mg/kg | Pb mg/kg | V mg/kg | Zn mg/kg |
|---|---|---|---|---|---|---|---|---|---|
| T. tomentosa | Plot 1 | 0.16 | 0.047 | 0.76 | 6.4 | 43 | 2.6 | 0.34 | 20 |
| Plot 2 | 0.27 | 0.043 | 0.35 | 15.0 | 28 | 6.5 | 0.35 | 19 | |
| Plot 3 | 0.30 | 0.058 | 0.36 | 6.0 | 61 | 2.7 | 0.47 | 23 | |
| Plot 4 | 0.15 | 0.041 | 0.33 | 5.1 | 28 | 2.6 | 0.30 | 17 | |
| F. excelsior | Plot 1 | 0.48 | 0.020 | 0.55 | 5.2 | 19 | 1.6 | 0.33 | 21 |
| Plot 2 | 0.40 | 0.210 | 0.35 | 9.3 | 40 | 2.0 | 0.24 | 22 | |
| Plot 3 | 0.15 | 0.022 | 0.25 | 25.0 | 25 | 1.9 | 0.33 | 16 | |
| Plot 4 | 0.09 | 0.018 | 0.22 | 4.0 | 17 | 1.1 | 0.22 | 15 | |
| P. nigra | Plot 1 | 0.25 | 0.049 | 0.74 | 5.2 | 28 | 4.2 | 0.16 | 23 |
| Plot 2 | 0.43 | 0.028 | 0.38 | 3.3 | 28 | 2.3 | 0.28 | 23 | |
| Plot 3 | 0.39 | 0.034 | 0.23 | 4.1 | 46 | 1.3 | 0.42 | 36 | |
| Plot 4 | 0.05 | 0.027 | 0.10 | 2.9 | 17 | 1.1 | 0.14 | 22 |
3.2. Leaf Traits
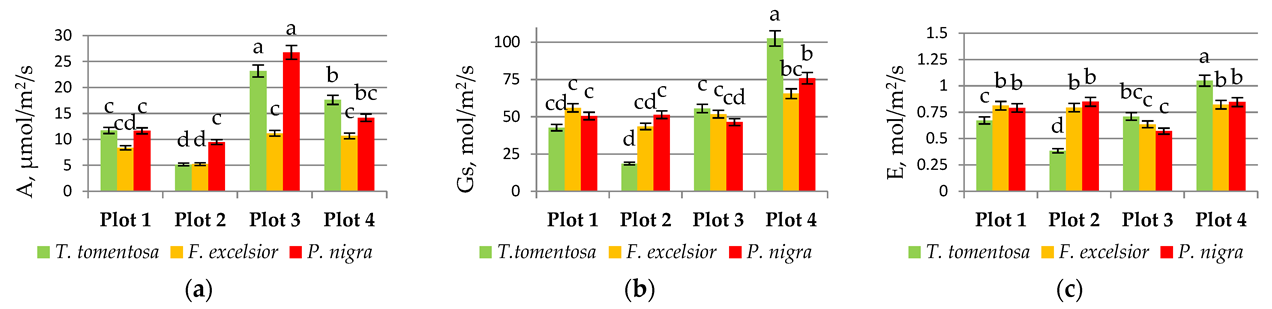
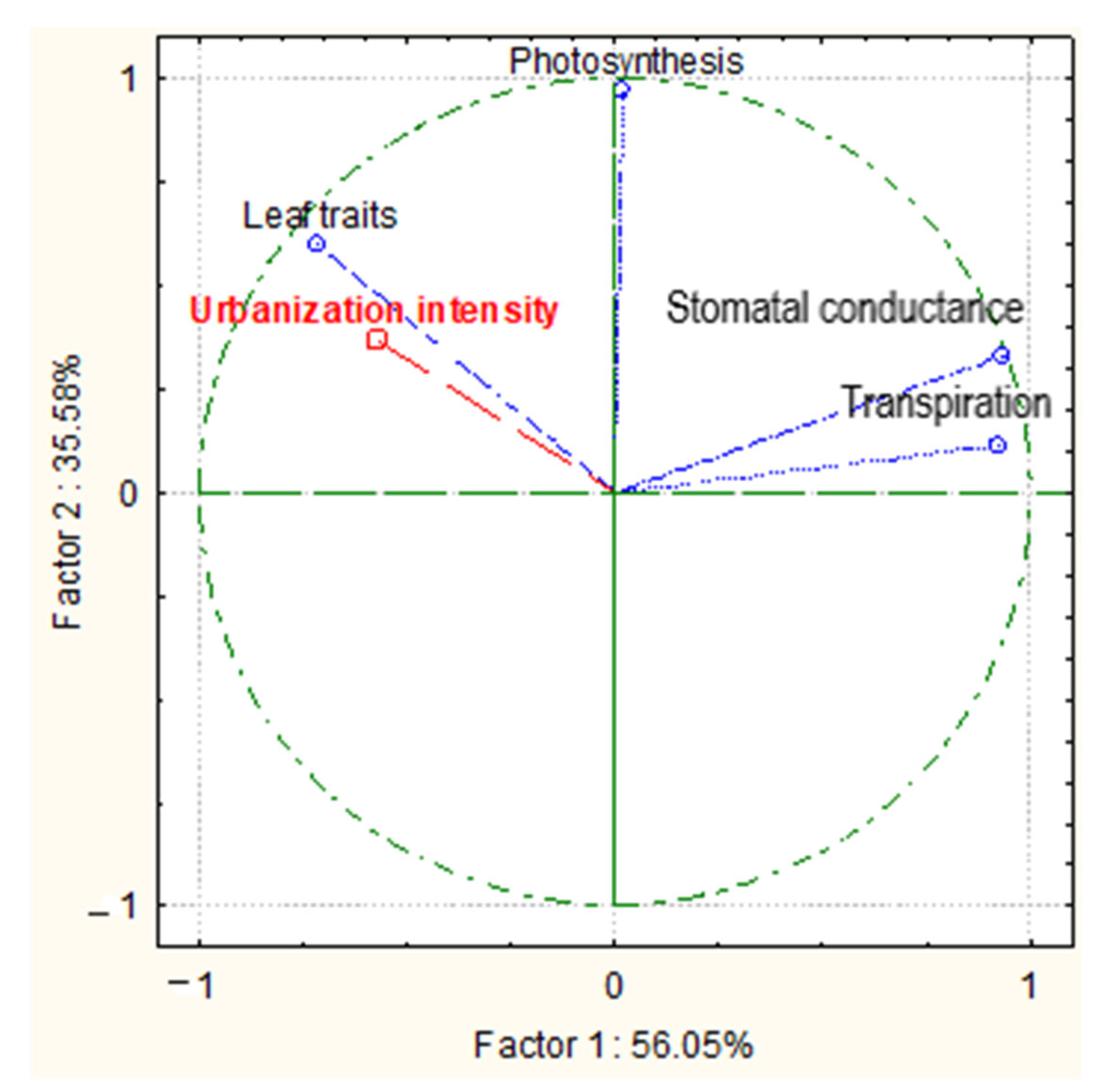
3.3. Physiological Traits
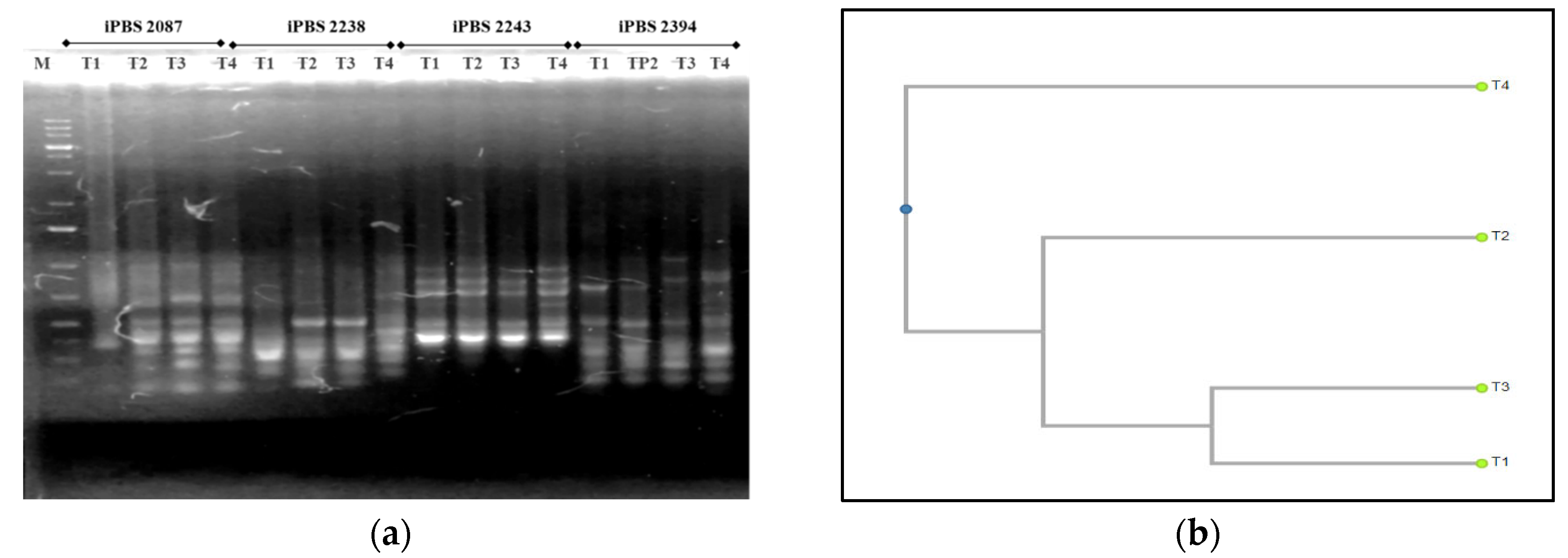
3.4. Molecular Traits
3.4.1. ISSR—PCR Analysis
3.4.2. iPBS—PCR Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brune, M. Urban Trees under Climate Change. Potential Impacts of Dry Spells and Heat Waves in Three German Regions in the 2050s; Report No. 24; Climate Service Center Germany: Hamburg, Germany, 2016. [Google Scholar]
- Locosselli, G.M.; de Camargo, E.P.; Moreira, T.C.L.; Todesco, E.; Andrade, M.F.; de André, C.D.S.; de André, P.A.; Singer, J.M.; Ferreira, L.S.; Saldiva, P.H.N.; et al. The role of air pollution and climate on the growth of urban trees. Sci. Total Environ. 2019, 666, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Schütze, G.; Perkins, D.; Rötzer, T.; Caldentey, J.; Koike, T.; Con, T.V.; et al. Climate change accelerates growth of urban trees in metropolises worldwide. Sci. Rep. 2017, 7, 15403. [Google Scholar] [CrossRef] [PubMed]
- Martínez-López, S.; Martínez-Sánchez, M.J.; Pérez-Sirvent, C.; Bech, J.; Martínez, M.d.C.G.; García-Fernandez, A.J. Screening of wild plants for use in the phytoremediation of mining-influenced soils containing arsenic in semiarid environments. J. Soils Sediments 2014, 14, 794–809. [Google Scholar] [CrossRef]
- Ilyas, M.; Liu, Y.Y.; Shah, S.; Ali, A.; Khan, A.H.; Zaman, F.; Yucui, Z.; Saud, S.; Adnan, M.; Ahmed, N.; et al. Adaptation of functional traits and their plasticity of three ornamental trees growing in urban environment. Sci. Hortic. 2021, 286, 110248. [Google Scholar] [CrossRef]
- Bierne, N.; Roze, D.; Welch, J.J. Pervasive selection or is it…? Why are FST outliers sometimes so frequent? Mol. Ecol. 2013, 22, 2061–2064. [Google Scholar]
- Garnier, E.; Navas, M.-L. A trait-based approach to comparative functional plant ecology: Concepts, methods and applications for agroecology. A review. Agron. Sustain. Dev. 2012, 32, 365–399. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Xu, L.; Chen, Z.; He, N. Variation in leaf morphological, stomatal, and anatomical traits and their relationships in temperate and subtropical forests. Sci. Rep. 2019, 9, 5803. [Google Scholar] [CrossRef]
- Chi, D.; Meerbeek, K.; Yu, K.; Degerickx, J.; Somers, B. Foliar optical traits capture physiological and phenological leaf plasticity in Tilia×euchlora in the urban environment. Sci. Total Environ. 2022, 805, 150219. [Google Scholar] [CrossRef]
- Mitu, K.J.; Islam, M.A.; Biswas, P.; Marzia, S.; Ali, M.A. Effects of different environmental pollutants on the anatomical features of roadside plants. Progress. Agric. 2019, 30, 344–351. [Google Scholar] [CrossRef]
- Andrianjara, I.; Bordenave-Jacquemin, M.; Roy, V.; Cabassa, C.; Federici, P.; Carmignac, D.; Marcangeli, Y.; Rouhan, G.; Renard, M.; Nold, F.; et al. Urban tree management: Diversity of Tilia genus in streets and parks of Paris based on morphological and genetic characteristics. Urban For. Urban Green. 2021, 66, 127382. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Chen, Y.; Berlyn, G.P. Photosynthetic parameters of urban greening trees growing on paved land. iForest—Biogeosci. For. 2019, 12, 403–410. [Google Scholar] [CrossRef]
- Rötzer, T.; Rahman, M.A.; Moser-Reischl, A.; Pauleit, S.; Pretzsch, H. Process based simulation of tree growth and ecosystem services of urban trees under present and future climate conditions. Sci. Total Environ. 2019, 676, 651–664. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, F.; Li, F.; Jin, G. Coordination of intra and inter-species leaf traits according to leaf phenology and plant age for three temperate broadleaf species with different shade tolerances. For. Ecol. Manag. 2019, 434, 63–75. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Shanker, A.; Venkateswarlu, B. (Eds.) Abiotic Stress in Plants. Mechanisms and Adaptations; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- McCarthy, D.M.; Mason-Gamer, R.J. Morphological variation in North American Tilia and its value in species delineation. Int. J. Plant Sci. 2020, 181, 175–195. [Google Scholar] [CrossRef]
- Phuekvilai, P.; Wolff, K. Characterization of microsatellite loci in Tilia platyphyllos (Malvaceae) and cross-amplification in related species. Appl. Plant Sci. 2013, 1, 1200386. [Google Scholar] [CrossRef] [PubMed]
- Gozukirmizi, N.; Yilmaz, S.; Marakli, S.; Temel, A. Retrotransposon-Based Molecular Markers; Tools for Variation Analysis in Plants. In Applications of Molecular Markers in Plant Genome Analysis and Breeding; Taški-Ajduković, K., Ed.; IntechOpen: Kolkata, India, 2015; pp. 19–45. [Google Scholar]
- Petrova, S. Efficiency of Pinus nigra J.F. Arnold in removing pollutants from urban environment (Plovdiv, Bulgaria). Environ. Sci. Pollut. Res. 2020, 27, 39490–39506. [Google Scholar] [CrossRef]
- Petrova, S.; Velcheva, I.; Nikolov, B.; Vasileva, T.; Bivolarski, V. Antioxidant Responses and Adaptation Mechanisms of Tilia tomentosa Moench, Fraxinus excelsior L. and Pinus nigra J. F. Arnold towards Urban Air Pollution. Forests 2022, 13, 1689. [Google Scholar] [CrossRef]
- Alue, B.A.; Salleh Hudin, N.; Mohamed, F.; Mat Said, Z.; Ismail, K. Plant Diversity along an Urbanization Gradient of a Tropical City. Diversity 2022, 14, 1024. [Google Scholar] [CrossRef]
- Khavaninzadeh, A.R.; Veroustraete, F.; Buytaert, J.A.N.; Samson, R. Leaf injury symptoms of Tilia sp. as an indicator of urban habitat quality. Ecol. Indic. 2014, 41, 58–64. [Google Scholar] [CrossRef]
- Petrova, S.; Velcheva, I.; Nikolov, B.; Marinov-Serafimov, P.; Golubinova, I.; Popov, V.; Valcheva, E.; Todorova, K. Assessment of the adaptation of some tree species to the urban environment (Plovdiv, Bulgaria). Comptes Rendus de L’Academie Bulgare des Sciences 2019, 72, 1676–1683. [Google Scholar]
- Haliloğlu, K.; Türkoğlu, A.; Öztürk, H.I.; Özkan, G.; Elkoca, E.; Poczai, P. iPBS-Retrotransposon Markers in the Analysis of Genetic Diversity among Common Bean (Phaseolus vulgaris L.) Germplasm from Türkiye. Genes 2022, 13, 1147. [Google Scholar] [CrossRef] [PubMed]
- Akçali Giachino, R.R. Investigation of the genetic variation of anise (Pimpinella anisum L.) using RAPD and ISSR markers. Genet. Resour. Crop Evol. 2020, 67, 763–780. [Google Scholar] [CrossRef]
- Silva, S.V.P.; Procunier, J.D. Genomic fingerprinting using the PCR-random amplified polymorphic DNA technique. In Methods of Gene Technology; Dale, J.W., Sanders, P.G., Eds.; JAI Press: Middlesex, UK, 1994; Volume 2, pp. 319–336. [Google Scholar]
- Denduangboripant, J.; Setaphan, S.; Suwanprasart, W.; Panha, S. Determination of Local Tobacco Cultivars Using ISSR Molecular Marker. Chiang Mai J. Sci. 2010, 37, 293–303. [Google Scholar]
- Kalendar, R.; Antonius, K.; Smykal, P.; Schulman, A.H. iPBS: A universal method for DNA fingerprinting and retrotransposon isolation. Theor. Appl. Genet. 2010, 121, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- STATISTICA (Data Analysis Software System), version 12; Computer Software; StatSoft Inc.: Tulsa, OK, USA, 2004.
- Ermakov, V.; Perelomov, L.; Khushvakhtova, S.; Tyutikov, S.; Danilova, V.; Safonov, V. Biogeochemical assessment of the urban area in Moscow. Environ. Monit. Assess. 2017, 189, 641. [Google Scholar] [CrossRef]
- Pan, R.; Wang, X.; Li, N. Plant Physiology; Higher Education Press: Beijing, China, 2012; pp. 69–119. [Google Scholar]
- Takagi, M.; Gyokusen, K. Light and atmospheric pollution affect photosynthesis of street trees in urban environments. Urban For. Urban Green. 2004, 2, 167–171. [Google Scholar] [CrossRef]
- Lenz, K.E.; Host, G.E.; Roskoski, K.; Noormets, A.; Sober, A.; Karnosky, D.F. Analysis of a Farquhar-von Caemmerer-Berry leaf-level photosynthetic rate model for Populus tremuloides in the context of modeling and measurement limitations. Environ. Pollut. 2010, 158, 1015–1022. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Wang, X. Impact of land pavement on photosynthetic characteristics of common greening trees in Beijing, China. Chin. J. Appl. Ecol. 2017, 28, 2423–2430. (In Chinese) [Google Scholar]
- Brandon, R.N. Adaptation and Environment; Princeton University Press: Princeton, NJ, USA, 1990. [Google Scholar]
- Ahmad, P.; Wani, M.R. (Eds.) Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment; Springer: New York, NY, USA, 2014; Volume 2. [Google Scholar]










| Motor Traffic (Car Number per 10 min) | Built-Up Area (%) | Urbanization Intensity | Experimental Plot |
|---|---|---|---|
| 0–10 | 0–10 | Low | Plot 4 |
| 10–50 | 10–40 | Medium | Plot 2 |
| 50–150 | 40–70 | High | Plot 1 |
| >150 | >70 | Very high | Plot 3 |
| Plant Location | Plant Species | As mg/kg | Cd mg/kg | Cr mg/kg | Cu mg/kg | Mn mg/kg | Pb mg/kg | V mg/kg | Zn mg/kg |
|---|---|---|---|---|---|---|---|---|---|
| Nursery | T. tomentosa | 0.18 | 0.08 | 0.64 | 7.4 | 63 | 3.9 | 0.46 | 18 |
| F. excelsior | 0.32 | 0.04 | 0.29 | 11.2 | 43 | 1.4 | 0.20 | 18 | |
| P. nigra | 0.07 | 0.09 | 0.34 | 3.2 | 8 | 3.3 | 0.23 | 26 |
| Plant Species | Plant Location | As mg/kg | Cd mg/kg | Cr mg/kg | Cu mg/kg | Mn mg/kg | Pb mg/kg | V mg/kg | Zn mg/kg |
|---|---|---|---|---|---|---|---|---|---|
| T. tomentosa | Plot 1 | 0.4 | 0.13 | 0.34 | 5.5 | 60 | 6.1 | 0.31 | 17 |
| Plot 2 | 0.2 | 0.16 | 0.53 | 4.5 | 100 | 4.8 | 0.30 | 14 | |
| Plot 3 | 0.4 | 0.17 | 0.66 | 7.1 | 115 | 6.5 | 0.48 | 16 | |
| Plot 4 | 0.2 | 0.14 | 0.37 | 5.4 | 82 | 3.7 | 0.23 | 14 | |
| F. excelsior | Plot 1 | 0.6 | 0.09 | 0.34 | 19.4 | 55 | 2.1 | 0.18 | 17 |
| Plot 2 | 0.4 | 0.14 | 0.25 | 6.3 | 39 | 4.1 | 0.18 | 12 | |
| Plot 3 | 0.8 | 0.13 | 0.34 | 8.8 | 58 | 3.6 | 0.21 | 18 | |
| Plot 4 | 0.4 | 0.06 | 0.25 | 5.1 | 36 | 2.9 | 0.14 | 9 | |
| P. nigra | Plot 1 | 0.29 | 0.13 | 0.48 | 3.2 | 8 | 3.4 | 0.28 | 23 |
| Plot 2 | 0.10 | 0.14 | 0.58 | 2.4 | 10 | 5.7 | 0.20 | 22 | |
| Plot 3 | 0.32 | 0.11 | 0.54 | 3.5 | 7 | 5.0 | 0.27 | 29 | |
| Plot 4 | 0.10 | 0.07 | 0.40 | 2.2 | 5 | 2.7 | 0.18 | 18 |
| Plant Species | Plant Location | Leaf Injuries (Percentage) | Class Injuries (1—Low; 5—Very High) |
|---|---|---|---|
| T. tomentosa | Plot 1 | 5%–10% | 2 |
| Plot 2 | 20%–25% | 2 | |
| Plot 3 | 40%–45% | 3 | |
| Plot 4 | 0 | 1 | |
| F. excelsior | Plot 1 | 0 | 1 |
| Plot 2 | 10%–15% | 2 | |
| Plot 3 | 0 | 1 | |
| Plot 4 | 0 | 1 | |
| P. nigra | Plot 1 | 10%–15% | 2 |
| Plot 2 | 10%–15% | 2 | |
| Plot 3 | 25%–30% | 3 | |
| Plot 4 | 1%–3% | 1 |
| iPBS Primers | Sequence of Primers (5′→3′) [9] | Tree Species | AP | PA | % PA | T | Fragment Size (bp) | P | % P |
|---|---|---|---|---|---|---|---|---|---|
| iPBS—2087 | GCA ATG GAA CCA | T. tomentosa | 4 | 4 | 100 | 13 | 75–1000 | 12 | 92.3 |
| P. nigra | 4 | 4 | 100 | 9 | 75–1300 | 7 | 77.8 | ||
| F. excelsior | 4 | 4 | 100 | 9 | 75–1300 | 7 | 77.8 | ||
| iPBS—2238 | ACC TAG CTC ATG ATG CCA | T. tomentosa | 4 | 2 | 50 | 12 | 100–500 | 12 | 100 |
| P. nigra | 4 | 2 | 50 | 12 | 75–1300 | 10 | 83.3 | ||
| F. excelsior | 4 | 1 | 25 | 7 | 100–600 | 1 | 14.28 | ||
| iPBS—2243 | AGT CAG GCT CTG TTA CCA | T. tomentosa | 4 | 1 | 25 | 6 | 350–900 | 1 | 16.66 |
| P. nigra | 4 | 2 | 50 | 10 | 250–1300 | 2 | 20 | ||
| F. excelsior | 4 | 3 | 75 | 10 | 250–1000 | 3 | 30 | ||
| iPBS—2394 | GAG CCT AGG CCA | T. tomentosa | 4 | 4 | 100 | 17 | 100–1000 | 16 | 94.11 |
| P. nigra | 4 | 4 | 100 | 10 | 75–900 | 7 | 70 | ||
| F. excelsior | 4 | 4 | 100 | 8 | 100–700 | 4 | 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, S.; Petkova, M. Plant Traits of Tilia tomentosa Moench, Fraxinus excelsior L., and Pinus nigra J.F.Arnold as a Proxy of Urbanization. Forests 2023, 14, 800. https://doi.org/10.3390/f14040800
Petrova S, Petkova M. Plant Traits of Tilia tomentosa Moench, Fraxinus excelsior L., and Pinus nigra J.F.Arnold as a Proxy of Urbanization. Forests. 2023; 14(4):800. https://doi.org/10.3390/f14040800
Chicago/Turabian StylePetrova, Slaveya, and Mariana Petkova. 2023. "Plant Traits of Tilia tomentosa Moench, Fraxinus excelsior L., and Pinus nigra J.F.Arnold as a Proxy of Urbanization" Forests 14, no. 4: 800. https://doi.org/10.3390/f14040800
APA StylePetrova, S., & Petkova, M. (2023). Plant Traits of Tilia tomentosa Moench, Fraxinus excelsior L., and Pinus nigra J.F.Arnold as a Proxy of Urbanization. Forests, 14(4), 800. https://doi.org/10.3390/f14040800