
Quercus suber Allometry in the West Mediterranean Basin


 ,
,  ,
, 
Abstract
1. Introduction
- join Quercus suber biomass datasets and compare the different allometries in cork oak growth among three countries—Portugal, Spain and Tunisia;
- is it feasible to develop a joint model that can consistently estimate biomass in the regions where cork oak is prevalent?
- is the addition of total tree height significant for biomass prediction?
- provide two alternative root biomass models and two additive SUR aboveground biomass models—one set as a function of du, to be used when tree height has not been measured, and the other as a function of du and h.
2. Materials and Methods
2.1. Dataset Description
2.2. Model Fitting
- (1)
- fitting individual allometric equations for each biomass compartment using the reduced and full model, without differentiating between countries;
- (2)
- addition of dummy variables for each country at each individual equation and selection of a model for each biomass compartment, in which all the parameters were significantly different from zero;
- (3)
- fitting aboveground components in a system of equations, in order to guarantee the additivity between total biomass and the other two biomass components.
3. Results
3.1. The Need for Country Specific Models and for including Total Height
3.2. Two Systems of Equations for Aboveground Biomass
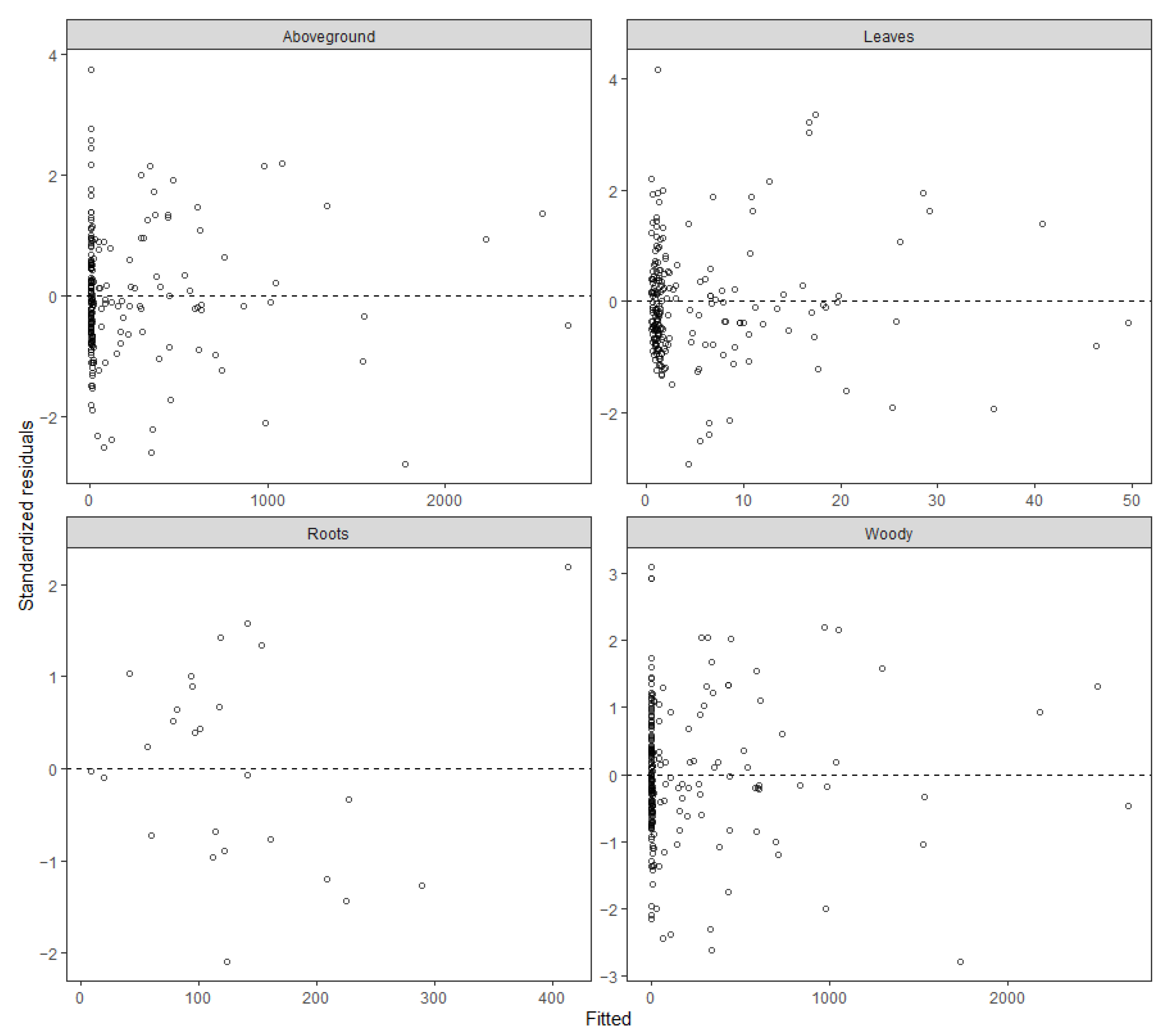
3.3. Evaluating the Models
4. Discussion
4.1. Advantages and Disadvantages of Joining a Dataset from Three Countries
4.2. Is It Feasible to Develop a Joint Model That Can Consistently Estimate Biomass in the Regions Where Cork Oak Is Prevalent?
4.3. Is the Addition of Total Tree Height Significant for Biomass Prediction?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pörtner, H.-O.; Roberts, D.C.; Tignor, M.; Poloczanska, E.S.; Mintenbeck, K.; Alegría, A.; Craig, M.; Langsdorf, S.; Löschke, S.; Möller, V.; et al. Climate Change 2022: Impacts, Adaptation, and Vulnerability; IPCC: Geneva, Switzerland, 2022. [Google Scholar]
- Keenan, T.F.; Williams, C.A. The Terrestrial Carbon Sink. Annu. Rev. Environ. Resour. 2018, 43, 219–243. [Google Scholar] [CrossRef]
- Santos, F.D.; Ferreira, P.L.; Pedersen, J.S.T. The Climate Change Challenge: A Review of the Barriers and Solutions to Deliver a Paris Solution. Climate 2022, 10, 75. [Google Scholar] [CrossRef]
- Andersen, S.P.; Allen, B.; Domingo, G.C. Biomass in the EU Green Deal: Towards Consensus on the Use of Biomass for EU Bioenergy; Policy Report; Institute for European Environmental Policy (IEEP): Brussels, Belgium, 2021. [Google Scholar]
- Brown, S. Measuring Carbon in Forests: Current Status and Future Challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef] [PubMed]
- António, N.; Tomé, M.; Tomé, J.; Soares, P.; Fontes, L. Effect of Tree, Stand, and Site Variables on the Allometry of Eucalyptus Globulus Tree Biomass. Can. J. For. Res. 2007, 37, 895–906. [Google Scholar] [CrossRef]
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and Stem Volume Equations for Tree Species in Europe; Silva Fennica, The Finnish Society of Forest Science: Helsinki, Finland, 2005; Volume 4, ISBN 9514019830. [Google Scholar]
- Weiskittel, A.R.; MacFarlane, D.W.; Radtke, P.J.; Affleck, D.L.R.; Temesgen, H.; Woodall, C.W.; Westfall, J.A.; Coulston, J.W. A Call to Improve Methods for Estimating Tree Biomass for Regional and National Assessments. J. For. 2015, 113, 414–424. [Google Scholar] [CrossRef]
- Ter-Mikaelian, M.T.; Korzukhin, M.D. Biomass Equations for Sixty-Five North American Tree Species. For. Ecol. Manag. 1997, 97, 1–24. [Google Scholar] [CrossRef]
- Trautenmüller, J.W.; Péllico Netto, S.; Balbinot, R.; Watzlawick, L.F.; Dalla Corte, A.P.; Sanquetta, C.R.; Behling, A. Regression Estimators for Aboveground Biomass and Its Constituent Parts of Trees in Native Southern Brazilian Forests. Ecol. Indic. 2021, 130, 108025. [Google Scholar] [CrossRef]
- Parresol, B.R. Assessing Tree and Stand Biomass: A Review with Examples and Critical Comparisons. For. Sci. 1999, 4, 573–593. [Google Scholar]
- Parresol, B.R. Additivity of Nonlinear Biomass Equations. Can. J. For. Res. 2001, 31, 865–878. [Google Scholar] [CrossRef]
- Behling, A.; Netto, S.P.; Sanquetta, C.R.; Corte, A.P.D.; Affleck, D.L.R.; Rodrigues, A.L.; Behling, M. Critical Analyses When Modeling Tree Biomass to Ensure Additivity of Its Components. An. Acad. Bras. Cienc. 2018, 90, 1759–1774. [Google Scholar] [CrossRef]
- Carvalho, J.P.; Parresol, B.R. Additivity in Tree Biomass Components of Pyrenean Oak (Quercus pyrenaica Willd.). For. Ecol. Manag. 2003, 179, 269–276. [Google Scholar] [CrossRef]
- Poudel, K.P.; Temesgen, H. Methods for Estimating Aboveground Biomass and Its Components for Douglas-Fir and Lodgepole Pine Trees. Can. J. For. Res. 2015, 46, 77–87. [Google Scholar] [CrossRef]
- Xie, L.; Li, F.; Zhang, L.; Widagdo, F.R.A.; Dong, L. A Bayesian Approach to Estimating Seemingly Unrelated Regression for Tree Biomass Model Systems. Forests 2020, 11, 1302. [Google Scholar] [CrossRef]
- Picard, N.; Boyemba Bosela, F.; Rossi, V. Reducing the Error in Biomass Estimates Strongly Depends on Model Selection. Ann. For. Sci. 2015, 72, 811–823. [Google Scholar] [CrossRef]
- Sileshi, G.W. A Critical Review of Forest Biomass Estimation Models, Common Mistakes and Corrective Measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Oubrahim, H.; Boulmane, M.; R Bakker, M.; Augusto, L.; Mohammed, H. Carbon Storage in Degraded Cork Oak (Quercus suber) Forest on Flat Lowlands in Morocco. Iforest Biogeosci. For. 2016, 9, 125–137. [Google Scholar] [CrossRef]
- Palma, J.H.N.; Paulo, J.A.; Tomé, M. Carbon Sequestration of Modern Quercus suber L. Silvoarable Agroforestry Systems in Portugal: A YieldSAFE-Based Estimation. Agrofor. Syst. 2014, 88, 791–801. [Google Scholar] [CrossRef]
- Correia, A.C.; Costa e Silva, F.; Correia, A.V.; Hussain, M.Z.; Rodrigues, A.D.; David, J.S.; Pereira, J.S. Carbon Sink Strength of a Mediterranean Cork Oak Understorey: How Do Semi-Deciduous and Evergreen Shrubs Face Summer Drought? J. Veg. Sci. 2014, 25, 411–426. [Google Scholar] [CrossRef]
- Paulo, J.A.; Tomé, M. Equações Para Estimação Do Volume e Biomassa de Duas Espécies de Carvalhos: Quercus Suber e Quercus Ilex; Publicações GIMREF RC1/2006; Universidade Técnica de Lisboa—Instituto Superior de Agronomia, Centro de Estudos Florestais: Lisboa, Portugal, 2006. [Google Scholar]
- Ruiz-Peinado, R.; Montero, G.; Del Rio, M. Biomass Models to Estimate Carbon Stocks for Hardwood Tree Species. For. Syst. 2012, 21, 42. [Google Scholar] [CrossRef]
- Zribi, L.; Chaar, H.; Khaldi, A.; Hanchi, B.; Mouillot, F.; Gharbi, F. Estimate of Biomass and Carbon Pools in Disturbed and Undisturbed Oak Forests in Tunisia. For. Syst. 2016, 25, e060. [Google Scholar] [CrossRef]
- Montero, G.; Ruiz-peinado, R.; Muñoz, M. Producción de Biomasa y Fijación de CO2 Por Los Bosques Españoles; INIA-Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria: Madrid, Spain, 2005; Volume 13, ISBN 84-7498-512-9. [Google Scholar]
- Laporta, L.; Domingos, T.; Marta-Pedroso, C. It’s a Keeper: Valuing the Carbon Storage Service of Agroforestry Ecosystems in the Context of CAP Eco-Schemes. Land Use Policy 2021, 109, 105712. [Google Scholar] [CrossRef]
- von Essen, M.; do Rosário, I.T.; Santos-Reis, M.; Nicholas, K.A. Valuing and Mapping Cork and Carbon across Land Use Scenarios in a Portuguese Montado Landscape. PLoS ONE 2019, 14, e02121. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.B.; Paulo, J.A.; Palma, J.H.N.; Tomé, M. Contribution of Cork Oak Plantations Installed after 1990 in Portugal to the Kyoto Commitments and to the Landowners Economy. For. Policy Econ. 2012, 17, 59–68. [Google Scholar] [CrossRef]
- Cañellas, I.; Sánchez-González, M.; Adame, P.; Moreno-Fernández, D.; Bogino, S.M.; Herrero, C.; Roig, S.; Tomé, M.; Paulo, J.A.; Bravo, F. Carbon Sequestration in Mediterranenan Oak Forests. In Managing Forest Ecosystems: The Challenge of Climate Change; Bravo, F., LeMay, V., Jandl, R., Eds.; Springer: Cham, Switzerland, 2017; pp. 403–427. [Google Scholar]
- Demertzi, M.; Sierra-Pérez, J.; Paulo, J.A.; Arroja, L.; Dias, A.C. Environmental Performance of Expanded Cork Slab and Granules through Life Cycle Assessment. J. Clean. Prod. 2017, 145, 294–302. [Google Scholar] [CrossRef]
- Demertzi, M.; Paulo, J.A.; Arroja, L.; Dias, A.C. A Carbon Footprint Simulation Model for the Cork Oak Sector. Sci. Total Environ. 2016, 566–567, 499–511. [Google Scholar] [CrossRef]
- Neumann, M.; Moreno, A.; Mues, V.; Härkönen, S.; Mura, M.; Bouriaud, O.; Lang, M.; Achten, W.M.J.; Thivolle-Cazat, A.; Bronisz, K.; et al. Comparison of Carbon Estimation Methods for European Forests. For. Ecol. Manag. 2016, 361, 397–420. [Google Scholar] [CrossRef]
- Paulo, J.; Tomé, M. Recolha de Dados Para Determinação de Biomassas e Volumes de Sobreiro (Quercus suber). Protocolo Para a Instalação de Parcelas Temporárias e Medição de Árvores Amostra; Publicações FORCHANGE RT3/2008; Universidade Técnica de Lisboa—Instituto Superior de Agronomia, Centro de Estudos Florestais: Lisboa, Portugal, 2008. [Google Scholar]
- Paulo, J.A.; Tomé, M.; Uva, J.S. Recolha de Dados de Biomassa de Azinheiras (Quercus Ilex). Protocolo Para a Instalação de Parcelas Temporárias e Medição Das Árvores Amostra.; Publicações GIMREF RT6/2001; Universidade Técnica de Lisboa—Instituto Superior de Agronomia, Centro de Estudos Florestais: Lisboa, Portugal, 2001. [Google Scholar]
- Varela, M.C.; Bellarosa, R.; Eriksson, G.; Bariteau, M.; Catalan, G.; Branco, T.; Barros, I.R.; Almeida, H.; Chambel, R.; Sbay, H.; et al. Handbook of the Concerted Action “European Network for the Evaluation of Genetic Resources of Cork Oak for Appropriate Use in Breeding and Gene Conservation Strategies”. Reference—FAIR 1 CT 95-0202; Varela, M.C., Ed.; Estação Florestal Nacional: Lisboa, Portugal, 2000; ISBN 972-95736-7-0. [Google Scholar]
- Paulo, J.A.; Tomé, M. Predicting Mature Cork Biomass with t Years of Growth from One Measurement Taken at Any Other Age. For. Ecol. Manag. 2010, 259, 1993–2005. [Google Scholar] [CrossRef]
- Attiwill, P.M.; Ovington, J.D. Determination of Forest Biomass. For. Sci. 1968, 14, 13–15. [Google Scholar]
- O’Brien, S.T.; Hubbell, S.P.; Spiro, P.; Condit, R.; Foster, R.B. Diameter, Height, Crown, and Age Relationships in Eight Neotropical Tree Species. Ecology 1995, 76, 1926–1939. [Google Scholar] [CrossRef]
- Thornley, J.H.M.; Johnson, I.R. Plant and Crop Modelling; Oxford University Press: Oxford, UK, 1990. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021.
- SAS/ETS 13.2 User’s Guide; SAS Institute Inc.: Cary, NC, USA, 2014.
- Jara, M.C.; Henry, M.; Réjou-Méchain, M.; Wayson, C.; Zapata-Cuartas, M.; Piotto, D.; Alice Guier, F.; Castañeda Lombis, H.; Castellanos López, E.; Cuenca Lara, R.; et al. Guidelines for Documenting and Reporting Tree Allometric Equations. Ann. For. Sci. 2015, 72, 763–768. [Google Scholar] [CrossRef]
- Soares, P.; Tomé, M.; Skovsgaard, J.P.; Vanclay, J.K. Evaluating a Growth Model for Forest Management Using Continuous Forest Inventory Data. For. Ecol. Manage. 1995, 71, 251–265. [Google Scholar] [CrossRef]
- Vanclay, J.K.; Skovsgaard, J.P. Evaluating Forest Growth Models. Ecol. Modell. 1997, 98, 1–12. [Google Scholar] [CrossRef]
- Myers, R.H. Classical and Modern Regression with Applications, 2nd ed.; Duxbury Press: Belmont, CA, USA, 1990. [Google Scholar]
- Assmann, E. The Principles of Forest Yield Study; Pergamon Press: Oxford, UK, 1970. [Google Scholar]
- Burkhart, H.E.; Tomé, M. Modeling Forest Trees and Stands; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; ISBN 978-94-007-1597-4. [Google Scholar] [CrossRef]
- Temesgen, H.; Affleck, D.; Poudel, K.; Gray, A.; Sessions, J. A Review of the Challenges and Opportunities in Estimating above Ground Forest Biomass Using Tree-Level Models. Scand. J. For. Res. 2015, 30, 326–335. [Google Scholar] [CrossRef]
- Forrester, D.I.; Tachauer, I.H.H.; Annighoefer, P.; Barbeito, I.; Pretzsch, H.; Ruiz-Peinado, R.; Stark, H.; Vacchiano, G.; Zlatanov, T.; Chakraborty, T.; et al. Generalized Biomass and Leaf Area Allometric Equations for European Tree Species Incorporating Stand Structure, Tree Age and Climate. For. Ecol. Manag. 2017, 396, 160–175. [Google Scholar] [CrossRef]
- Dutcă, I.; Mather, R.; Ioraș, F. Sampling Trees to Develop Allometric Biomass Models: How Does Tree Selection Affect Model Prediction Accuracy and Precision? Ecol. Indic. 2020, 117, 106553. [Google Scholar] [CrossRef]
- David, T.S.; Pinto, C.A.; Nadezhdina, N.; Kurz-Besson, C.; Henriques, M.O.; Quilhó, T.; Cermak, J.; Chaves, M.M.; Pereira, J.S.; David, J.S. Root Functioning, Tree Water Use and Hydraulic Redistribution in Quercus suber Trees: A Modeling Approach Based on Root Sap Flow. For. Ecol. Manag. 2013, 307, 136–146. [Google Scholar] [CrossRef]
- David, T.S.; Pinto, C.A.; Nadezhdina, N.; David, J.S. Water and Forests in the Mediterranean Hot Climate Zone: A Review Based on a Hydraulic Interpretation of Tree Functioning. For. Syst. 2016, 25, eR02. [Google Scholar] [CrossRef]
- Paulo, J.A.; Tomé, J.; Tomé, M. Ajustamento Simultâneo de Equações de Biomassa de Azinheira. In Proceedings of the Actas do X Congresso da Sociedades Portuguesa de Estadística, Porto, Portugal, 25–28 September 2002. [Google Scholar]
- Andivia, E.; Bou, J.; Fernández, M.; Caritat, A.; Alejano, R.; Vilar, L.; Vázquez-Piqué, J. Assessing the Relative Role of Climate on Litterfall in Mediterranean Cork Oak Forests. IForest 2018, 11, 786–793. [Google Scholar] [CrossRef]
- Sánchez-González, M.; Cañellas, I.; Montero, G. Generalized Height-Diameter and Crown Diameter Prediction Models for Cork Oak Forests in Spain. Investig. Agrar. Sist. Recur. For. 2007, 16, 76. [Google Scholar] [CrossRef]
- Menéndez-Miguélez, M.; Calama, R.; Del Río, M.; Madrigal, G.; López-Senespleda, E.; Pardos, M.; Ruiz-Peinado, R. Species-Specific and Generalized Biomass Models for Estimating Carbon Stocks of Young Reforestations. Biomass Bioenergy 2022, 161, 10645. [Google Scholar] [CrossRef]
- Sánchez-González, M.; Calama, R.; Cañellas, I.; Montero, G. Variables Influencing Cork Thickness in Spanish Cork Oak Forests: A Modelling Approach. Ann. For. Sci. 2007, 64, 301–312. [Google Scholar] [CrossRef]
- Sánchez-González, M.; Tomé, M.; Montero, G. Modelling Height and Diameter Growth of Dominant Cork Oak Trees in Spain. Ann. For. Sci. 2007, 62, 633–643. [Google Scholar] [CrossRef]
- Pasalodos-Tato, M.; Pukkala, T.; Cañellas, I.; Sánchez-González, M. Optimizing the Debarking and Cutting Schedule of Cork Oak Stands. Ann. For. Sci. 2018, 75, 61. [Google Scholar] [CrossRef]
- Faias, S.P.; Paulo, J.A.; Tomé, M. Inter-Tree Competition Analysis in Undebarked Cork Oak Plantations as a Support Tool for Management in Portugal. New For. 2020, 51, 489–505. [Google Scholar] [CrossRef]
- Palma, J.H.N.; Paulo, J.A.; Faias, S.P.; Garcia-Gonzalo, J.; Borges, J.G.; Tomé, M. Adaptive Management and Debarking Schedule Optimization of Quercus Suber L. Stands under Climate Change: Case Study in Chamusca, Portugal. Reg. Environ. Chang. 2015, 15, 1569–1580. [Google Scholar] [CrossRef]
- Campos, P.; Daly-Hassen, H.; Oviedo, J.L.; Ovando, P.; Chebil, A. Accounting for Single and Aggregated Forest Incomes: Application to Public Cork Oak Forests in Jerez (Spain) and Iteimia (Tunisia). Ecol. Econ. 2008, 65, 76–86. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable 1 | n | Min. | Median | Mean | Max. | sd |
|---|---|---|---|---|---|---|
| du (cm) | 212 | 2.45 | 5.95 | 13.49 | 79.44 | 14.68 |
| h (m) | 212 | 1.52 | 2.44 | 4.75 | 15.80 | 3.93 |
| wl (kg) | 212 | 0.10 | 1.25 | 4.95 | 51.28 | 8.52 |
| ww (kg) | 212 | 0.64 | 6.51 | 178.86 | 3032.49 | 412.02 |
| wa (kg) | 212 | 0.99 | 7.67 | 183.81 | 3078.68 | 418.79 |
| wr (kg) | 26 | 7.42 | 116.67 | 132.10 | 454.06 | 88.89 |
| Model Description | Coefficients | Performance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reduced model | wl | 0.09 *** | 0.02 *** | 0.06 *** | 1.58 *** | 0.86 | |||||
| ww | 0.08 *** | 0.24 * | 2.60 *** | 2.13 *** | 2.49 *** | 0.87 | |||||
| wa | 0.15 *** | 2.42 *** | 2.23 *** | 2.31 *** | 0.88 | ||||||
| wr 1 | 0.26 2 | 1.76 *** | 1.86 *** | 0.87 | |||||||
| Full model | wl | 0.10 *** | 0.02 *** | 0.05 *** | 1.21 *** | 1.80 *** | 0.89 | ||||
| ww | 0.10 *** | 2.07 *** | 1.48 ** | 1.84 *** | 0.75 *** | 1.41 * | 0.92 | ||||
| wa | 0.14 *** | 1.97 *** | 1.81 *** | 1.75 *** | 0.74 *** | 0.89 | |||||
| wr 1 | 0.16 2 | 1.43 *** | 0.76 *** | 0.89 | |||||||
| Model Description | Coefficients | Performance | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Reduced model | wl | 0.09 *** | 0.02 *** | 0.06 *** | 1.60 *** | 0.86 | |||||
| ww | 0.08 *** | 0.26 ** | 2.59 *** | 2.09 *** | 2.48 *** | 0.89 | |||||
| wa | wl + ww | 0.89 | |||||||||
| Full model | wl | 0.11 *** | 0.03 *** | 0.05 *** | 1.13 *** | 0.65 *** | 0.89 | ||||
| ww | 0.10 *** | 2.07 *** | 1.40 *** | 1.84 *** | 0.75 *** | 1.54 ** | 0.93 | ||||
| wa | wl + ww | 0.93 | |||||||||
| Model Description | Cross-Validation Metrics | ||||
|---|---|---|---|---|---|
| mPRESS | maPRESS | ||||
| INDIVIDUAL EQUATIONS | Reduced model | wl | −0.0032 | 0.14 | 0.84 |
| ww | −0.0005 | 0.06 | 0.85 | ||
| wa | −0.0286 | 0.36 | 0.86 | ||
| wr | 3.17 | 25.50 | 0.59 | ||
| Full model | wl | −0.0013 | 0.08 | 0.87 | |
| ww | 0.0002 | 0.04 | 0.90 | ||
| wa | 0.0002 | 0.05 | 0.88 | ||
| wr | 1.48 | 24.28 | 0.84 | ||
| SUR | Reduced model | wl | 0.0029 | 0.14 | 0.86 |
| ww | 0.0020 | 0.04 | 0.89 | ||
| wa | 0.0053 | 0.08 | 0.89 | ||
| Full model | wl | −0.0060 | 0.08 | 0.89 | |
| ww | −0.0002 | 0.04 | 0.93 | ||
| wa | −0.0007 | 0.05 | 0.93 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jorge, C.; Tomé, M.; Ruiz-Peinado, R.; Zribi, L.; Paulo, J.A. Quercus suber Allometry in the West Mediterranean Basin. Forests 2023, 14, 649. https://doi.org/10.3390/f14030649
Jorge C, Tomé M, Ruiz-Peinado R, Zribi L, Paulo JA. Quercus suber Allometry in the West Mediterranean Basin. Forests. 2023; 14(3):649. https://doi.org/10.3390/f14030649
Chicago/Turabian StyleJorge, Catarina, Margarida Tomé, Ricardo Ruiz-Peinado, Lobna Zribi, and Joana Amaral Paulo. 2023. "Quercus suber Allometry in the West Mediterranean Basin" Forests 14, no. 3: 649. https://doi.org/10.3390/f14030649
APA StyleJorge, C., Tomé, M., Ruiz-Peinado, R., Zribi, L., & Paulo, J. A. (2023). Quercus suber Allometry in the West Mediterranean Basin. Forests, 14(3), 649. https://doi.org/10.3390/f14030649